

Abridged English version
Estuarine environments are in constant reduction under the pressure of slow changes in sea level. This trend is accelerated by shore management impacts (such as land reclamation, and dyking...). Long-term monitoring is thus necessary to understand the induced consequences on the functioning of these systems and to predict impacts of anthropic activities.
A long-term monitoring of the morpho-sedimentary facies and associated macrobenthic fauna has been carried out since 1969 in the Bay des Veys (French coast of the Eastern English Channel) and a predictive evolution pattern has been elaborated. Within this context, a new map of the macrozoobenthic communities allows us to discuss the evolution of their spatial distribution compared to their predicted pattern.
This estuarine bay of 37 km2, open toward NNE, is submitted to a 7-m tidal range. The latest phase of land reclamation, in 1972, was followed by an important development of oyster farming in the eastern mud flats. The macrozoobentic cartography was obtained from a set of 74 sample stations in 2000 (three samples of 0.02 m2 per station, sewed on a 1-mm square mesh) distributed along 16 transects.
Communities were characterized by factorial component analysis and hierarchical clustering, with the help of constancy C and fidelity F indexes applied to the main species.
Sixty-seven taxa were identified, with crustaceans, molluscs and annelids contributing each a third of the total number. Bivalves and annelids represent respectively 75 and 20% of total biomasses. Biomasses were higher in the far end of the bay and decreased towards the mouth. Density followed the same pattern, but with an east–west asymmetry. Density was the highest in the tidal flats of the western part of the bay due to the large abundance of Hydrobia ulvae and Corophium arenarium.
Benthos was distributed around the Boreal-Atlantic Macoma community. The five principal communities identified followed the same scheme. In the far end of the bay, we found a transitional estuarine assemblage characterized by Mya arenaria, Abra tenuis, H. ulvae and Cyathura carinata. In the midway part, we observed a composite estuarine assemblage under marine influence. In the central and western zones, communities were typical of surge-exposed marine influence, with the presence of Bathyporeia sarsi and, locally, with B. pilosa and C. arenarium and the high densities of H. ulvae. The eastern tidal flat was under sheltered marine influence, with Lanice conchilega and especially Scoloplos armiger. In the most external area, marine influence was detected by the presence of B. elegans.
In comparison with previous studies and the predictable model, we found the same estuarine-marine gradient and an increase of eastern tidal flat particularity. The continuous marine intrusion on the central tidal flat was amplified, with a fading of the local transitional community and the expansion of a marine influence-community towards the northern area of the bay. Consequently, the central and western biocenotic characters were mixed. On the eastern tidal flat, protected by the channel, changes occurred with the expansion of L. conchilega associated with oyster farming management, to the detriment of the more typical community characterised by U. poseidonis and S. armiger. We only found a residual patch of the community in the southern area of the oyster beds, with a recent enrichment with Cerastoderma edule.
Comparison between 2000 data and precedent results shows a constant evolution of the western and central macrozoobenthos communities of the bay, which are under marine influence. Before 1985, eastern macrozoobenthos communities had not shown any obvious evolution, but since then the development of oyster farming activities has induced fast transformations with consequently a critical change in biocenotic communities.
1 Introduction
Les milieux littoraux sont soumis à d'importantes modifications environnementales, tant naturelles qu'anthropiques, en particulier dans la frange intertidale estuarienne où les paramètres écologiques peuvent rapidement évoluer. La transgression marine, entraı̂nant un comblement naturel, est accélérée par les aménagements littoraux (polders, endigages, canalisations, etc.). À cette évolution générale se superposent d'autres impacts anthropiques (effluents urbains et industriels, conchyliculture, etc.), qui peuvent fortement perturber le fonctionnement de ces milieux.
Le macrozoobenthos, en tant que maillon intermédiaire de la chaı̂ne alimentaire, intègre rapidement les modifications des caractéristiques environnementales [1]. Le suivi de la faune macrozoobenthique permet d'étudier les conséquences de ces évolutions naturelles et anthropiques, tout en intégrant les modifications biotiques et abiotiques du milieu [2,3].
En baie des Veys, le suivi de la macrofaune benthique est associé à une étude sédimentaire à long terme. Étudiés de façon continue entre 1973 et 1999, huit campagnes de terrain successives ont permis de suivre l'évolution biosédimentaire ; ces campagnes ont débouché sur des représentations cartographiques dont l'enchaı̂nement a permis de définir une cinétique spatio-temporelle des grands ensembles caractéristiques (estuarien, marin ou de transition). L'extrapolation de cette analyse fonctionnelle de la baie des Veys constitue la base d'un modèle prédictif proposé par Sylvand [4].
Nous proposons de vérifier les hypothèses émises dans ce modèle concernant l'évolution du macrozoobenthos :
- (1) à l'ouest, une intrusion d'espèces marine vers le sud ;
- (2) au centre, une lente progression des espèces estuariennes vers le nord ;
- (3) à l'est, une régression des espèces originelles en raison d'une extension depuis le sud d'espèces estuariennes et depuis le nord d'un substrat à Lanice conchilega associé à la conchyliculture.
Dans cet objectif, nous utilisons la cartographie de 1985, la plus documentée, comme référence [3,5–9] ce qui nous permet d'étudier l'évolution du macrozoobenthos en 15 ans. Nous décrivons la répartition du macrozoobenthos en 2000, puis nous la comparons à celle observée en 1985.
Nous avons réalisé en 2000 un échantillonnage quantitatif stratifié organisé suivant des radiales, qui concerne la majeure partie de la zone intertidale de la baie. Nous identifions et décrivons les principaux assemblages d'espèces présents en 2000. Leurs répartitions sont obtenues en regroupant les stations d'échantillonnage dont les assemblages d'espèces sont proches en composition et structure numérique. Ces assemblages d'espèces sont ensuite caractérisés par leurs espèces dominantes ou caractéristiques [10]. En extrapolant les principaux résultats obtenus, nous avons élaboré des représentations cartographiques des assemblages macrozoobenthiques. Nous faisons correspondre chacun de ces assemblages à ceux obtenus lors de la cartographie de référence de 1985. Ils sont comparés sur la base de leur structure spécifique, numérique et de leur répartition spatiale.
