

1 Introduction
Among extant snakes (∼3150 species), Caenophidia or “advanced” snakes are a monophyletic group comprising the great majority (∼2620 species) of species [1–4]. The American caenophidian snake fauna comprises five families: the Viperidae and Elapidae – both displaying a front-fanged venom system – and the colubroidean families Natricidae, Colubridae and Dipsadidae [2,5]. The latter is one of the largest families of snakes (> 700 species), with all living representatives restricted to the New World [6,7]. Their closest relatives are the Pseudoxenodontidae [2,3,5] and the genus Thermophis [8,9], both from Asia. Dipsadids probably have an Asian-North American origin, as do all the other American caenophidians [7].
Dipsadids are primarily tropical, with most occurring in Central America, South America and the West Indies. They vary greatly in body size (up to 280 cm adult total length [10]) and in ecology. Most species feed on frogs and lizards, but some specialize on snakes or slugs, snails, and earthworms.
Following Cadle's [11–14] pioneering work based on immunological data, three DNA studies have focused on higher level dipsadid relationships: Vidal et al. [7], Pinou et al. [15], and Zaher et al. [16], and several studies have focused on subgroups of dipsadids: Feldman and Spicer [17] (Contia), Feldman and Spicer [18] (Contia and Diadophis), Mulcahy [19] (tribe Leptodeirini), Fontanella et al. [20] (Diadophis), Mulcahy [21] (Hypsiglena), Daza et al. [22] (tribe Leptodeirini, Leptodeira), Mulcahy and Macey [23] (Hypsiglena), and Hedges et al. [24] (tribe Alsophiini).
Higher level studies generally agree on the delineation of three major clades of Dipsadidae broadly consistent in their distribution: a South American clade including West Indies (Xenodontinae), a Central American clade (Dipsadinae) and a North American clade (Heterodontinae Bonaparte, 1845; having priority to Heterodontinae Gray, 1851).
Although heterodontines include only five genera (Carphophis, Contia, Diadophis, Farancia and Heterodon) and nine species (but several “cryptic” species to be described), they have been the subject of very detailed studies [17,18,20]. This is not the case for the two species-rich Neotropical dipsadid subfamilies: the dipsadines and xenodontines.
The dipsadines include 25 genera (∼350 species): Adelphicos, Amastridium, Atractus, Chapinophis, Chersodromus, Coniophanes, Cryophis, Dipsas, Geophis, Hypsiglena, Imantodes, Leptodeira, Ninia, Plesiodipsas, Pliocercus, Pseudoleptodeira, Rhadinaea, Rhadinophanes, Sibon, Sibynomorphus, Tantalophis, Tretanorhinus, Trimetopon, Tropidodipsas, Urotheca [1,4,16,19,21,22,25,26]. Among dipsadines, Eridiphas was synonymized with Hypsiglena [21], and Daza et al. [22] found Tantalophis to belong to the subfamily. Resolutions of several intergeneric relationships have been proposed recently: the genera Imantodes and Leptodeira are sister-groups (tribe Leptodeirini) as are Pseudoleptodeira and Hypsiglena [19,22]. Moreover, Cryophis, Atractus, Sibon, Ninia, Sibynomorphus and Dipsas form a clade (tribe Dipsadini), as do Amastrium, Coniophanes and Rhadinae [16,19,22].
