

Abridged English version
Since the works of Sernander published in 1906, few exhaustive studies have been carried out on seed dispersal by ants in Europe. Moreover, most of them concern non-mutualistic ants of the Mediterranean area, while temperate forest ecosystems are largely ignored. Nevertheless, 24 to 30% of European forest plants are myrmecochorous species, making ants a major force in the spread of plants.
Understanding the determinisms and consequences of seed dispersal by ants is of outstanding importance, since it contributes to shape biodiversity patterns. On the one hand, the morphological traits of seeds are known to influence ants' behaviour, and consequently are a fundamental determinant of the ability of the plants to be dispersed by the insects. On the other hand, ant-dispersed seeds are protected from granivorous species, are transported away from parent plants that may inhibit the growth of seedlings, and are more prompt to colonize distant and new sites than barochorous species. A consequence of seed dispersal by ants should be a large benefit for myrmecochorous plants, i.e. the expansion of the plants' range and the improvement of their survival in the field.
In this study, we aim at (1) establishing an experimental hierarchy among 33 vascular plants collected in forest according to their ability to have their seeds dispersed by the ant Myrmica ruginodis, (2) evaluating the role of the seed characteristics (weight, length …) in the efficiency of dispersal by ants, (3) comparing plant dispersal ability with their frequency in the field.
The study was carried out in the Picardie region, in northwestern France. Myrmica ruginodis was chosen for the present study because this ant species is reportedly one of the most active seed dispersers in the temperate forests of Europe and is one of the dominant forest ant species in the region. Seeds of 33 plant species were collected at maturity in different forest sites and cooled immediately after collection and during storage to keep them fresh. Eighteen colonies of Myrmica ruginodis containing a minimum of 500 workers were collected in different forest sites. Their nests were dismantled and placed in experimental trays in the laboratory. Before each experimental test, a set of 10 seeds was defrosted and placed 15 cm away from ant nests. Five repetitions of this test were made for each plant species at different dates in July 2006 and using different colonies. The number of seeds removed by ants was monitored at , , , and after the first contact between a seed and an ant.
Temporal variations of seed removal for each plant species were fitted with Lowess curves using the software program ADE-4. Differences in seed removal rates among plant species were compared using Kruskal–Wallis tests. In order to examine the relationships between the seed characteristics and the rates of seed removal by ants, a set of seed traits was extracted from a European plant trait data base: flowering phenology, diaspore and germinule weight, thickness, width, and length. The cover and frequency of plant species in the field were extracted from a previous study carried out in Compiegne forest, the third largest plain forest in France. Spearman correlation analyses were used to assess the association between experimental dispersal rates, plant traits and plant frequency in the field.
Results showed that the elaiosome-bearing seeds (species Chelidonium majus, Colchicum autumnale, Luzula forsteri, Viola reichenbachiana, Luzula multiflora, Melica uniflora) and the small and light caryopses of the grasses (Holcus mollis, Agrostis capillaris, Deschampsia flexuosa, Poa trivialis, Holcus lanatus) were the most efficiently collected by ants. Chelidonium majus and Colchicum autumnale exhibited the highest seed removal rate: 1.67 seeds were collected per minute. The rate of seed removal was negatively correlated with diaspore length/width and length/thickness ratios and with germinule length and length/width ratio. No significant correlation was found between the rate of seed removal and the frequency of the plants in the field.
This study highlighted two types of dispersal strategy among forest plant species. The first strategy involves plants that produce an elaiosome and the mutualism-type behaviour of the ants. The second strategy is opportunistic and can be described as an outcome of seed predation. This opportunistic dispersal involves grasses with small and very light seeds that are easy for ants to carry, and the granivorous behaviour of the ants. The seed of the grasses does not have a specialised organ such as the elaiosome, but the whole seed containing interesting nutrients for ants is used to attract the insect. The main inconvenience of the opportunistic dispersal is the large quantity of seed gathered by ants and definitively sacrificed. The gradient of seed removal, negatively correlated with the size of the seeds, was interpreted as a trade-off between the risk for small seeds to be predated by ants and the benefit for heavy seeds to be dropped or lost along the way to the nest. Our results led us to reconsider the notion of myrmecochory, which is restricted to plants producing elaiosomes and/or attractive chemical compounds. Indeed, unspecialised seeds of grasses are as efficiently dispersed by ants as elaiosome-bearing seeds are. No correlation was found between experimental seed removal rate and plant frequency on the scale of the whole forest. This result suggests that other factors, such as local soil conditions or disturbances, may mask the role of ants and must be examined first to explain plant spread.
The quantity and the diversity of seeds potentially dispersed by Myrmica ruginodis indicate that this ant species plays a major role in plant diversity variations in the French forests. We suggest that the quality of the habitat and the nests of Myrmica ruginodis should be preserved during foresters' management activities by leaving fallen trees and branches on the forest floor.