Nous discutons ensuite ces évolutions et celles envisagées par le modèle prédictif.
2 Matériels et méthodes
2.1 Site d'étude
La baie des Veys se situe sur le littoral de la baie de Seine occidentale (France, Manche orientale). S'ouvrant vers le nord-nord-est sur près de 8 km, elle est protégée des houles dominantes de secteur ouest. Elle est soumise à un marnage de l'ordre de 7 m (régime macrotidal) et découvre à basse mer sur près de 37 km2 (Fig. 1). Elle comporte un schorre de 4 km2 dominé par la spartine (Spartina anglica). La slikke de la zone d'étude, qui couvre 23 km2, est parcourue par des chenaux de Carentan (ouest) et d'Isigny (est) et leurs divers bras morts. La baie se partage ainsi en quatre parties : le flanc ouest, l'estran central délimité par les deux chenaux, le flanc est et enfin les bancs de sables du centre-nord de la baie, en dehors de la zone d'étude.
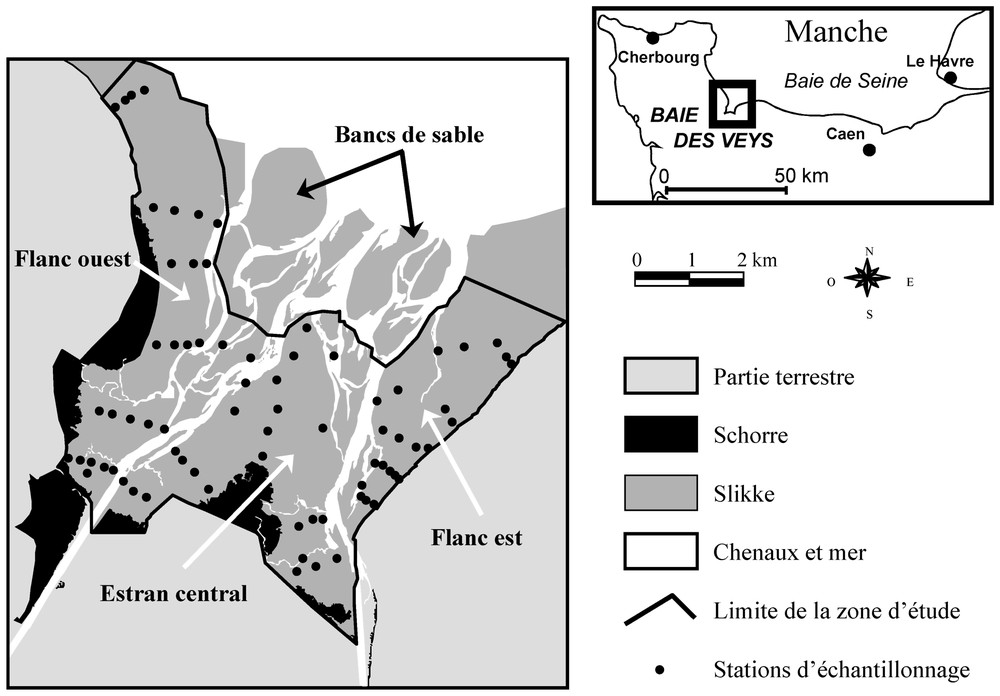
Zone d'étude et localisation des 74 stations d'échantillonnage de 2000, organisées en 16 transects et complétées par six stations supplémentaires. Trois grands secteurs, délimités par les chenaux, ont été échantillonnés : le flanc ouest, l'estran central et le flanc est. Les bancs de sable inaccessibles du centre de la baie n'ont pas pu être échantillonnés.
Comme la majorité des baies et estuaires de la façade ouest de l'Europe, le site a été l'objet d'une succession d'endigages, ainsi que d'un développement d'activités conchylicoles. Les premiers endigages commencèrent en baie des Veys au début du XVIIIe siècle, le dernier s'étant achevé en 1972 [11]. Sur ce site, des concessions conchylicoles sur bouchots et tables s'étendent sur 22 ha au nord-ouest. Au nord-est, des tables conchylicoles (ostréiculture dominante) ont été installées à partir des 1968 et couvrent actuellement 162 ha, après une forte extension en 1985.
2.2 Méthodologie
Afin de cartographier le macrozoobenthos de la baie des Veys, nous avons repris le plan général d'échantillonnage utilisé dans les campagnes précédentes. En novembre 2000, l'échantillonnage stratifié [12] est organisé selon 16 radiales, complété par six stations isolées (Fig. 1). Chaque radiale comprend trois à six stations, réparties depuis le plus haut niveau de la slikke jusqu'aux chenaux ou à la mer. À chaque station, trois échantillons ont été prélevés. Les échantillons sont extraits à l'aide d'un carottier (TASM, [13]) de 0,02 m2, sur une profondeur de 25 cm. Ils sont ensuite tamisés sur une maille carrée de 1 mm [14]. Pour tenir compte de certaines espèces qui peuvent s'enfouir plus profondément (principalement Mya arenaria, mais aussi Arenicola marina et Scrobicularia plana) et représenter un complément de biomasse important [4], l'échantillonnage est complété par un prélèvement à la bêche sur les 25 cm suivants. Les échantillons sont fixés par une solution salée de formaldéhyde à 5 % additionnée de floxine B. Les biomasses, exprimées en gramme de poids sec sans cendre (AFDW), ont été obtenues par séchage à l'étuve à 60 °C pendant une semaine, puis calcination au four à 450 °C pendant 3 h.
En 1985, l'échantillonnage a été réalisé sur sept radiales, selon la même méthode, totalisant 31 stations. La profondeur de prélèvement se limitait aux 25 premiers centimètres.
