

Abridged English version
All the recordings were made in an embanked marsh on the French Atlantic coast, near La Rochelle. These earthen ponds were built for various productions: salt, oysters, fish... Their depth does not exceed one metre and their surface area is commonly between 1000 and 5000 m2. These coastal ponds usually communicate with the sea through sluice gates and they have an intermittent marine water supply. This periodic communication with the sea enables them to receive a seasonal supply of larvae and juveniles of various sea fish, as well as benthic invertebrates. Practically all fish species found in a European macrotidal salt marsh can enter this kind of habitat and colonise it, at least temporarily, from the end of spring to the beginning of winter. That is the case of the following species: common sole, Senegalese sole, plaice, European sea bass, gilthead sea bream, European eel, mullets and gobiids. However, very few of them are able to survive during winter. Among them we can mention: the European eel (Anguilla anguilla L.) and some species of Gobiidae.
Sounds were detected with a hydrophone (Bruël and Kjaer, type 8101). The hydrophone was placed in a pond, one meter from the bank. The sounds were amplified with Bruël and Kjaer measuring amplifier (type 2610) and recorded with a Nagra IV SJS magnetic tape recorder. Sounds were analysed using a Tektronix 2622 analyser and its IP analysis software.
The recorded sounds were generally a very regular succession of a single pulse, but sometimes a series of sounds composed of one, two, three or four successive and very close pulses was heard. Their temporal characteristics are summarised in Table 1. All these sound series were unceasingly emitted during time periods of several minutes. Sonograms of these recorded sounds show that they seem to have a harmonic structure, with most of their energy lying below 1500 Hz. The frequency spectra of these sounds (Fig. 1) are characterised in the range of 0–2000 Hz by four frequency bands: 120–350, 520–760, 895–1130 and 1280–1560 Hz, with the greatest energy on 280 and 350 Hz (64.1 dB re. 1 μPa rms).
Caractéristiques temporelles des sons enregistrés exprimées en ms
| Durée pulse 1 | Durée pulse 2 | Durée pulse 3 | Durée pulse 4 | Période inter pulse 1–2 | Période inter pulse 2–3 | Période inter pulse 3–4 | Durée du son | Intervalle entre deux sons | |
| Son à 4 pulses | |||||||||
| Nombre | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 11 |
| Moyenne | 9,39 | 9,57 | 9,75 | 9,45 | 52,82 | 71,02 | 88,64 | 221,43 | 1134,30 |
| S.E. | 1,11 | 1,27 | 1,37 | 1,45 | 0,32 | 0,60 | 1,21 | 1,74 | 11,04 |
| Son à 3 pulses | |||||||||
| Nombre | 19 | 19 | 19 | 19 | 19 | 19 | 14 | ||
| Moyenne | 10,45 | 10,16 | 10,39 | 52,09 | 70,81 | 133,17 | 908,05 | ||
| S.E. | 0,98 | 1,35 | 0,90 | 0,37 | 0,62 | 1,14 | 15,27 | ||
| Son à 2 pulses | |||||||||
| Nombre | 18 | 18 | 18 | 18 | 16 | ||||
| Moyenne | 13,91 | 13,26 | 60,26 | 73,56 | 585,39 | ||||