1 Introduction
Depuis les travaux remarquables du botaniste Sernander publiés en 1906 [1], très peu d'études ont été menées de façon exhaustive sur la dispersion des graines par les fourmis en Europe. Les recherches expérimentales ou théoriques récentes ne mettent en jeu qu'un nombre très réduit de plantes, dépassant difficilement les trois espèces [2–7]. De plus, les seules études comparant un grand nombre de genres ou de familles de plantes sous l'angle de la myrmécochorie ne sont pas expérimentales [8]. En Europe, le champ des investigations scientifiques sur le transport des graines par les fourmis est encore plus réduit, car il concerne essentiellement les fourmis à régime strictement granivore et les plantes du Bassin méditerranéen [9–11]. Les relations de véritable mutualisme plantes–fourmis, via la présence d'élaiosomes ou de composés chimiques attractifs sur les graines [12], sont encore plus rarement explorées lorsqu'il s'agit d'écosystèmes forestiers tempérés [13,14].
Vingt-quatre pour cent des espèces végétales des forêts sont pourtant dispersés par les fourmis en Europe tempérée [15]. Ce chiffre s'élève à 30% pour les forêts tempérées de l'hémisphère nord [8]. Cela signifie que, sans les fourmis, au moins un quart des espèces herbacées des bois n'auraient plus le moyen de migrer efficacement et seraient réduites à la barochorie et/ou à la multiplication végétative. Les espèces de fourmis les plus actives dans la dispersion des graines comme les fourmis rousses des bois [16] étaient abondantes autrefois. En effet, l'utilisation de leur couvain pour nourrir les faisans d'élevage a été couramment pratiqué au cours des siècles passés [17]. Ces espèces, maintenant protégées en Europe, ont régressé à cause des activités humaines et sont absentes de nombreuses forêts [18,19]. Dans le Nord de la France, les genres Myrmica et Leptothorax sont les plus fréquemment observés en milieu forestier fermé [20] et sont les derniers animaux invertébrés capables d'assurer une véritable fonction de dispersion pour les plantes n'ayant pas de fruits charnus ou d'adaptations à l'anémochorie. Cela fait des fourmis une force majeure influençant la répartition spatiale des plantes [21] et de leurs pluies de graines [22]. De surcroît, les fourmis participent aussi à la survie des espèces végétales en enterrant leurs graines dans des microsites favorables, en les soustrayant aux prédateurs et aux intempéries et en minimisant la compétition entre plantules et géniteurs [21,23].
Comprendre le déterminisme et les conséquences de la myrmécochorie est crucial pour comprendre l'état actuel de la biodiversité, fortement dépendante des patrons spatiaux de la pluie de graines [22,24]. Chaque espèce végétale possède des traits de vie morphologiques, chimiques ou phénologiques propres, qui la prédisposent plus ou moins à être dispersée par les fourmis [12,25]. Ces traits seraient donc les déterminants principaux de leur capacité à la myrmécochorie. La conséquence serait un avantage sélectif pour les espèces myrmécochores, plus aptes à se déplacer et à coloniser de nouveaux sites favorables dans un environnement changeant.
Nous proposons de comparer expérimentalement la capacité de 33 espèces végétales à disperser leurs graines par la fourmi rouge des bois Myrmica ruginodis et de vérifier si elle favorise une large distribution des plantes adultes sur le terrain. Plus spécifiquement, nous cherchons à (1) établir un classement des espèces végétales en fonction leur capacité à la myrmécochorie, (2) évaluer le rôle des caractéristiques des graines (poids, longueur, etc.) dans la performance de collecte des graines par les fourmis, (3) comparer les performances expérimentales de dispersion des espèces végétales à leur fréquence sur le terrain.
2 Matériel et méthodes
2.1 Région d'étude
L'étude s'intéresse aux espèces végétales et aux fourmis des forêts de Picardie. Le climat de la région est de type océanique et comporte des influences continentales modérées [26]. Les températures moyennes mensuelles varient entre 3 et 19 °C et les précipitations moyennes mensuelles de 38 à 66 mm. L'altitude varie entre 0 et 290 m. Le substrat géologique correspond à des dépôts sédimentaires de craie blanche du Crétacé supérieur (environ la moitié ouest de la région) et des dépôts de l'Éocène inférieur et moyen et de l'Oligocène supérieur pour la partie sud-est de la région. Les forêts picardes sont, soit gérées en futaie ou plus rarement taillis sous futaie par l'Office national des forêts [27,28] pour les forêts domaniales (Compiègne dans l'Oise, Creuse et Crécy-en-Ponthieu dans la Somme) ou communales (Frémontiers dans la Somme), soit le plus souvent en taillis ou en futaie très irrégulière pour les forêts privées. Les essences dominantes des forêts sont le hêtre (Fagus sylvatica), les chênes pédonculé et sessile (Quercus robur et Quercus petraea), le charme (Carpinus betulus) et le pin sylvestre (Pinus sylvestris). Les détails sur les conditions écologiques caractérisant ces forêts sont disponibles dans diverses synthèses régionales et des études locales [28,29]. Les espèces de fourmis les plus fréquemment rencontrées à l'intérieur des parcelles forestières sont Myrmica ruginodis et Leptothorax nylanderi.