2.3 Analyse des données
2.3.1 Macrozoobenthos en 2000
Pour caractériser l'ensemble des relevés effectués en 2000, nous utilisons, comme descripteurs du macrozoobenthos : l'abondance (rapportée au mètre carré), l'indice de richesse spécifique S (calculé à partir des taxa identifiés, certains d'entre eux représentant plusieurs espèces), l'indice synthétique de diversité de Shannon–Weaver H′ [15] et la biomasse (rapportée au mètre carré). Ces descripteurs sont calculés après cumul des répliquats de chaque station. Les moyennes sont accompagnées de leur écart type (s.d.).
Une analyse factorielle des correspondances [16], couplée à une classification ascendante hiérarchique sur les distances euclidiennes des coordonnées factorielles, en utilisant la méthode de classification de Ward [17], permet de déterminer les différents groupes de stations présentant des assemblages d'espèces de composition spécifique et numérique similaire. Cette analyse est effectuée avec le logiciel ADE-4 [18]. Les abondances de chaque taxa par station ont été normalisées par transformation log10(x+1) [3,19–21]. Les espèces présentant une fréquence d'occurrence inférieure à 5 % ont été exclues de même qu'une station pratiquement dépourvue de macrozoobenthos.
Pour identifier les espèces caractérisant les assemblages obtenus, nous utilisons deux indices biocénotiques employés par Bachelet, Dauvin [19,20] et Retière [22] : l'indice de constance C, qui est le rapport entre le nombre de stations de l'assemblage où l'espèce est présente et le nombre total de stations de cet assemblage, exprimé en pourcentage, et l'indice de fidélité F, défini par le rapport entre l'indice de constance d'une espèce dans un assemblage et la somme des deux indices de constance maximum de l'espèce sur l'ensemble des assemblages considérés [22]. Pour caractériser les différents assemblages, nous utilisons les espèces préférentes (66,7 %⩽F<90 %), électives (90 %⩽F<100 %) et exclusives (F=100 %), ainsi que les espèces constantes (50 %<C⩽75 %) et régulières (C>75 %). La classe régulière a été créée pour cette étude en raison de l'importance du nombre d'espèces constantes. Les espèces communes (25 %<C⩽50 %) ne sont citées qu'à titre indicatif.
Nous cartographions la répartition du macrozoobenthos en 2000 en reliant spatialement les groupes de stations ayant les même assemblages d'espèces.
2.3.2 Comparaison du macrozoobenthos entre 1985 et 2000
À partir des répartitions spatiales des assemblages d'espèces, nous associons les assemblages de 1985 correspondants à ceux de 2000. Pour confirmer ces correspondances et comparer les évolutions de chacun de ces assemblages, nous les classons en utilisant la même technique de classification que celle décrite précédemment (2.2.2). Cette analyse est réalisée sur les abondances spécifiques moyennes des différents assemblages de 1985 et de 2000. Seuls les taxa 24 ayant une fréquence d'occurrence supérieure à 5 % sur les deux périodes d'échantillonnage ont été retenus. M. arenaria, non échantillonnée en 1985, n'a pas été retenue pour cette analyse. En raison d'incertitudes sur l'identification des espèces du genre Corophium en 1985, nous avons additionné les abondances des différentes espèces de ce genre et les avons notés Corophium spp. Les différences entre assemblages d'espèces sont exprimées en pourcentage de dissimilarité maximale observée.
Après avoir caractérisé les différents assemblages de 1985 par la constance et la fidélité de leurs taxa comme pour ceux de 2000, nous comparons ensuite, entre 1985 et 2000, leur composition en espèces caractéristiques (accompagnatrices et discriminantes).
La cartographie de 1985 [3,5–9] est ensuite comparée à celle de 2000 afin de décrire l'évolution spatiale des différents assemblages d'espèces.
3 Résultats
3.1 Macrozoobenthos en 2000
3.1.1 Caractéristiques générales
Les trois groupes taxonomiques les plus représentés (Tableau 1), en nombre de taxa, sont les Annélides (27 taxa), les Arthropodes (24 taxa) et les Mollusques (12 taxa). Ils constituent la quasi-totalité de l'abondance (99,7 %) et de la biomasse (99,5 %). Les Arthropodes sont les plus représentés, avec plus d'un tiers (37,5 %) des 15 700 individus capturés. Ils contribuent pourtant peu à la biomasse présente en baie des Veys, avec seulement 1,5 %. Les Mollusques représentent près d'un tiers des individus (30,7 %) en raison de la forte dominance numérique du Gastéropode Hydrobia ulvae (26,2 % du benthos total), les autres Mollusques appartenant exclusivement à la classe des Bivalves, qui contribuent à la plus grande partie de la biomasse (76 %). Le dernier tiers des individus sont des Annélides (31,5 %) et représente 20 % de la biomasse.