| S.E. | 3,08 | 2,56 | 0,91 | 2,75 | 12,48 | ||||
| Son à 1 pulse | |||||||||
| Nombre | 54 | 53 | |||||||
| Moyenne | 7,58 | 100,19 | |||||||
| S.E. | 0,55 | 3,18 |
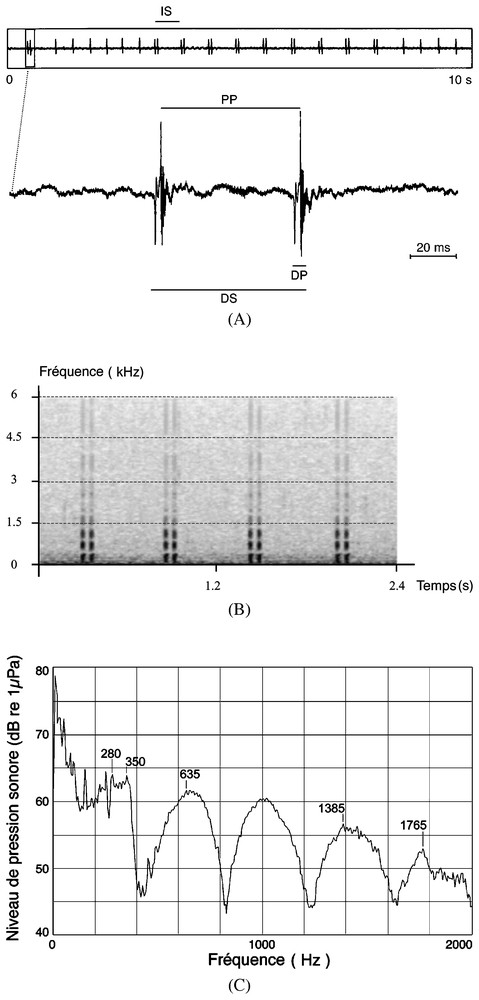
Caractéristiques acoustiques d'une émission sonore composée d'une succession de sons comprenant un ou deux pulses (14–15 h, 16/02/94). (A) Représentation oscillographique d'une émission sonore dans deux échelles de temps (10 s et 195 ms) et indication des caractéristiques temporelles mesurées: DP, durée d'un pulse; PP, période des pulses; DS, durée d'un son; IS, intervalle entre deux sons. (B) Sonogramme (largeur du filtre: 25 Hz). (C) Spectre de fréquence (spectre moyenné sur 30 spectres instantanés).
These sound emissions varied according to the time period. Seasonal variability first of all, since in a year and a half only two sound emissions were heard and recorded (February 1994 and April 1996). Daily variability also, since the temporal patterning of the sound emission varied between the beginning and the end of the day, starting with 1-pulsed sound series and finishing with 3- or 4-pulsed sound series.
Passive acoustic detection of sounds produced by fishes or invertebrates in natural conditions poses rapidly the problem of species identification. In our case, to identify the species producing the sound emissions we have just described, we disposed of three main sources of information: (1) a catalogue of recorded sounds of identified species, (2) a temporal patterning of the pulses, giving a very particular sound, and (3) precise data on the habitat and environmental conditions associated with the recorded sound emissions.
Amalgamating the first two sources of information and proceeding by deduction, we observed a close similarity between the studied sound emissions and those described for Anguilla rostrata. This species produced a long-continuated, low clucking that resembles put-put of an outboard motor. The definition of this American eel song underlines the originality of the temporal patterning of these sounds, which clearly distinguishes them from those produced by other fish species. The identification of these sounds to the ‘put-put’ of an outboard motor corresponds exactly to the sound image given by the regular single-pulse sounds. Such a similarity leads us to attribute the sound emissions recorded in a French Atlantic coastal pond to the European eel (Anguilla anguilla).
1 Introduction
Le premier enregistrement des émissions sonores analysées dans cette étude est dû au hasard. L'enregistrement initialement programmé concernait des mesures de bruits ambiants, dans des marais salés, durant une période de temps calme, pour une recherche sur les interrelations entre les poissons et les événements météorologiques [1,2]. Ce premier enregistrement a été suivi d'une écoute passive, effectuée chaque mois durant plus d'un an, dont les seuls résultats furent la détection de ces émissions sonores en février 1994, puis en avril 1996.