The xenodontines include 49 genera (∼330 species): Alsophis, Apostolepis, Arrhyton, Boiruna, Borikenophis, Caaeteboia, Calamodontophis, Caraiba, Clelia, Conophis, Cubophis, Ditaxodon, Drepanoides, Echinantera, Elapomorphus, Erythrolamprus, Gomesophis, Haitiophis, Helicops, Hydrodynastes, Hydrops, Hypsirhynchus, Ialtris, Liophis, Lygophis, Magliophis, Manolepis, Mussurana, Oxyrhopus, Phalotris, Philodryas, Phimophis, Pseudalsophis, Pseudoboa, Pseudoeryx, Pseudotomodon, Psomophis, Ptychophis, Rhachidelus, Saphenophis, Siphlophis, Tachymenis, Taeniophallus, Thamnodynastes, Tomodon, Tropidodryas, Umbrivaga, Uromacer, Xenodon [1,4,16,24,26–27]. Three studies have generated 12S and 16S rRNA sequences for multiple xenodontines: Vidal et al. [7], Hedges et al. [24] and Zaher et al. [16]. The relationships, classification and biogeography of West Indian xenodontines (Tribe Alsophiini, genera Alsophis, Arrhyton, Borikenophis, Caraiba, Cubophis, Haitiophis, Hypsirhynchus, Ialtris, Magliophis and Uromacer) have been covered extensively by Hedges et al. [24], and are not dealt with further here. Among mainland xenodontines, the monophyly of the Hydropsini (genera Helicops, Hydrops and Pseudoeryx), the Xenodontini, and the Pseudoboini has been recovered both by Vidal et al. [7] and Zaher et al. [16]. Among Xenodontini, Zaher et al. [16] proposed several nomenclatural changes, as they resurrected Lygophis, synonymized Lystrophis and Waglerophis with Xenodon, and synonymized Erythrolamprus with Liophis. The latter of these actions is incorrect because Erythrolamprus Boie, 1826, has priority over Liophis Wagler, 1830 [27]. The Xenodontini therefore provisionally includes the genera Liophis, Erythrolamprus, Lygophis, Xenodon and Umbrivaga; the latter genus has never been sampled in molecular studies.
Within Pseudoboini, two genera were recently described, Boiruna [28] and Mussurana [16], and the tribe currently includes the following genera: Boiruna, Clelia, Drepanoides, Mussurana, Oxyrhopus, Phimophis, Pseudoboa, Siphlophis, and Rhachidelus; the latter genus has never been sampled in molecular studies. Other monophyletic groups of genera are the Elapomorphini (Apostolepis, Elapomorphus, and Phalotris) [16], the Tachymenini (Calamodontophis, Gomesophis, Pseudotomodon, Ptychophis, Tachymenis, Thamnodynastes and Tomodon) [16] and the Philodryadini [7,16]. Among Philodryadini, Zaher et al. [16] synonymized Pseudablabes and Xenoxybelis with Philodryas, so that the tribe now includes Philodryas and Ditaxodon; the latter genus has never been sampled in molecular studies. The affinities within xenodontines of several previously sampled genera remain less clear, so that we are hesitant to assign them to separate tribes pending further studies (contra Zaher et al. [16]). These are Caaeteboia (previously Liophis amarali), Pseudalsophis (previously Alsophis elegans), Psomophis, Tropidodryas, Taeniophallus, Conophis, and Hydrodynastes [7,16]. Three other genera are considered xenodontines [1,16], but have never been sampled in molecular studies: Echinantera, Manolepis, and Saphenophis.
Finally, 13 enigmatic Neotropical genera representing 23 species are considered to be Dipsadidae incertae sedis [1,16]: Cercophis, Crisantophis, Diaphorolepis, Emmochliophis, Enuliophis, Enulius, Hydromorphus, Lioheterophis, Nothopsis, Sordellina, Synophis, Uromacerina and Xenopholis.
In this study, we combined the 12S and 16S sequences previously obtained by Vidal et al. [7] and Zaher et al. [16] and produced new sequences from 18 taxa including seven genera not previously sampled: Crisantophis, Echinantera, Manolepis, Nothopsis, Trimetopon, Umbrivaga and Xenopholis.
2 Materials and methods
DNA extraction was using Winnepenninckx et al.’s [29] protocol or the DNeasy Tissue Kit from Qiagen. Samples used for this work, with corresponding localities and voucher details are listed in the Appendix.
Amplification was achieved using the following sets of primers: L2510, 5’-CGC-CTG-TTT-ATC-AAA-AAC-AT-3’ [30]; L16, 5’-ACG-GCC-GCG-GTA-YCC-TAA-CCG-TG-3’ [7] and H3056, 5’-CTC-CGG-TCT-GAA-CTC-AGA-TCA-CGT-AGG-3’ [31] for the 16SrRNA gene; L12, 5’-CGC-CAA-AYA-ACT-ACG-AG-3’ [7]; H1478, 5’-TGA-CTG-CAG-AGG-GTG-ACG-GGC-GGT-GTG-T-3’ [32] and H1557, 5’-GTA-CAC-TTA-CCT-TGT-TAC-GAC-TT-3’ [33] for the 12SrRNA gene. Both strands of PCR products were sequenced manually or using the CEQ cycle sequencing kit (Beckman) in the CEQ-2000 DNA Analysis System (Beckman), or the BigDye sequencing kit (Applied Biosystems) in the ABI Prism 3100-Avant Genetic Analyser.