2.2 Choix des espèces
Plusieurs raisons ont motivé le choix des espèces végétales testées dans le cadre de cette étude. D'abord, Chelidonium majus a été choisie parce que ses graines possèdent un élaiosome très développé et sont connues pour être très utilisées par différentes espèces de fourmis, notamment forestières [1,4,16,19,25,30–32]. Cette qualité permet de les utiliser en tant que « standard » pour des comparaisons bibliographiques. Cependant, Chelidonium majus n'est pas une espèce strictement forestière ; c'est pourquoi nous avons choisi un jeu d'espèces plus typiquement forestières, dont la dispersion des graines par les fourmis est, soit connue, i.e. Allium ursinum, Anemone nemorosa, Carex pilulifera, Colchicum autumnale, Holcus lanatus, Hyacinthoides non-scripta, Luzula multiflora, Melica uniflora, Viola reichenbachiana [1,2,19,25,32–37], soit inconnue, i.e. Bromus racemosus, Capsella bursa-pastoris, Dactylis glomerata, Deschampsia flexuosa, Fragaria vesca, Milium effusum, Paris quadrifolia, Poa trivialis, Prunella vulgaris, Rumex crispus, Stachys sylvatica. D'autres espèces testées appartiennent à un genre connu pour contenir des espèces dispersées par les fourmis : Agrostis capillaris, Carex flacca, Carex pendula, Carex sylvatica, Euphorbia amygdaloides, Holcus mollis, Lamium galeobdolon, Luzula forsteri, Myosotis scorpioides, Primula elatior, Ranunculus repens [8,37]. Bien qu'anémochores, les graines de Betula pendula ont aussi été testées, car elles sont très abondantes sous les arbres géniteurs et pourraient fournir une source de nourriture conséquente pour des fourmis à régime même partiellement granivore.
L'espèce de fourmi rouge des bois Myrmica ruginodis Nyl. (Hyménoptère, Formicidés) a été sélectionnée pour l'étude parce que c'est l'une des deux fourmis les plus communes des forêts de la région. Par ailleurs, elle est connue pour être un disperseur de graines très efficace dans les forêts et les landes nord-européennes [7,14,34,38]. Cette espèce installe son nid dans des fragments de branches en décomposition sur le sol, sous la litière ou sous la mousse.
La nomenclature des espèces suit celle de Lambinon et al. (2004) pour les plantes [39] et de Bolton (1995) pour les fourmis [40].
2.3 Protocole expérimental
Les graines de 33 espèces végétales ont été récoltées à leur maturité sur différents sites forestiers de Picardie (Compiègne dans l'Oise, Villers-Bretonneux et Creuse dans la Somme, et Villers-Cotterêts dans l'Aisne), puis congelées dans des piluliers séparés à , dans un délai de 2 h suivant la récolte. Ceci permet de conserver la fraîcheur des graines, matures à différentes périodes de l'année, et de tester leur pouvoir attractif dans des conditions expérimentales identiques.
Dix-huit colonies de Myrmica ruginodis ont été récoltées en juin 2006 dans les forêts de Picardie (Compiègne, Villers-Bretonneux, Creuse). Chacun des nids de chaque colonie (morceau de bois en décomposition) a été déstructuré afin de vérifier la présence de reine(s) et du couvain, et a été conservé à condition de contenir au moins 500 ouvrières. Les nids avec leur matériel de construction ont été placés dans des boîtes en PVC de 1632 cm3 (, , ) disposées dans des bacs expérimentaux (, ) en laboratoire à température ambiante (Fig. 1). Une solution d'alcool à 70° et de talc a été étalée sur les bords des bacs pour éviter la fuite des fourmis. Les nids ont été régulièrement humectés à l'eau du robinet et les fourmis nourries avec du miel et de l'eau (environ tous les trois jours) durant au moins 20 jours avant le début des expériences, afin de permettre aux colonies de s'équilibrer avec leur nouveau milieu.
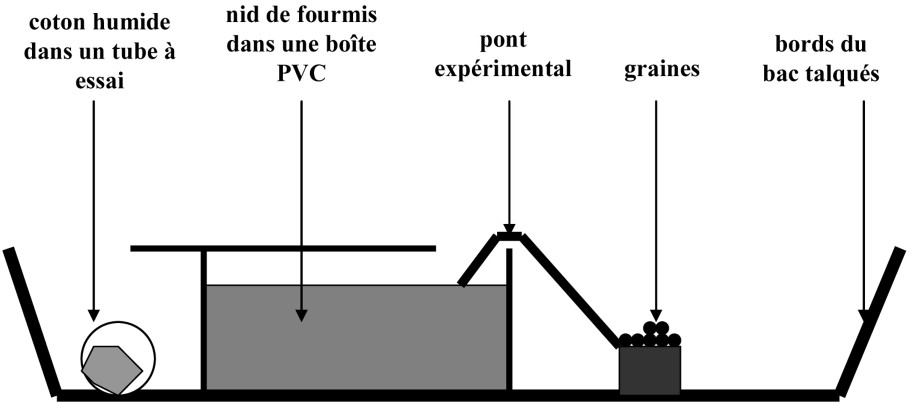
Système expérimental contenant la colonie de fourmis rouge des bois Myrmica ruginodis.