Liste des taxa recensés en 1985 et 2000, les cellules grisées indiquent la présence du taxon aux dates de prélèvement
La richesse spécifique varie de 2 à 18 espèces par station (moyenne 9,6, s.d. 3,6). Elle est plus élevée sur la haute slikke, à proximité du schorre. Faible sur les rives du chenal ouest et sur une partie du chenal est, le nombre d'espèces augmente sur les vasières de l'estran central et au milieu de l'estran est. L'indice de diversité est compris entre 0,47 et 3,49 pour une moyenne de 2,11 (s.d. 0,68). Il est faible au centre du site, particulièrement près des chenaux, et croı̂t vers le fond de la baie et vers la mer. L'abondance présente une forte variabilité (moyenne 3790 s.d. 9184 ind·m−2) et varie de 50 à 68 800 ind·m−2. Elle suit une distribution spatiale proche de celle de la richesse spécifique. Toutefois, de fortes abondances sont observées sur les niveaux supérieurs du flanc ouest, attribuées à d'importantes concentrations en H. ulvae et Corophium arenarium (26,2 % et 12,7 % de l'ensemble des individus récoltés). Les biomasses varient très fortement de 0,02 à près de 500 g·m−2. La moyenne de 41,31 (s.d. 95,48) g·m−2 n'est pas représentative de l'ensemble du site, deux zones de fond de baie présentant des biomasses extrêmement fortes dues à la présence de M. arenaria. Ce bivalve de grande taille représente en effet 58,5 % de la biomasse collectée. Pour une meilleure compréhension de la répartition spatiale de ce paramètre, les biomasses de M. arenaria ont été exclues de l'analyse. Celles du genre Ensis et de l'espèce Acrochnida brachiata ont été également supprimées de la biomasse totale pour des raisons similaires. La moyenne est alors de 16,24 (s.d. 17,63) g m−2, avec un maximum de 75,98 g m−2. Les biomasses sont alors plus importantes en fond de baie et sur les hauts niveaux, pour décroı̂tre vers la mer et les niveaux les plus bas, c'est-à-dire près des chenaux. Seule une zone à forte biomasse sur l'estran est ne suit pas ce schéma de distribution, en raison d'une forte densité de Cerastoderma edule.
3.1.2 Détermination des différents assemblages d'espèces en 2000
L'analyse factorielle des correspondances a été effectuée sur 73 stations et 30 des 33 taxa ayant une fréquence d'occurrence en 2000 supérieure à 5 %. Trois taxa présentant des distributions atypiques n'ont pas été retenus. Il s'agit de Crangon crangon, épibenthos distribué de façon aléatoire sur le site, de Glycera convoluta, présente en faible effectif et répartie elle aussi de façon aléatoire, ainsi que de Spiophanes bombyx, présentant sur une station une densité extrêmement élevée, masquant les autres distributions. La projection schématique des stations sur le plan factoriel (Fig. 2) est représentée sur les deux premiers axes, qui contribuent à 33 % de l'inertie totale. Les classifications ont été effectuées, en prenant en compte les sept premiers axes, totalisant une inertie de 64 %.
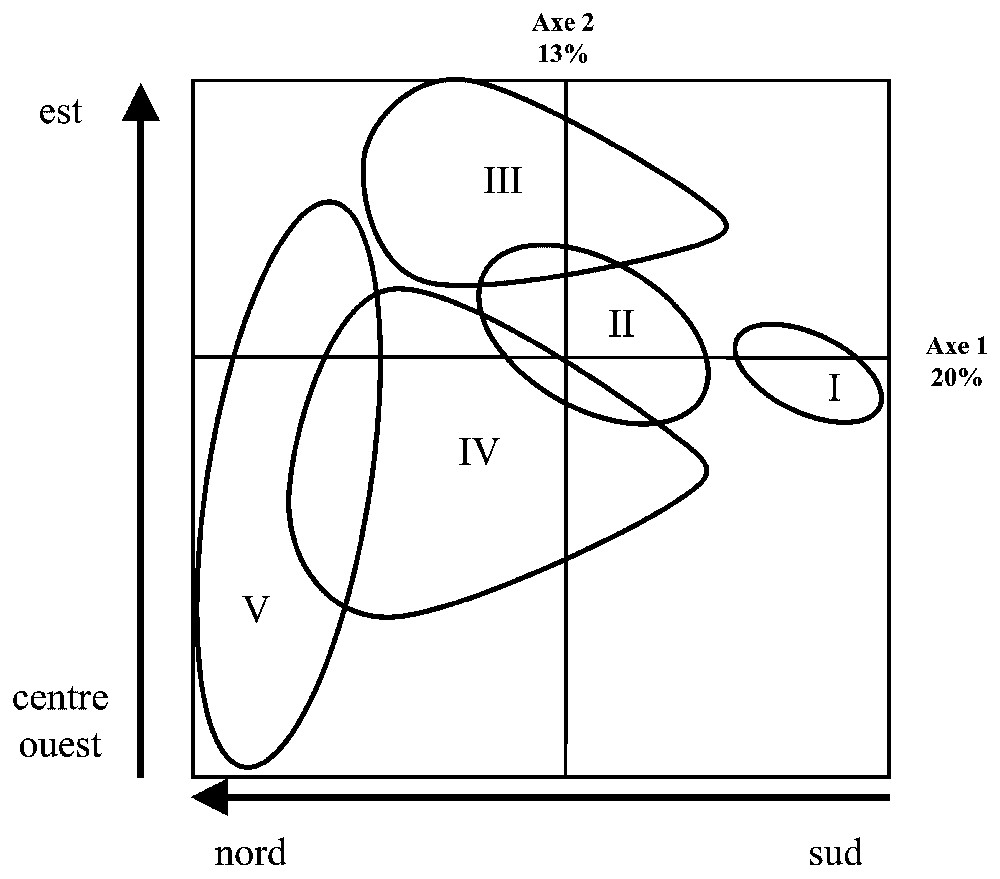
Projection schématique des stations échantillonnées en 2000 sur le premier plan de l'analyse factorielle des correspondances. Cinq groupes de stations définies selon leurs assemblages d'espèces similaires s'organisent selon un gradient sud–nord correspondant à un gradient estuarien–marin représenté par le premier axe, et selon une opposition entre les stations du centre et de l'ouest de la baie et celles située à l'est, correspondant à une opposition entre stations exposées et stations abritées, représentée par le second axe.
La projection factorielle des stations (Fig. 2), associée à la classification, présente, sur le premier axe, un gradient depuis le fond de la baie jusqu'à son embouchure. Le deuxième axe révèle un clivage est–ouest du site, matérialisé par le chenal est.