The two strands obtained for each sequence were combined using BioEdit [34]. The sequences generated for this work are deposited in GenBank under accession numbers GU018143-GU018178. Sequence entry and alignment were performed with MUST2000 [35] and MEGA4 [36]. For 12S, alignment was guided by a secondary structure model [37] and it built on a previous alignment [7] that has been appraised independently [38]. In our 16S alignment, one obvious and highly variable loop region was removed. Following this, in both genes, only unambiguous gaps of at most three base pairs length were retained. In all further analyses, gaps were treated as missing data. Alignments can be obtained from Nicolas Vidal. Alignments resulted in 303 12S rRNA sites (173 variable sites, 148 of which are parsimony informative) and 374 16S rRNA sites (168 variable sites, 132 of which are parsimony informative) for 130 taxa.
We built phylogenies from the combined 12S and 16S rRNA fragments using Maximum Likelihood (ML) and Bayesian methods of inference. Based on previously obtained results [2], we used an elapid (Micrurus surinamensis), two natricids (Natrix natrix and Rhabdophis subminiatus), and two colubrids (Grayia ornata and Rhinobothryum lentiginosum) as outgroups. ML analyses were performed with RAxML 7.0.4 [39–40], and Bayesian analyses were performed with Bayes 3.1 [41]. For both analyses, we used a GTR model as inferred by Modeltest using the AIC criterion [42]. Bayesian analyses were performed by running 5,000,000 generations in four chains, saving the current tree every 100 generations. The last 48,000 trees were used to construct a 50% majority rule consensus tree. For the ML analysis, we performed 1000 bootstrap replicates.