Malgré l'importante compilation réunie sur les sons émis par de nombreuses espèces de poissons de l'Atlantique nord [3], il est souvent difficile de reconnaı̂tre en mer la signature acoustique de telle ou telle espèce de poissons, et ceci pour plusieurs raisons. Tout d'abord, la grande ressemblance des spectres de fréquence des sons émis par les poissons, où toute l'énergie acoustique se situe sur les basses fréquences, ce qui génère une dominance de sons graves et réduit les possibilités d'une identification basée sur des fréquences particulières. Ensuite, parce que les sons attribués à telle ou telle espèce, ayant été le plus souvent enregistrés en aquarium et après un choc électrique [4], représentent essentiellement des signaux d'alarme ou de détresse, ce qui restreint de beaucoup l'éventail des productions sonores associées à des comportements particuliers (reproduction, territorialité, identification spécifique, alimentation…). Aussi, les études les plus récentes qui utilisent des observations faites en aquarium ont-elles essayé de placer le poisson dans des conditions de confort proches des conditions naturelles. Une telle approche a ainsi permis de corréler des émissions sonores à des comportements particuliers : alimentation [5,6], intimidation [7–10], reproduction [10–12] ou défense du territoire [13].
Même si la composition fréquentielle de ces émissions sonores apporte peu d'éléments à la reconnaissance interspécifique, leurs modulations temporelles ont, au contraire, assez vite attiré l'attention sur leur potentiel de discrimination interspécifique [14]. Les caractéristiques temporelles du signal sont ainsi devenues peu à peu les principaux caractères d'identification des espèces productrices de sons [15–17]. En se fondant sur ces acquis, notre étude avait pour objectif de comparer les signaux recueillis en marais maritime à ceux d'espèces connues, pour tenter d'en identifier l'espèce émettrice.
2 Présentation de la zone d'étude
Tous les enregistrements ont été effectués dans un marais salé endigué, situé près de La Rochelle, sur la côte atlantique française, entre 1994 et 1996. Ces marais sont constitués de bassins de terre, qui furent construits avec des intentions de productions diverses : production de sel, d'huı̂tres, de poissons…). Leur profondeur n'excède pas 1 m et leur surface s'étend, le plus souvent, entre 1000 et 5000 m2. Ces bassins de terre communiquent généralement avec la mer par des vannes et bénéficient d'un apport intermittent d'eau de mer [18]. Cette communication périodique avec la mer les rend aptes à accueillir, selon les saisons, des larves et des juvéniles de poissons aussi bien que d'invertébrés benthiques. En fait, toutes les espèces de poissons qui se rencontrent dans les marais salés intertidaux des côtes d'Europe [19] peuvent pénétrer dans ce type d'habitat et le coloniser, au moins temporairement, de la fin du printemps au début de l'hiver. C'est le cas des espèces suivantes : sole commune, sole sénégalaise, plie, bar, dorade royale, anguille européenne, ainsi que de plusieurs espèces de muges et de Gobiidés. Cependant, un très petit nombre d'entre elles sont capables de survivre aux conditions hivernales et parmi elles nous pouvons mentionner l'anguille européenne et deux ou trois espèces de Gobiidés.
L'enregistrement des émissions sonores étudiées a été réalisé dans l'un des petits bassins de terre du site expérimental du CREMA. Ce bassin, d'une superficie de 250 m2, a fait l'objet d'une pêche à la nasse durant 9 jours (juillet 1995), qui a permis la capture de cinq individus d'anguille, le plus grand mesurant 480 mm (180 g) et les quatre autres étant d'une taille moyenne comprise entre 200 et 400 mm [20]. Cet échantillonnage correspond bien au niveau des captures et à la distribution de taille des individus colonisant un marais endigué de la baie de Bourgneuf de 300 ha [21].