Le jour de chaque expérience, les graines à tester ont été décongelées 2 h avant le début des tests. Les colonies utilisées n'ont pas été nourries durant les 3 jours précédant les tests. Pour chaque espèce végétale, 10 graines ont été déposées sur un bouchon cylindrique en PVC incurvé au sommet (10 mm de hauteur et 22 mm de diamètre), dont les bords ont été talqués. Les bouchons ont été disposés dans les bacs et reliés à l'entrée du nid de fourmis par un pont en carton de 15 cm de long et 2,5 cm de large. Cinq répétitions des tests ont été réalisées par espèce végétale, soit un total de 50 graines par espèce, en utilisant à chaque fois des colonies de fourmis différentes. Les ponts et les bouchons ont été changés après chaque test. Le nombre de graines collectées par les fourmis dans chaque bouchon expérimental a été noté aux temps suivants à partir du premier contact graine-fourmi : 5 min, 15 min, 30 min, 60 min et 24 h. Durant l'expérience, la température ambiante a été contrôlée. Différentes observations sur le comportement de collecte et de transport des graines par les fourmis (transport ou non, morsure des graines, graines relâchées et reprises, consommation de l'élaiosome) ont été notées durant ces expériences.
2.4 Traits de vie et fréquences des espèces végétales sur le terrain
Afin d'examiner la relation entre les caractéristiques des graines et leur capacité à être dispersées par les fourmis, nous avons extrait de la base de données Biolflor [41] une série traits concernant la phénologie de floraison, et les caractéristiques des diaspores et des germinules. La diaspore est définie comme l'unité de dispersion et la germinule comme l'unité de germination sensu Klotz, 2002. La diaspore et la germinule peuvent correspondrent à la graine, mais cette dernière peut aussi être intégrée dans d'autres structures additionnelles, ou alors des structures additionnelles peuvent être attachées à la graine. Les traits des diaspores et des germinules sont relatifs à leur poids et à l'encombrement qu'elles pourraient générer lors de leur transport : épaisseur, largueur, longueur, rapport longueur/largueur, rapport longueur/épaisseur, rapport longueur/poids et poids.
Une étude de la flore précédemment réalisée sur 64 parcelles de la forêt de Compiègne [42], représentatives des conditions écologiques du massif, a été utilisée pour extraire les abondances des espèces végétales dans la strate herbacée et leurs fréquences relatives sur le terrain et les comparer à leur performance de transport par les fourmis. La forêt de Compiègne est la troisième plus grande forêt domaniale de plaine de France et regroupe un ensemble de conditions écologiques représentatives des forêts du Nord de la France, et plus largement du Nord-Ouest de l'Europe. Une description des conditions écologiques du site est détaillée dans des études locales récentes [28,42].
2.5 Analyses statistiques
Pour chaque espèce végétale, nous avons calculé les moyennes et les erreurs standard du pourcentage de graines récoltées aux différents temps de l'étude et du rythme de capture moyen (nombre de graines collectées par heure) basé sur la prise des graines durant les 30 premières minutes. Des profils moyens de collecte des graines par les fourmis ont été modélisés par des courbes de Lowess sous le logiciel ADE-4 [43]. Les comparaisons de collecte des graines entre espèces végétales ont été effectuées par des tests de Kruskal–Wallis (). Des analyses de corrélation de Spearman ont été utilisées pour mettre en relation le temps de capture des graines avec les traits de vie des espèces, leur recouvrement et leur fréquence sur le terrain.
3 Résultats
3.1 Performances de collecte des graines
La Fig. 2 ordonne les espèces végétales en fonction des rangs moyens des tests de Kruskal–Wallis ( ; ; ) et permet de distinguer trois groupes de plantes selon l'appétence de leurs graines pour les fourmis (les ouvrières étant la seule caste à participer à la collecte des graines). Un premier groupe d'espèces, de Colchicum autumnale à Melica uniflora, du haut en bas de la Fig. 2, possède des graines efficacement collectées par les fourmis (100% de graines collectées à 24 h pour la plupart des espèces) et inclut les graines à élaiosome très développé (Chelidonium majus, Luzula forsteri, Luzula multiflora, Viola reichenbachiana). Le deuxième groupe d'espèces montre un gradient très étalé de collecte des graines d'Euphorbia amygdaloides à Dactylis glomerata (Fig. 2), tandis que le troisième groupe comprend des espèces dont les graines intéressent très peu les fourmis (de Ranunculus repens à Rumex crispus). Le Tableau 1 donne les valeurs numériques des rythmes de capture des graines. Les graines de Chelidonium majus et de Colchicum autumnale sont les plus rapidement collectées au rythme de 100 graines par heure ou 1,67 graines par minute, c'est-à-dire que 1200 graines pourraient être potentiellement collectées dans les mêmes conditions en une demi-journée.