Ces stations se répartissent en cinq groupes. Le groupe I est représenté par les stations les plus internes du site. Le groupe II fait la transition entre le groupe I et les groupes III et IV. Les stations du groupe III représentent la partie est du site. Le groupe IV rassemble les stations présentes à l'ouest et au centre de la baie, dans sa partie médiane. Enfin, le groupe V rassemble les stations les plus externes situées dans la partie ouest et centrale de la baie. Ses stations présentent l'assemblage d'espèces le plus différent de l'ensemble du site, plus proche toutefois du groupe IV que des autres. Enfin, cinq stations n'ont pu être associées à un groupe. Leur composition est très variable, sans montrer de caractères particuliers, si ce n'est une pauvreté à la fois spécifique et numérique. Ces stations, présentes près des zones de divagation des chenaux, ont été exclues des représentations graphiques.
À ces cinq groupes de stations correspondent cinq assemblages d'espèces distincts, qui présentent 28 taxa ayant une valeur d'indice de constance supérieure à 25 % dans au moins un des assemblages (Tableau 2). Sept espèces sont communes, constantes ou régulières dans la plupart des cinq assemblages d'espèces et caractérisent l'ensemble du peuplement de la baie. Deux espèces occupent tout le site. Il s'agit de C. edule, et dans une moindre mesure de Pygospio elegans (qui n'est pas commune dans l'assemblage III). Deux espèces se retrouvent dans tous les assemblages, excepté celui le plus externe de la baie (l'assemblage V). Ce sont Eteone longa et Capitella capitata. De même, Nephtys hombergii, Spio filicornis et Urothoe poseidonis sont notées dans tous les assemblages, excepté le plus interne (assemblage I). Ces espèces ne permettent pas de discriminer les assemblages entre eux, mais leur sont caractéristiques. Nous les appellerons des espèces (ou taxa) accompagnatrices. Nous caractérisons donc les assemblages par les autres taxa présents, associés plus spécifiquement à un ou deux assemblages, que nous appellerons espèces (ou taxa) discriminantes.
Composition des assemblages déterminés en 2000 à partir des taxa présentant un indice de constance (C) supérieur à 25% et/ou un indice de fréquence (F) supérieur à 66,7%. c: taxon commun (25%<C⩽50%); C: taxon constant (50%<C⩽75%); R: taxon régulier (75%<C); p: taxon préférent (66,7%<F⩽90%); e: taxon électif (90%<F<100%); E: taxon exclusif (F=100%)
| Assemblages d'espèces déterminés en 2000 | |||||
| I | II | III | IV | V | |
| Mya arenaria | C–E | ||||
| Abra tenuis | c–E | ||||
| Hydrobia ulvae | C–p | ||||
| Carcinus maenas | c–p | ||||
| Cyathura carinata | c–p | ||||
| Heteromastus filiformis | R | C | |||
| Macoma balthica | R | C | |||
| Hediste diversicolor | R | R | c | ||
| Oligochaete spp. | R | C | c | ||
| Arenicola marina | C | R | c | ||
| Eteone longa | R | R | c | c | |
| Capitella capitata | c | C | C | c | |
| Cerastoderma edule | R | R | c | R | c |
| Pygospio elegans | R | C | c | ||
| Nephtys hombergii | C | C | C | C | |
| Spio filicornis | c | C | C | C | |
| Urothoe poseidonis | C | C | c | c | |
| Corophium arenarium | c | C | |||
| Lanice conchilega | c–p | ||||
| Scoloplos armiger | C–p | c | c | ||
| Eurydice pulchra | C | ||||
| Bathyporeia sarsi | c | R–p | c | ||
| B. pilosa | c | R | C | ||
| Tanaissus lilljeborgi | c | c | |||
| Acrocnida brachiata | c–p | ||||
| Nemertea spp. | c–p | ||||
| Scolelepis squamata | c–p | ||||
| B. elegans | C– E |
L'assemblage le plus interne, I, est caractérisé par trois espèces préférentes, H. ulvae, Carcinus maenas et Cyathura carinata, et par deux espèces exclusives, M. arenaria et Abra tenuis. L'assemblage II n'est constitué d'aucune espèce caractéristique, mais d'une partie des espèces présentes dans l'assemblage I et dans les assemblages III et IV. Il présente ainsi de nombreuses espèces constantes ou régulières, mais aucune préférente, élective ou exclusive. L'assemblage III se caractérise par un faible nombre d'espèces constantes et par deux espèces préférentes, L. conchilega, espèce commune, et Scoloplos armiger, espèce constante. L'assemblage IV est constitué d'un grand nombre d'espèces communes ou constantes. Il se particularise par la présence de C. arenarium, Eurydice pulchra, constantes dans cet assemblage uniquement, et par la présence d'une espèce régulière, Bathyporeia pilosa. L'espèce caractérisant le mieux cet assemblage est B. sarsi, qui est régulière et préférente. L'assemblage V, le plus externe au site, est constitué de quelques espèces communes ou constantes, qu'il partage surtout avec l'assemblage IV. Il se caractérise par trois taxa préférents, A. brachiata, les némertes et Scolelepis squamata ainsi que par une espèce exclusive, B. elegans.
3.1.3 Comparaison du macrozoobenthos entre 1985 et 2000
Concernant les 80 taxa recensés sur les deux périodes (Tableau 1), 40 % sont communs aux deux échantillonnages. Les taxa observés uniquement une année sont à 90 % des espèces rares ayant une fréquence d'occurrence inférieure à 5 % et présentant de très faibles abondances.
Les distributions spatiales des assemblages d'espèces de 1985 (Fig. 3a) et de 2000 (Fig. 3b) montrent une répartition relative inchangée, nous permettant d'associer les assemblages I, II et III de 1985 à ceux de 2000. Les deux assemblages les plus externes du flanc ouest et de l'estran central (IV et V) ont une répartition relative ne nous permettant pas de les distinguer d'une année à l'autre. Les résultats de la classification ascendante hiérarchique (Fig. 4) nous permettent de déterminer les assemblages les plus proches et d'identifier les correspondances entre 1985 et 2000 des assemblages IV et V.