Dans ce bassin de 250 m2, des sons ont été détectés à deux reprises, en février 1994 et en avril 1996, au cours d'une série d'écoutes effectuées à différentes périodes : février 1994 (2 j), juillet 1995 (2 j), août 1995 (2 j), novembre 1995 (1 j), février 1996 (3 j), mars 1996 (4 j) et avril 1996 (3 j). Le 16 février 1994, les conditions météorologiques étaient les suivantes : vent faible (3 à 8 m s−1), ciel couvert (irradiance de 10 à 65 mW cm−2), température assez basse (5 à 11 °C dans l'air, 6 à 7,5 °C dans l'eau), pression atmosphérique élevée (1016 mbar) et les enregistrements ont été réalisés entre 10 h et 19 h, les émissions sonores étant continues. Le lendemain matin, elles avaient cessé. Le 15 avril 1996, les conditions météorologiques étaient les suivantes : vent faible (3 à 6 m s−1), ciel couvert (irradiance de 10 à 90 mW cm−2), température (16 °C dans l'air, 17–18 °C dans l'eau) et pression atmosphérique élevée (1024 mbar). Les enregistrements ont été effectués entre 17 h et 19 h où l'émission était continue, mais perturbée par le bruit d'écoulement d'eau du trop plein du bassin. Le lendemain matin, l'émission sonore avait cessé.
3 Enregistrement et analyses des sons
Les sons sont détectés par un hydrophone (Bruël & Kjaer, type 8101) d'une sensibilité de −184 dB re. 1 V μPa−1 et ayant une réponse en fréquence plate jusqu'à 120 kHz. Les émissions sonores recueillies sur cet hydrophone sont calibrées en utilisant le calibreur Bruël & Kjaer (type 4223), qui délivre, dans le coupleur de l'hydrophone 8101, un niveau de pression sonore de 157 dB re. 1 μPa sur la fréquence de 250 Hz. L'hydrophone est immergé dans le bassin à 1 m de la berge. Les sons sont amplifiés par un amplificateur de mesure (Bruël & Kjaer, type 2610) et enregistrés sur la bande d'un enregistreur magnétique Nagra IV SJS.
Les sons sont analysés à l'aide d'un analyseur Tektronix 2622 et du logiciel IP qui lui est associé (analyses temporelle et fréquentielle du signal). Après avoir été numérisés grâce à une carte d'acquisition 16-bit équipée d'un filtre anti-repliement, les signaux sont traités par le logiciel SYNTANA [22] pour en faire une analyse sonographique. Seules les caractéristiques temporelles usuelles [23] ont été mesurées (Fig. 1A) :
- – durée du pulse (DP) et du son (DS) en millisecondes (ms) : mesure de l'intervalle de temps entre le début d'un pulse ou d'un son et la fin de ceux ci ;
- – intervalle entre deux sons (IS) en millisecondes (ms) : intervalle de temps entre le pic principal du premier pulse d'un son et le pic principal du premier pulse du son suivant ;
- – périodes inter pulses 1–2, 2–3, 3–4 en millisecondes (ms) : intervalle de temps entre les principaux pics de deux pulses successifs.
4 Résultats
Les sons enregistrés se présentent, soit comme une succession très régulière d'un son composé d'un seul pulse, soit d'une succession tout aussi régulière de sons composés en alternance ou régulièrement de un, deux (Fig. 1A), trois ou quatre pulses très rapprochés. Ces émissions sonores sont produites sans interruption sur des durées de plusieurs dizaines de minutes. Leurs caractéristiques temporelles sont regroupées dans le Tableau 1. Leur sonogramme (Fig. 1B) montre une structure harmonique avec un maximum d'énergie acoustique placée sur des fréquences inférieures à 1500 Hz. Les analyses de spectres ont été réalisées sur des moyennes additives de 30 spectres instantanés correspondant à 30 simples ou doubles pulses capturés par la fonction trigger. Les spectres de fréquence de ces émissions sonores sont caractérisés, entre 0 et 2000 Hz, par quatre bandes de fréquences (Fig. 1C) : 120–350, 520–760, 895–1130 et 1280–1560 Hz, avec un maximum d'énergie acoustique sur les fréquences comprises entre 280 et 350 Hz (64,1 dB re. 1 μPa rms).