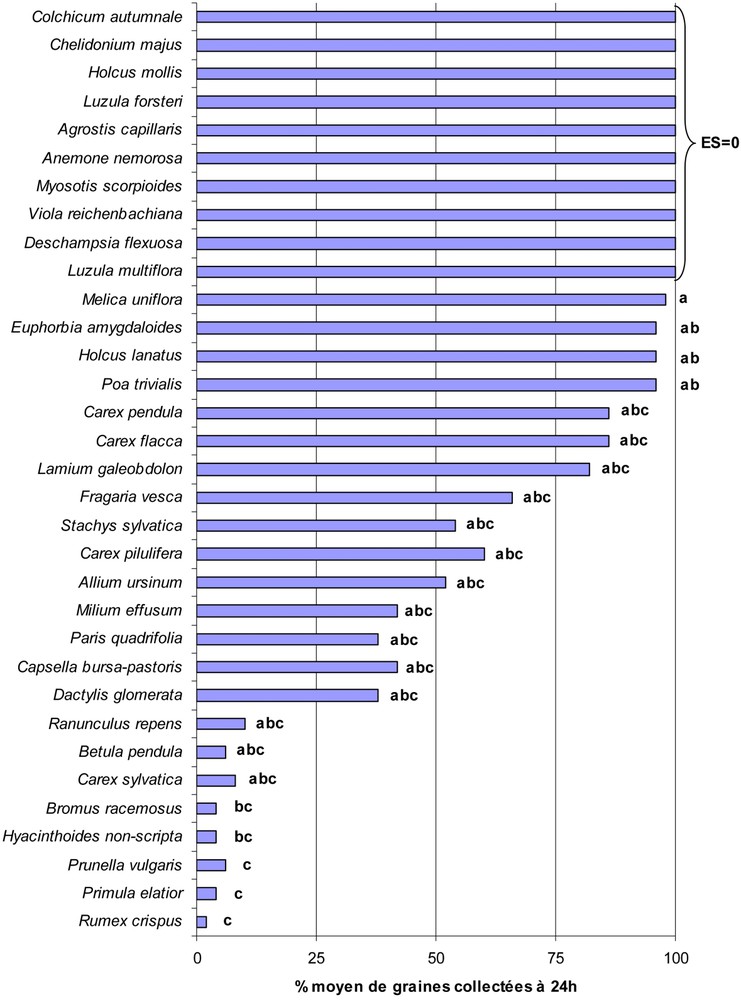
Comparaison du pourcentage de graines collectées par les fourmis après 24 heures d'expérience entre les espèces végétales. Les histogrammes indiquent les moyennes des pourcentages de graines collectées par espèce. Les espèces sont ordonnées selon les rangs moyens des tests de Kruskal–Wallis : DF=22 ; F=11,71 ; p<0,0001. Les espèces dont toutes les graines ont été collectées, c'est-à-dire présentant une variabilité inter-répétitions nulle à 24 h (erreur standard = 0) ont été exclues du test et présentées en haut du graphique.
Valeurs moyennes pour chaque espèce végétale du rythme de capture moyen des graines (nombre de graines collectées par heure) calculées sur la prise des graines à T+30 min (correspondant au palier des courbes de collecte pour la majorité des espèces, voir Fig. 3) et T+24 h pour les espèces dont aucune graine n'a été collectée à T+30 min
| Espèces végétales | Rythme de capture (n-graines/h) | ELAIO | MANIP | NETT | ABAND | REPR |
| Moy. ± 1 S.E. | ||||||
| Chelidonium majus | 100,00±0,00 | X | ||||
| Colchicum autumnale | 100,00±0,00 | X | ||||
| Holcus mollis | 98,00±2,00 | |||||
| Luzula forsteri | 94,00±4,00 | X | ||||
| Holcus lanatus | 88,00±7,35 | |||||
| Agrostis capillaris | 84,00±16,00 | |||||
| Melica uniflora | 76,00±17,49 | |||||
| Euphorbia amygdaloides | 74,00±9,27 | |||||
| Anemone nemorosa | 72,00±17,15 | X | ||||
| Lamium galeobdolon | 66,00±20,88 | |||||
| Deschampsia flexuosa | 66,00±20,88 | X | ||||
| Myosotis scorpioides | 66,00±16,00 | X | X | |||
| Viola reichenbachiana | 48,00±14,63 | |||||
| Poa trivialis | 44,00±14,35 | |||||
| Dactylis glomerata | 38,00±0,00 | X | ||||
| Carex pendula | 34,00±16,91 | |||||
| Luzula multiflora | 32,00±8,60 | |||||
| Fragaria vesca | 22,00±2,00 | X | ||||
| Stachys sylvatica | 18,00±9,17 | |||||
| Paris quadrifolia | 16,00±11,22 | X | X | |||
| Milium effusum | 14,00±9,80 | X | X | |||
| Carex flacca | 10,00±10,00 | |||||
| Capsella bursa-pastoris | 6,00±4,00 | |||||
| Allium ursinum | 4,00±2,45 | X | ||||
| Carex pilulifera | 4,00±2,45 | |||||
| Rumex crispus | 2,00±2,00 | X | ||||
| Carex sylvatica | 2,00±2,00 | |||||
| Betula pendula | 0,13±0,08 | X | ||||
| Prunella vulgaris | 0,13±0,13 | |||||
| Bromus racemosus | 0,08±0,05 | X | ||||
| Hyacinthoides non-scripta | 0,08±0,05 | X | ||||
| Primula elatior | 0,08±0,08 | |||||
| Ranunculus repens | 0,04±0,16 |
3.2 Profils des courbes de collecte des graines et comportements des fourmis
Les courbes de collecte des graines (Fig. 3) montrent que certaines espèces atteignent très rapidement le palier maximum, soit 100% de prélèvement par les fourmis. C'est le cas des espèces Colchicum autumnale et Chelidonium majus, dont 100% des graines ont été collectés en 30 min. Holcus mollis et Luzula forsteri atteignent ce palier au bout de 60 min. Pour la majorité des espèces, la zone d'inflexion de la courbe avant le palier se situe à 30 min. Ce temps a pour cela été utilisé pour calculer le rythme de capture des graines (Tableau 1). À cet instant, 50% des graines ont été collecté pour plus de 36% des espèces végétales testées. La Fig. 3, en comparaison des résultats de la Fig. 2, montre que pour les espèces Luzula multiflora, Poa trivialis, Carex pendula, Carex flacca, Carex pilulifera, Allium ursinum, Dactylis glomerata, le prélèvement des graines est tardif, et surtout enregistré à la fin de l'expérience.