3 Results and discussion
Dipsadids are monophyletic with the North American genera (Heterodontinae: Farancia; Carphophis + Contia + Diadophis; Heterodon) lying outside a clade comprising all other sampled dipsadids, consistent with the results of Vidal et al. [7] and Zaher et al. [16]. The phylogenetic status of the Heterodontinae remains uncertain because of low support for the basalmost nodes in the tree (Fig. 1a).
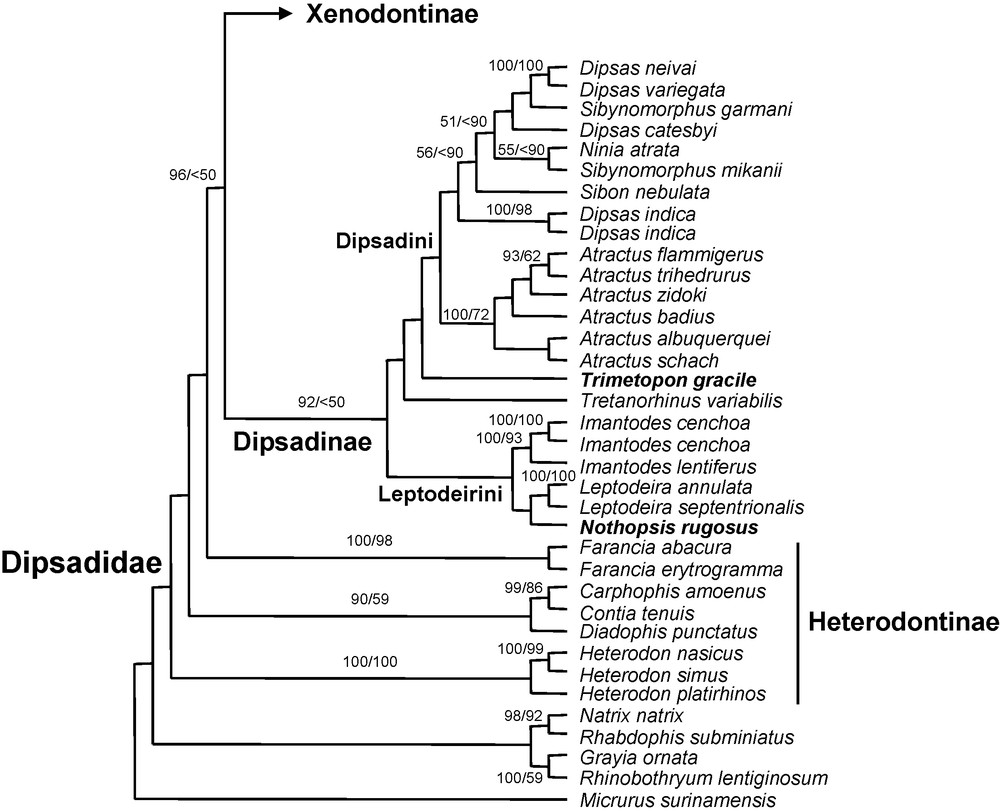
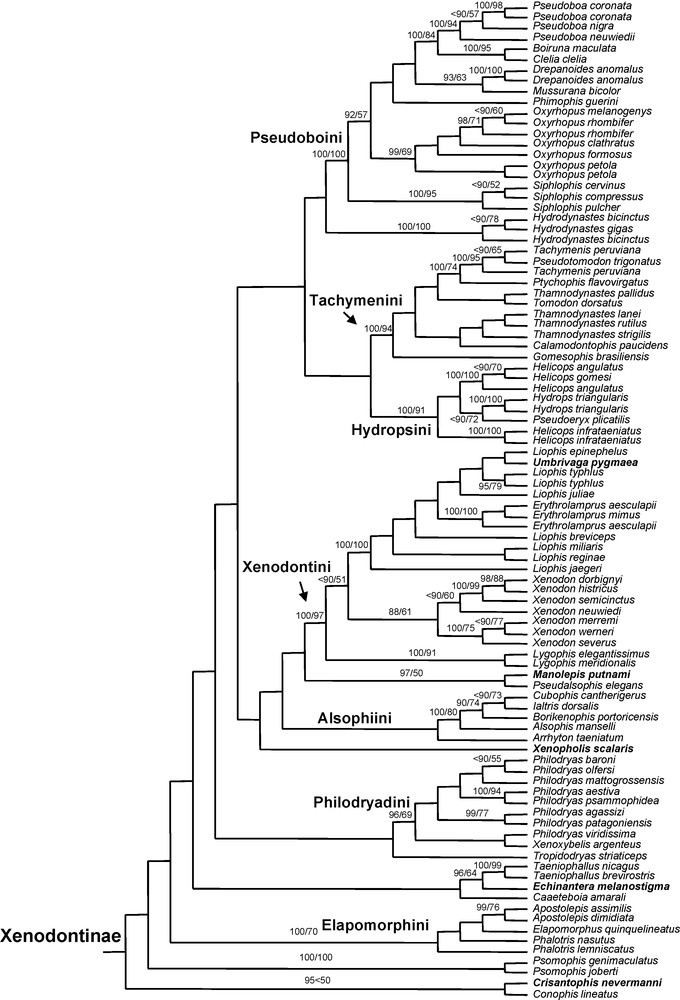
Bayesian majority rule consensus tree (12S and 16S rRNA, 130 taxa, 677 sites). Nodes with values are supported by Bayesian posterior probabilities above 90% (first value) and ML bootstrap values above 50% (second value). Never before sampled genera are in bold.
The dipsadines are monophyletic (PP value: 92%, ML BP value < 50%) and include the Central American genus Trimetopon. The monotypic Nothopsis is the sister-group to Leptodeira, those two genera forming a clade with Imantodes that is sister to all other sampled dipsadines. We therefore add Nothopsis to the Tribe Leptodeirini. The other half of the basal split among dipsadines comprises Tretanorhinus, Trimetopon, Atractus, Dipsas, Sibon, Sibynomorphus and Ninia. The monophyly of the tribe Dipsadini, including at least Atractus, Dipsas, Sibon, Sibynomorphus, Ninia and Cryophis (not sampled here) is therefore confirmed. The genera Dipsas and Sibynomorphus are each paraphyletic, but with moderate support, and more data will be needed to resolve the issue.