Ces émissions sonores sont peu fréquentes, puisque nous n'avons pu les enregistrer que deux fois en un an et demi de prospection. Leur modulation varie parfois au cours de la journée. En février 1994, on a pu observer, en fin de matinée, des sons composés d'une succession régulière d'un seul pulse (Fig. 2), puis sont apparus, en début d'après midi, des sons où se mélangeaient des successions de un et deux pulses (Fig. 1A) et, en fin d'après midi, des séries de signaux à trois (Fig. 3) et quatre pulses (Fig. 4). En avril 1996, les sons étaient composés d'une succession régulière mais plus lente d'un seul pulse (Fig. 5).
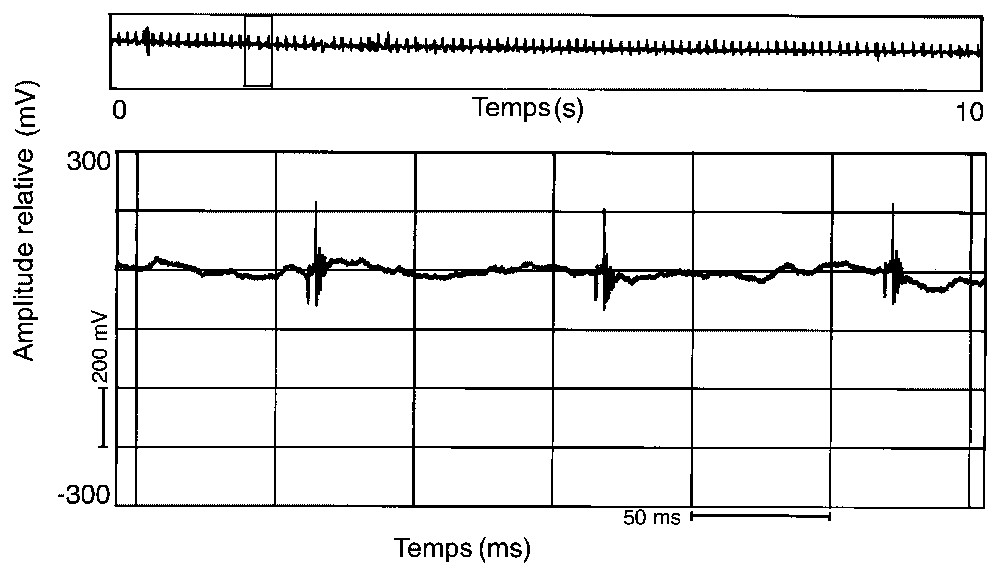
Représentation oscillographique d'une émission sonore constituée d'une succession de sons composés d'un seul pulse et donnant une image sonore de moteur hors bord (10–12 h, 16/02/1994).
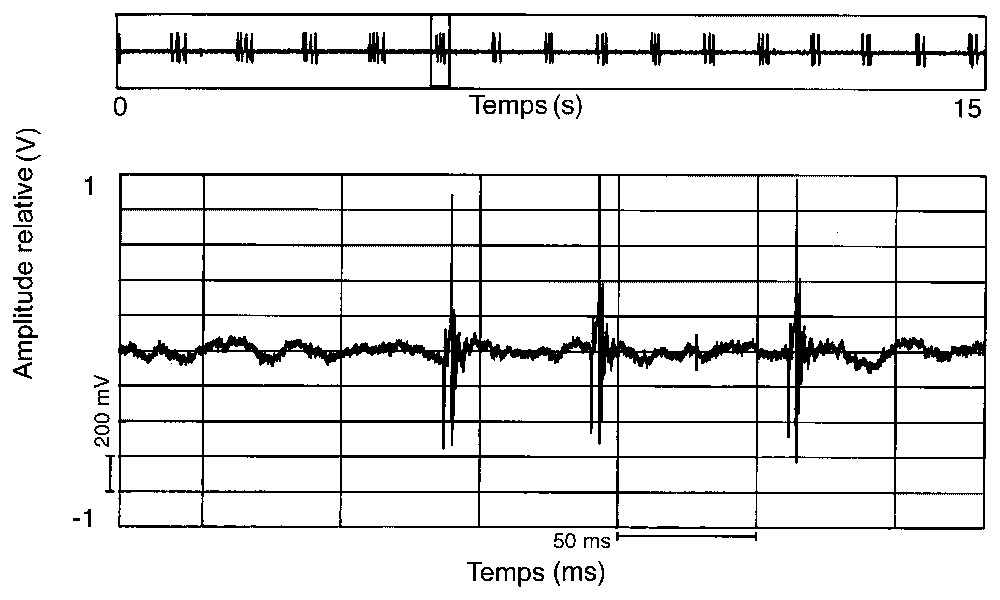
Représentation oscillographique d'une émission sonore constituée d'une succession de sons composés de trois pulses (17 h, 16/02/1994).