Profils moyens de collecte des graines par les fourmis (valeurs cumulées) modélisés par des courbes de Lowess.
Les comportements des fourmis ont été notés au cours de l'expérience (Tableau 1). Les fourmis consomment les élaiosomes de certaines diaspores de Chelidonium majus et transportent ensuite les graines sans élaiosomes. Elles manipulent et nettoient la surface des graines à revêtement lisse et brillant (aspect huileux) ou contenues dans une baie (Paris quadrifolia). Des graines d'Anemone nemorosa, Betula pendula, Bromus racemosus, Fragaria vesca, Hyacinthoides non-scripta, Rumex crispus sont parfois relâchées durant le transport et définitivement abandonnées.
3.3 Corrélations entre le temps de capture des graines, leurs traits et la fréquence des plantes sur le terrain
Les analyses de corrélation de Spearman montrent qu'il n'existe pas de corrélation significative entre le temps de capture des graines et la phénologie de floraison (Tableau 2). Cependant, les pourcentages de graines collectées aux temps et sont négativement corrélés aux rapports longueur/largeur et longueur/épaisseur des diaspores et à la longueur et au rapport longueur/largeur des germinules. Aucune corrélation significative n'a non plus été trouvée entre le rythme de capture moyen des graines et le recouvrement ( ; ; ) ou la fréquence ( ; ; ) des plantes sur le terrain.
Coefficients de corrélations de Spearman (notés R) entre les traits phénologiques et morphologiques des diaspores et des germinules et le pourcentage de graines collectées aux temps T+15 min, T+30 min, T+24 h de l'expérience, et la vitesse de collecte des graines
| Traits des graines | Temps expérimental | Vitesse de collecte | |||
| 15 min | 30 min | 24 h | |||
| FLORAISON : | |||||
| Début | R | −0,163 | −0,149 | −0,026 | −0,078 |
| p | 0,364 | 0,407 | 0,887 | 0,668 | |
| n | 33 | 33 | 33 | 33 | |
| Fin | R | 0,023 | −0,010 | 0,091 | −0,044 |
| p | 0,899 | 0,957 | 0,614 | 0,808 | |
| n | 33 | 33 | 33 | 33 | |
| Durée | R | 0,320 | 0,278 | 0,324 | 0,160 |
| p | 0,069 | 0,118 | 0,066 | 0,374 | |
| n | 33 | 33 | 33 | 33 | |
| DIASPORES : | |||||
| Épaisseur | R | 0,169 | 0,201 | 0,117 | 0,214 |
| p | 0,421 | 0,335 | 0,577 | 0,305 | |
| n | 25 | 25 | 25 | 25 | |
| Largeur | R | −0,012 | 0,021 | −0,072 | 0,022 |
| p | 0,953 | 0,915 | 0,719 | 0,912 | |
| n | 27 | 27 | 27 | 27 | |
| Longueur | R | −0,282 | −0,216 | −0,198 | −0,187 |
| p | 0,154 | 0,278 | 0,322 | 0,349 | |
| n | 27 | 27 | 27 | 27 | |
| Longueur/largeur | R | −0,41 8(*) | −0,384(*) | −0,299 | −0,345 |
| p | 0,030 | 0,048 | 0,130 | 0,078 | |
| n | 27 | 27 | 27 | 27 | |
| Longueur/épaisseur | R | −0,431(*) | −0,386 | −0,279 | −0,369 |
| p | 0,032 | 0,056 | 0,177 | 0,070 | |
| n | 25 | 25 | 25 | 25 | |
| Longueur/poids | R | −0,317 | −0,334 | −0,229 | −0,292 |
| p | 0,131 | 0,111 | 0,282 | 0,166 | |
| n | 24 | 24 | 24 | 24 | |
| Poids | R | 0,100 | 0,130 | 0,071 | 0,109 |
| p | 0,619 | 0,519 | 0,723 | 0,587 | |
| n | 27 | 27 | 27 | 27 | |
| GERMINULES : | |||||
| Épaisseur | R | 0,039 | 0,079 | 0,064 | 0,088 |
| p | 0,836 | 0,678 | 0,738 | 0,645 | |
| n | 30 | 30 | 30 | 30 | |
| Largeur | R | −0,099 | −0,060 | −0,042 | −0,053 |
| p | 0,596 | 0,748 | 0,824 | 0,777 | |
| n | 31 | 31 | 31 | 31 | |
| Longueur | R | −0,376(*) | −0,310 | −0,173 | −0,262 |
| p | 0,034 | 0,085 | 0,344 | 0,147 | |
| n | 32 | 32 | 32 | 32 | |
| Longueur/largeur | R | −0,357(*) | −0,343 | −0,265 | −0,283 |
| p | 0,048 | 0,059 | 0,149 | 0,122 | |
| n | 31 | 31 | 31 | 31 | |
| Longueur/épaisseur | R | −0,345 | −0,317 | −0,238 | −0,284 |
| p | 0,062 | 0,088 | 0,206 | 0,129 | |
| n | 30 | 30 | 30 | 30 | |
| Longueur/poids | R | −0,267 | −0,285 | −0,195 | −0,252 |
| p | 0,170 | 0,141 | 0,321 | 0,196 | |
| n | 28 | 28 | 28 | 28 | |
| Poids | R | 0,083 | 0,120 | 0,076 | 0,110 |
| p | 0,670 | 0,534 | 0,694 | 0,569 | |
| n | 29 | 29 | 29 | 29 |
4 Discussion
4.1 Efficacité variable de l'élaiosome dans la dispersion des graines