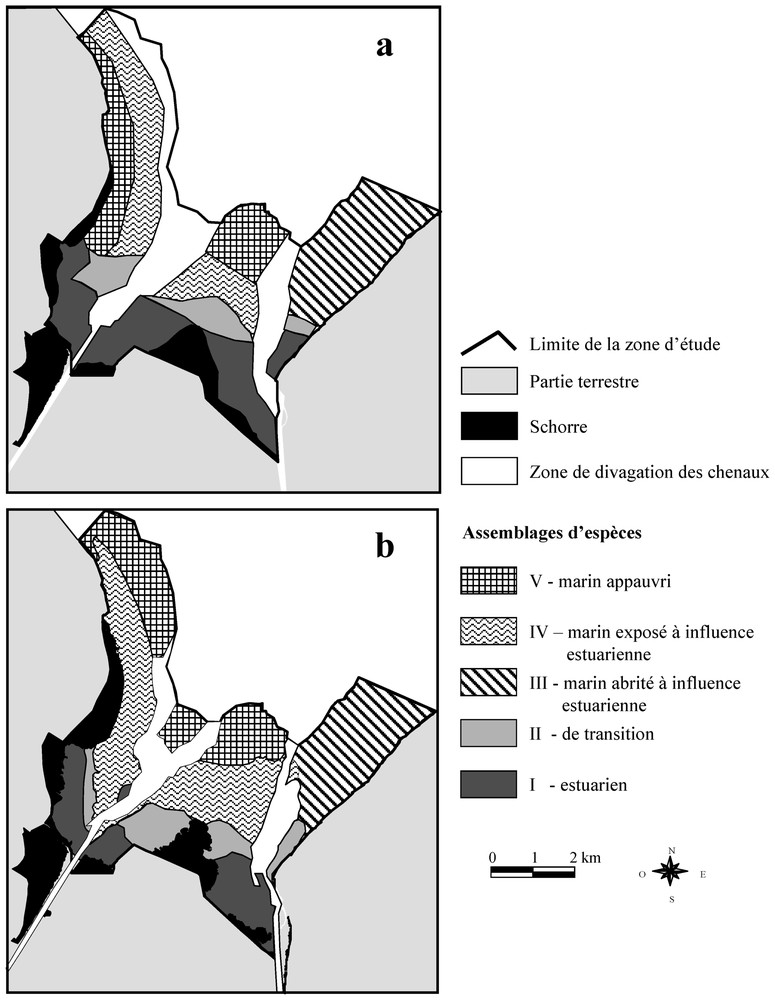
(a) Répartition des assemblages d'espèces en 1985 (d'après [7]). (b) Répartition des assemblages d'espèces en 2000.
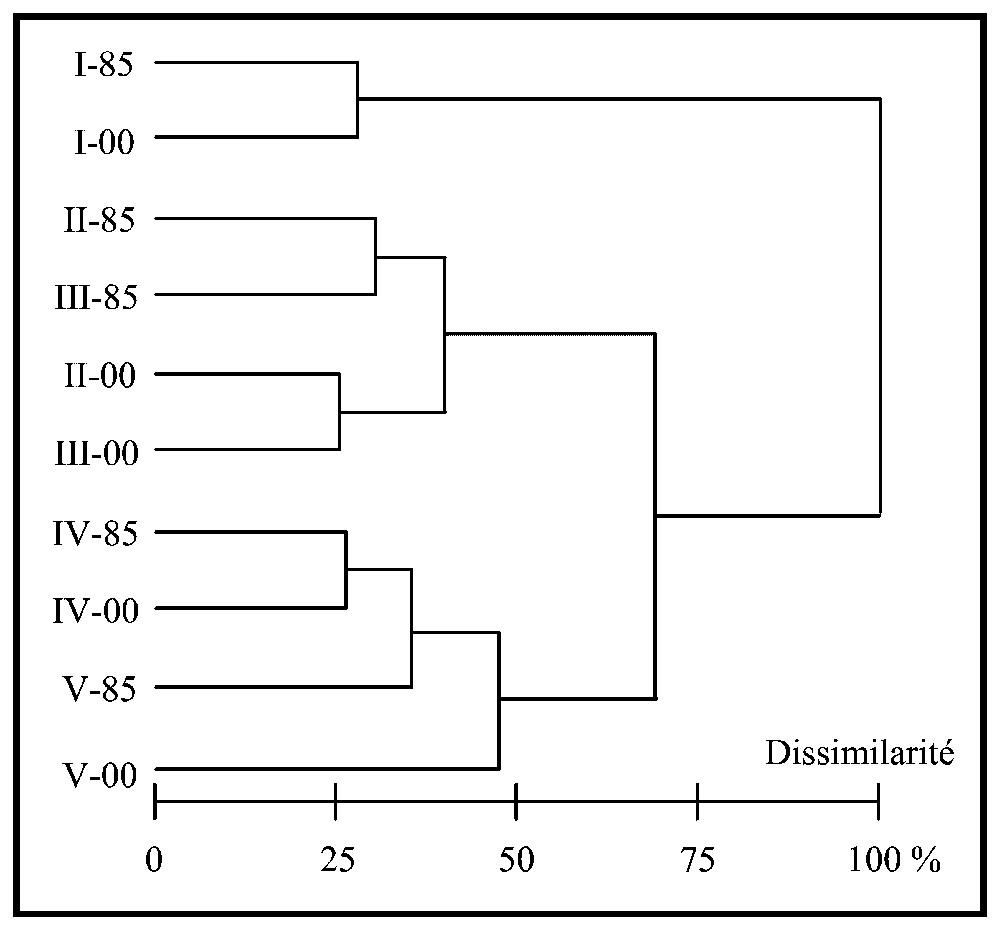
Classification ascendante hiérarchique des assemblages d'espèces de 1985 (notés -85) et de 2000 (notés -00) réalisée sur les abondances taxonomiques moyennes de chaque assemblage (distance euclidienne et méthode de classification de Ward). La dissimilarité maximale observée (100%) est de 1,9344. Le codage des assemblages de 1985 a été réalisé a posteriori. Les assemblages ont été ordonnés (de I à V et de 85 à 00) afin de faciliter les comparaisons.