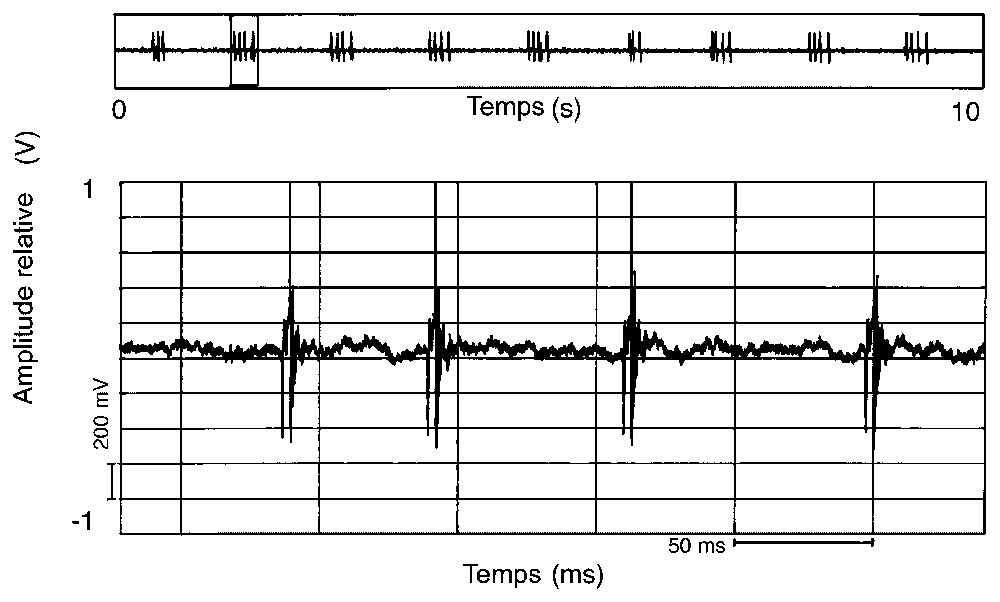
Représentation oscillographique d'une émission sonore constituée de sons composés de quatre pulses (19 h, 16/02/1994).

Représentation oscillographique de l'émission sonore enregistrée le 15 avril 1996 (18 h) et constitués d'une succession de sons composés d'un seul pulse. Le signal est filtré à travers un filtre passe haut de 500 Hz pour réduire les interférences liées au bruit d'écoulement d'eau en sortie du bassin.