Cette étude confirme que les graines à élaiosome sont efficacement dispersées par les fourmis, en particulier pour les espèces Chelidonium majus, Viola reichenbachiana, Luzula multiflora et Melica uniflora. Cependant, la présence d'un élaiosome chez d'autres espèces (Lamium galeobdolon, Carex pilulifera) connues pour leurs performances dans la myrmécochorie [34,44] ne leur garantit pas d'être rangées parmi celles les plus rapidement et efficacement transportées par les fourmis. Ce résultat est surtout surprenant pour Carex pilulifera, une espèce à statut myrmécochore bien confirmé, capable de nourrir des colonies de Myrmica ruginodis de façon exclusive après la période de fructification de la plante [14]. Une explication possible de ces résultats est la « fraîcheur » variable des élaiosomes. Lors de la récolte des graines de Lamium galeobdolon et Carex pilulifera, les élaiosomes avaient une apparence déjà desséchée sur les plantes. Les élaiosomes ne conserveraient donc pas leur « fraîcheur » de façon identique au cours du temps selon les espèces de plantes. En effet, leur pouvoir attractif est dû à la présence de composés chimiques [38] dont la teneur est souvent variable entre individus d'une même espèce [45,46] et pourrait décliner rapidement après la maturation et la chute des graines [14].
4.2 Reconsidération de la notion d'espèce myrmécochore
D'autres paramètres interviennent donc dans l'appétence des graines pour les fourmis et nous amènent à reconsidérer la notion d'espèce myrmécochore. Si certaines espèces sont incontestablement adaptées à la dispersion de leurs graines par les fourmis grâce à des tissus attractifs nettement visibles comme l'élaiosome, et sont qualifiées pour cela de myrmécochores, notre expérience montre qu'un grand nombre de graines sans adaptations spécifiques apparentes sont tout aussi efficacement transportées par les fourmis. C'est le cas de Poacées (Holcus mollis, Agrostis capillaris, Deschampsia flexuosa) dont les graines, petites et légères, sont faciles à transporter. De plus, le fruit des Poacées, le caryopse, contient un albumen amylacé abondant limité par une assise à gluten, une forme de réserve énergétiquement intéressante pour les fourmis. La stratégie de dispersion des graines des Poacées via l'alimentation des fourmis est opportuniste, c'est-à-dire que ce n'est pas un tissu spécialisé, mais la graine entière, qui fait office d'appât. Leur transport reste toutefois très efficace. L'inconvénient de cette stratégie opportuniste est qu'une large fraction des graines des Poacées est sacrifiée aux fourmis et consommée dans le nid (observations personnelles). Ce mécanisme de dispersion semble bénéficier davantage aux fourmis qu'aux plantes [21]. Ce n'est pas le cas des plantes mutualistes, dont les graines à élaiosomes sont avantageusement soustraites aux prédateurs [34] et attirent préférentiellement les fourmis mutualistes plutôt que granivores [38]. Finalement, il est difficile de tracer la frontière de la myrmécochorie au sein des espèces végétales. La notion d'espèce myrmécochore est surtout définie de manière binaire à partir de la présence/absence de tissus adaptés aux fourmis et ne tient absolument pas compte de la performance réelle du transport des graines. Les résultats de l'étude montrent cependant qu'il existe un gradient très étalé des performances de dispersion des espèces herbacées forestières par les fourmis, qui n'est pas uniquement dû à la présence de l'élaiosome.
Les paramètres morphologiques des graines sont rarement pris en compte dans la définition des espèces myrmécochores. Notre étude montre pourtant qu'ils ont une incidence sur la relation de mutualisme entre les plantes et les fourmis : plus le rapport longueur/largeur des graines est élevé, moins les graines sont rapidement transportées. Ce rapport révèle l'encombrement causé par la graine lors de son transport ; un rapport « optimum » de 1 correspondrait à une graine sphérique dont le centre de gravité se situerait entre les mandibules de l'insecte lors du transport (donc facile à transporter). L'irrégularité morphologique des graines pourrait donc gêner leur transport et protéger la graine de la granivorie des fourmis. Ces traits morphologiques pourraient également être le résultat d'un compromis (trade-off) entre, d'une part, le pouvoir attractif de la graine (épaisseur des réserves, petites graines faciles au transport), qui favorise la dispersion mais augmente le risque de consommation, et d'autre part, la difficulté de transport qui pousserait la fourmi à abandonner ou perdre la graine au cours de son trajet vers le nid, favorisant la survie des futures plantules. En effet, certaines graines ont été lâchées et abandonnées au cours de l'expérience. Des études confirment ce phénomène dans la nature, en montrant que des graines sont régulièrement perdues dans des anfractuosités du sol lors de leur transport, ceci contribuant à leur survie et au succès de la germination [47].