Cette classification a été réalisée à partir d'une analyse factorielle des correspondances sur les 25 principaux taxa décrivant les assemblages en 1985 et 2000 suivie d'une classification ascendante hiérarchique effectuée à partir des coordonnées factorielles des sept premiers axes, représentant 98 % de l'inertie totale. La dissimilarité maximale observée entre les différents assemblages d'espèces de 1985 et 2000 concerne les assemblages I des deux années par rapport à tous les autres.
Les assemblages présentant les structures les plus semblables entre les deux années sont les assemblages I et IV avec respectivement 28 % et 26 % de dissimilarité d'une année sur l'autre. Les assemblages II et III présentent 40 % de dissimilarité d'une année sur l'autre. Ces assemblages ont des structures plus proches la même année que d'une année à l'autre. L'assemblage V est celui ayant le plus évolué au cours du temps. Il présente une dissimilarité de 47 % entre 1985 et 2000. Il est plus proche de l'assemblage IV en 1985 (35 % de dissimilarité) qu'en 2000 (47 % de dissimilarité).
La comparaison des taxa, au moins constants ou préférents, des assemblages de 1985 et de 2000 (Tableau 3), fait apparaı̂tre des différences principalement dans les assemblages II, IV et V. Les différences de l'assemblage IV s'expliquent par le nombre important de taxa dont la valeur de l'indice de constance est proche du changement de classe, passant de 1985 à 2000 de commun à constant ou inversement. Il n'y a donc pas de différence notable d'une année sur l'autre. L'assemblage V ne conserve qu'une espèce caractéristique, B. elegans, et acquiert d'autres espèces accompagnatrices. L'assemblage II conserve les taxa présents en 1985, auxquels s'ajoutent des espèces accompagnatrices, qui étaient présentes uniquement en 1985 dans l'assemblage I, et qui se retrouvent, en 2000, dans les assemblages I et II.
Comparaison, dans chaque assemblage d'espèces, de la présence des taxa caractéristiques utilisés pour la classification ascendante hiérarchique (Fig. 3) en 1985 et 2000. Les cellules grisées indiquent la présence des taxa ayant un indice de constance (C) supérieur à 50% et/ou un indice de fréquence (F) supérieur à 66,7%
La distribution spatiale des assemblages d'espèces entre 1985 et 2000 (Fig. 3 a et b, respectivement), montre une régression de l'assemblage I vers le sud accompagnée d'un déplacement de l'assemblage II. L'assemblage III occupe le même espace. L'assemblage IV progresse en direction du sud, au centre et à l'ouest de la baie, en occupant l'espace laissé vacant par l'assemblage II. Au centre de la baie, il s'étend aussi vers le nord. Sur la partie ouest de la baie, l'assemblage IV occupe l'espace où se trouvait l'assemblage V, en haute slikke. Cet assemblage se trouve toutefois présent dans la partie nord-ouest, où il était absent.
4 Discussion
La faune macrozoobenthique de la baie des Veys est similaire à celle observée dans l'aire de répartition géographique boréo-atlantique. Elle s'organise autour d'une « communauté à Macoma » [23], qui se retrouve, à quelques variations spécifiques près, sur la plupart des estuaires de cette région, comme ceux du Nord de la France [3,5,7,19,24], mais aussi de l'Europe [25], dans les ı̂les Britanniques [26] et en Amérique du Nord [27–29].
Dans la baie des Veys, le macrozoobenthos se distribue en cinq assemblages d'espèces s'organisant selon un gradient terre–mer associé à une dissymétrie est–ouest.
D'après la typologie des habitats marins proposée par Picton et Costello [30], les assemblages d'espèces I et II sont constitués d'espèces caractérisant l'habitat qu'ils appellent : biotope à « H. diversicolor et M. balthica dans un estran à vases sableuses ». L'assemblage I correspond au sub-biotope à « H. diversicolor, M. balthica et M. arenaria dans un estran à vases sableuses ». L'assemblage II est un assemblage de transition entre le I et le III et IV. Bien que ne présentant pas d'espèces discriminantes, il est constitué de la majorité des espèces accompagnatrices que l'on retrouve dans ces trois assemblages. Il correspond au sub-biotope à « H. diversicolor, M. balthica et A. marina dans un estran à sable vaseux ou vases sableuses ».
Sur le flanc est, l'assemblage III se caractérise principalement par l'importance de L. conchilega et S. armiger, espèces caractérisant le biotope à « L. conchilega dense dans un bas niveau d'estran sableux ». Sur le flanc ouest et l'estran central, l'assemblage IV correspond à un complexe de biotope d'« estran à sable vaseux » comprenant principalement un biotope à « polychètes et C. edule dans un estran à sable fin ou à sable vaseux » sur les plus bas niveaux et un biotope à « B. pilosa et Corophium spp. dans un haut niveau d'estran à sable fin légèrement vaseux ». La distinction entre ces deux biotopes n'est toutefois pas apparue clairement lors de la classification des stations et ne nous permet pas de les distinguer spatialement.
Le dernier assemblage, désigné par le numéro V, le plus externe au site, présente des espèces caractérisant le biotope à « amphipodes fouisseurs et polychètes dans un estran à sable propre ». Bien que nous ayons observé que très peu de Pontocrates spp., il contient principalement des espèces caractérisant le sub-biotope à « amphipodes fouisseurs Pontocrates spp. et Bathyporeia spp. dans un estran à sable propre ».
La distribution des biotopes décrite en 2000 est assez similaire à celle de 1985, avec toutefois une évolution marquée de leurs frontières.