5 Discussion
La détection et l'enregistrement de sons émis par des poissons ou des invertébrés dans leur environnement naturel et en l'absence de contrôle visuel posent rapidement le problème de l'identification des espèces productrices de ces signaux sonores [4]. Dans notre cas, en raison de la turbidité importante de l'eau, aucun contrôle par caméra sous-marine n'a pu être réalisé. Pour tenter d'identifier l'espèce productrice des sons que nous venons de décrire, nous disposions de trois sources principales d'informations : (1) un catalogue de sons enregistrés à partir d'espèces bien identifiées et originaires de la partie nord-ouest de l'océan Atlantique [3], (2) une modulation temporelle du signal en séries de pulses unique ou multiple donnant un son très particulier et (3) une indication très précise de l'habitat et des conditions environnementales associées aux périodes de détection et d'enregistrement. En croisant les deux premières sources d'information, on ne peut manquer de relever la grande similitude des émissions sonores étudiées avec celles décrites chez l'anguille américaine.
En effet, l'anguille américaine est connue pour produire deux types de sons : un bruit sourd de cognement et un long et continu pétillement qui ressemble au put-put d'un moteur hors bord. Ce pétillement a été entendu dans l'air en plusieurs occasions, en particulier durant les déplacements terrestres de ce poisson lors de nuits obscures et humides. Cependant, il n'a jamais été enregistré sous l'eau et on ne dispose pas non plus d'enregistrements aériens ayant été analysés [3,24].
L'identification de ce deuxième type de son (pétillement) au bruit d'un moteur hors-bord correspond exactement à l'image sonore produite par les séries continues composées d'un seul pulse que nous avons enregistrée dans les marais (Figs. 2 et 5). De plus, ces sons ont une modulation temporelle exceptionnelle et jamais rencontrée chez les autres espèces de poissons productrices de sons. La régularité bien particulière de ces émissions sonores, composées d'un ou plusieurs pulses très régulièrement espacés (Tableau 1) et émises de façon continue durant plusieurs dizaine de minutes, n'a d'équivalent que chez l'anguille américaine, ce qui oriente notre identification de l'espèce émettrice vers l'anguille européenne (Anguilla anguilla). En effet, si d'autres espèces de poissons vivant dans les marais côtiers, en particulier les Gobiidés, sont capables d'émissions sonores, celles-ci sont toujours irrégulières, discontinues dans le temps et composées de trains de pulses alternant avec des périodes plus ou moins prolongées de silence [25,26].
Enfin, ce rapprochement se voit encore conforté par le lieu et la période des enregistrements, fin de l'hiver–début du printemps. Comme nous l'avons indiqué, les bassins d'enregistrement peuvent accueillir bon nombre d'espèces de poissons côtiers durant une partie de l'année [19]. Mais, à la fin de l'hiver, ne sont présentes dans ces bassins que les espèces ayant pu survivre aux conditions de températures hivernales, ce qui réduit la diversité du peuplement à quelques Gobiidés et à l'anguille.
Pour l'anguille américaine, on dispose d'une mise en relation entre les sons de type pétillement continu et les migrations terrestres nocturnes de l'espèce [24]. On peut donc supposer une relation identique pour l'anguille européenne, chez laquelle les sons de pétillement continu, enregistrés sous l'eau, seraient des signaux indicateurs ou annonciateurs d'une sortie imminente du bassin par reptation sur la berge. Ceci est conforté par l'absence de ces émissions sonores lors des écoutes réalisées le lendemain matin. De tels signaux jouent-ils un rôle dans la synchronisation des départs de plusieurs individus ? Pourquoi semblent-ils se poursuivre en milieu aérien au risque d'une prédation accrue ? Il est évident que nombre d'investigations restent à mener à bien pour parvenir à une compréhension précise et claire de la fonction biologique attachée à ces émissions sonores.
Ainsi, les émissions sonores analysées, en raison de leur modulation temporelle, de leur continuité sur des périodes de plusieurs dizaines de minutes, des caractéristiques du milieu et de la saison où les enregistrements ont été réalisés, nous paraissent devoir être attribuées à l'anguille européenne (Anguilla anguilla), en attendant qu'on puisse en obtenir une confirmation indubitable en aquarium, ce que nous n'avons pu réussir à faire jusqu'à présent, pas plus que ce ne fut le cas pour l'anguille américaine (Anguilla rostrata) [24].