Le cas de la mélique (Melica uniflora) est très particulier, parce que c'est la seule Poacée qui possède un élaiosome développé et dont les graines sont de loin les plus lourdes (parmi celles des autres Poacées). Comme les résultats montrent que les fourmis moissonnent plus rapidement les Poacées à petites graines, le succès de la mélique apparaît contradictoire à l'examen de ses propres traits morphologiques et pondéraux. Pour cette espèce, le développement de l'élaiosome au cours de l'évolution de la plante aurait pu permettre l'augmentation du poids des graines, donc des réserves, donc de la chance de survie des plantules, sans porter préjudice au caractère attractif de la graine, c'est-à-dire à sa fitness, au sens darwinien du terme. L'élaiosome aurait compensé le désavantage de la prise de poids de la graine pour son transport et maintenu le pouvoir sélectif de la plante.
4.3 Rôle des fourmis et succès des plantes dans leur milieu de vie
Bien que l'effet des fourmis sur la répartition des plantes soit bien connu [3,22], nos résultats ne montrent pas de corrélation significative entre l'efficacité expérimentale du transport des graines et la fréquence ou l'abondance des plantes sur le terrain à l'échelle du massif forestier. Même si les fourmis jouent un rôle important dans la dispersion et le maintien des espèces en enterrant leurs graines dans des microsites favorables, elles ne garantissent pas la survie des plantes face au filtre environnemental forestier constitué d'un ensemble complexe de conditions écologiques très contraignantes, comprenant notamment les propriétés du sol, la luminosité et les perturbations anthropiques [48,49]. Bien que les fourmis compensent parfois l'effet destructeur de ces perturbations sur les populations végétales forestières [2], nos analyses, confrontant les résultats expérimentaux à la réalité du terrain, sont au contraire en accord avec d'autres études montrant que d'autres facteurs limitent la migration des plantes forestières, malgré la fonction de disperseur de graines assurée par les fourmis [50]. De plus, les facteurs locaux, édaphiques ou topographiques, peuvent aussi influencer indirectement, mais fortement, la dispersion des graines des plantes myrmécochores en réduisant la densité locale des fourmis [51], et donc expliquer la distorsion entre les résultats expérimentaux et la répartition réelle des plantes sur le terrain. Une étude à échelle plus fine sur quelques mètres carrés autour des nids de fourmis devrait permettre de mettre en évidence des patrons de dispersion locaux, impossibles à observer à notre échelle d'étude, c'est-à-dire celle du massif forestier. Enfin, les relations plantes–fourmis sont presque toujours asymétriques et les bénéfices « attendus » autant par les plantes que par les fourmis ne sont pas toujours réalisés. D'une part, la myrmécochorie est le plus souvent décrite comme le résultat de l'évolution des plantes « trompant » les fourmis [21] ; d'autre part, la dispersion des graminées semble plus avantageuse au jabot social des fourmis qu'à la survie des graines. Ces mécanismes très complexes et particuliers à chaque espèce masquent probablement, ou empêchent, la formation d'une relation linéaire entre l'efficacité de la dispersion et le succès des plantes in situ.
L'examen de l'écologie des espèces végétales indique que les deux espèces les plus efficacement dispersées par les fourmis sont des espèces non strictement forestières : Colchicum autumnale poussant principalement sur les talus des chemins et Chelidonium majus sur les zones d'éboulis et sur les murs des habitations. Cela suggère que Myrmica ruginodis pourrait occuper un rôle important dans l'équilibre lisière–forêt, notamment en permettant la pénétration des espèces de lisière à l'intérieur des parcelles forestières où se trouvent les nids de fourmis. Ainsi, elles accélèreraient la colonisation des trouées naturelles ou créées par l'homme, isolées dans les parcelles forestières par les plantes herbacées des lisières. Toutefois, les espèces non forestières (Rumex crispus, Bromus racemosus, Prunella vulgaris, Ranunculus repens, Dactylis glomerata, Capsella bursa-pastoris) sont en majorité très peu transportées, confirmant la spécialisation de Myrmica ruginodis aux écosystèmes forestiers.
5 Conclusions
Cette étude montre que deux stratégies de dispersion des graines se distinguent et font appel, soit au comportement de mutualisme, soit au comportement granivore des fourmis. Les espèces dites myrmécochores possèdent des tissus adaptés à un comportement mutualiste des fourmis et sont très rapidement collectées. Les Poacées n'ont pas d'organe spécialisé, mais montrent un gradient d'attractivité qui est le résultat d'un compromis entre le bénéfice du transport par les fourmis et le risque d'être consommé par des fourmis occasionnellement granivores. Au regard de la diversité et de la quantité des graines potentiellement transportées par les fourmis, il apparaît important de préserver les colonies de Myrmica ruginodis en maintenant une bonne qualité de leur lieu de nidification et de leur habitat en général, c'est-à-dire en conservant un minimum de bois mort sur le sol des parcelles forestières.
Remerciements
Les auteurs tiennent particulièrement à remercier M. Alain Lenoir pour son aide dans la détermination des espèces de fourmis et pour ses commentaires sur le manuscrit.