Les différences entre les listes des taxa observés en 1985 et en 2000 concernent uniquement les espèces les moins représentées et n'indiquent pas de modification dans la composition spécifique du macrozoobenthos de la baie des Veys. Toutes les espèces présentent en 2000 avaient déjà été décrites sur ce site à l'exception de Pontocrates altamarinus. Ces différences mineures pourraient être dues à la stratégie d'échantillonnage. Depuis le début des études macrozoobenthiques sur ce site, il n'a pas été observé de modification de la biodiversité spécifique.
Globalement, nous sommes en présence de la même répartition spatiale organisée en cinq types différents d'assemblages d'espèces. L'assemblage I, de type estuarien, ne se modifie pas, mais régresse spatialement. Le II, en tant qu'assemblage de transition, reflète la composition des assemblages I, III et IV de chaque année, les changements observés provenant des modifications des assemblages III et IV. Il comprend plus d'espèces accompagnatrices du I en 2000 qu'en 1985. Ceci peut s'expliquer par le changement de statut de certaines zones, qui sont passées de I à II.
Le flanc oriental de la baie, accueillant l'assemblage III, a toujours constitué une entité parfaitement délimitée par l'abri des roches de Grandcamp au nord et le chenal d'Isigny à l'ouest, sans grande transformation morphologique avant 1985. Le chenal d'Isigny constitue une barrière hydrodynamique efficace, qui confine les caractéristiques du macrozoobenthos local en limitant le transit larvaire latéral [19,31]. De ce fait, les perturbations apportées par la rapide extension des concessions conchylicoles ont également été confinées. L'assemblage originel à S. armiger et U. poseidonis, qui se rapproche d'un biotope à « polychètes et C. edule dans un estran à sable fin ou à sable vaseux », [30] était parfaitement homogène sur cette zone, exceptions faites de sa partie sud, à tendance estuarienne, et d'une frange des plus bas niveaux, où apparaissaient quelques espèces marine typiques (A. brachiata et Sthenelais boa). En 2000, si le rôle du chenal d'Isigny et des roches de Grandcamp demeure inchangé, le macrozoobenthos a subi d'importantes modifications. L. conchilega et sa faune associée se sont développés en situation atypique, c'est-à-dire en moyenne-haute plage plutôt que dans les bas niveaux où il est habituellement décrit [30] : le peuplement typique à S. armiger et U. poseidonis devient résiduel, limité au centre de la zone considérée. Le rôle majeur des installations conchylicoles dans cette transformation est appuyé par les travaux de Sornin [32] et Ropert [33], qui font état d'un envasement conséquent associé aux rangs de tables à huı̂tres, accompagné en baie des Veys par une prolifération rapide de L. conchilega [33].
En ce qui concerne le centre de la baie, il n'est plus abrité au nord par un corps sableux central limitant l'influence marine. L'assemblage IV qui lui est associé n'a pas changé, mais s'étend plus au sud en 2000. Le centre du site est de ce fait devenu cénotiquement plus homogène et a perdu sa caractéristique de zone de transition, au profit d'un caractère marin plus prononcé.
L'assemblage V se diversifie, se développe et devient plus structuré (plus d'espèces communes entre les stations). Spatialement, il s'homogénéise. D'abord situé en haute-moyenne plage à l'ouest de la baie, cette zone a été colonisée rapidement par le schorre à S. anglica ; cet assemblage à influence marine prononcée s'est ensuite reconstitué dans de plus bas niveaux sur l'estran nouvellement apparu, en raison du déplacement des chenaux vers l'est.
Le milieu des années 1980 marque ainsi un changement dans l'évolution de la baie des Veys. De 1974 à 1985 [7,8,11], l'évolution cénotique intertidale a été progressive à l'ouest, et se poursuit de 1985 à 2000 conformément au modèle prédictif. Au centre, l'intrusion marine a pris le pas sur le gradient sud–nord, estuarien-marin, ce qui nuance l'application locale de ce modèle. Le flanc est, qui n'a pas subit d'évolution notable jusqu'en 1985, est l'objet de profondes modifications, conformément au modèle, avec cependant une caractérisation estuarienne atténuée dans sa partie la plus au sud ; dans le maintien de conditions naturelles, cette évolution locale n'aurait pas dû avoir lieu et le modèle nous donne l'opportunité de suivre une modification essentiellement due à l'impact d'un aménagement, celui de la zone conchylicole du flanc est.
La prise en compte du modèle d'évolution du macrozoobenthos et des nuances que nous avons apportées permet d'envisager l'évolution future de la zone intertidale. D'ouest en est, l'intrusion marine occidentale a atteint le fond de la baie et ne devrait guère s'étendre davantage, tandis que la progression du schorre à S. anglica continuera de réduire l'espace estuarien. Au centre, les espèces macrozoobenthiques estuariennes seront repoussées par la progression marine en cours. Le faciès originel du flanc est, à S. armiger et U. poseidonis, tendra à disparaı̂tre face à la pression envahissante de L. conchilega, mais avec une cinétique supérieure à celle qui affecte la transformation du reste de la baie. Le suivi du flanc est nécessiterait donc un pas de temps plus court pour mieux comprendre une évolution plus rapide, qui est fonction de l'aménagement conchylicole local. Une éventuelle comparaison de son évolution macrozoobenthique avec celle d'autres sites estuariens est rendue difficile face aux caractères originaux présentés par la baie des Veys : c'est un espace intertidal étendu, plus vaste que celui de l'estuaire de la Seine, mais alimenté en eau douce par de petits débits, singulièrement orienté vers le nord-nord-est, possédant de vastes parcs à huı̂tres ainsi qu'une population proliférante remarquable de L. conchilega et qui profite d'une acquisition de données environnementales continue depuis plus de 30 ans. La baie des Veys est à notre connaissance le seul site profitant d'une description évolutive à long terme appliquée à la fois à un milieu naturel et à un site littoral aménagé.
Remerciements
Cette étude a bénéficié des financements de la Réserve naturelle de Beauguillot, du Parc naturel régional des Marais du Bessin et du Cotentin et de la direction régionale de l'environnement de Basse-Normandie.