Among xenodontines, the monophyly of the following six previously recognized tribes is retrieved with good to strong support (PP values from 96 to 100%, BP values from 69 to 100%): Elapomorphini, Hydropsini, Philodryadini, Pseudoboini, Tachymenini and Xenodontini. The monophyly of the Alsophiini is retrieved with moderate support (PP 86%, BP 47%).
The tribe Elapomorphini includes Apostolepis, Elapomorphus, and Phalotris, the paraphyly of the latter is only weakly supported.
The Hydropsini includes Helicops, Hydrops, and Pseudoeryx, the latter two genera being sister groups. The genus Helicops is recovered as paraphyletic, but with weak support, and we consider the genus likely to be monophyletic given the available evidence [16]. The sequences deposited by Zaher et al. [16] under the names Helicops pictiventris and H. infrataeniatus are virtually identical (p-distance: 0.3%) and are here treated as deriving both from Helicops infrataeniatus given that H. pictiventris was synonymized with this species by Lema et al. [43].
Among Philodryadini, Philodryas appears to be paraphyletic with respect to Xenoxybelis, but with low support, and we recommend the recognition of the morphologically distinctive genus Xenoxybelis pending further studies with denser taxonomic and character sampling.
The tribe Pseudoboini includes Boiruna, Clelia, Drepanoides, Mussurana, Oxyrhopus, Phimophis, Pseudoboa and Siphlophis. Within Oxyrhopus, O. rhombifer (sampled from Brazil and Paraguay) appears paraphyletic with respect to Oxyrhopus melanogenys (sampled from French Guiana), but the two samples of O. petola (Costa Rica and Ecuador) do form a clade.
The tribe Tachymenini includes Calamodontophis, Gomesophis, Pseudotomodon, Ptychophis, Tachymenis, Thamnodynastes and Tomodon, as found by Zaher et al. [16]. The genus Thamnodynastes is recovered as polyphyletic, but with low support.
The tribe Xenodontini (represented here by 21 species) includes Lygophis, Xenodon, Liophis, Erythrolamprus and Umbrivaga. If Lygophis and Xenodon are each monophyletic, the genus Liophis is paraphyletic with respect to Erythrolamprus and Umbrivaga. Umbrivaga pygmaea is not the type species of Umbrivaga, but the genus includes only three species and based on traditional data [44] we expect U. mertensi (the type species) and U. pyburni to cluster with U. pygmaea when molecular data for these species are available [44]. Erythrolamprus is confirmed as monophyletic. Erythrolamprus mimus is probably the species most likely to have challenged the monophyly of the genus given its peculiar tooth morphology [45–48], but this has not happened and we fully expect the genus to remain monophyletic when other taxa are sampled, including the type species (Coluber venustissimus Wied-Neuwied, 1821; presently E. aesculapii venustissimus). Regarding Liophis, the type species is L. cobella, and according to Fernandes et al. [49], the species L. breviceps (here sampled) belongs to the L. cobella group. In view of the available evidence, Liophis is clearly paraphyletic and so some genus-level changes to the taxonomy of Xenodontini will be required to maintain taxon monophyly. The two most obvious solutions would seem to be to synonymize both Liophis and Umbrivaga with Erythrolamprus, or to synonymize some Liophis with Umbrivaga and delay other possible changes among other Liophis until more analyses are conducted with greater taxon and character sampling. Whichever way is chosen, workers should be aware of the inadequacy of the current taxonomy.
The following genera cannot be allocated to the above tribes with any reasonable confidence based on our results: Caaeteboia, Echinantera and Taeniophallus, Tropidodryas, Manolepis and Pseudalsophis, Xenopholis, Psomophis, Hydrodynastes, Conophis and Crisantophis. Zaher et al. [16] assigned several of these genera to particular tribes, but we consider this premature and suggest that additional studies including greater taxon and character sampling are required. For example, Zaher et al. [16] created the tribe Saphenophiini to include Pseudalsophis and Saphenophis (the latter genus has never been sampled in molecular studies) and the tribe Conophiini to include Conophis and Manolepis (the latter genus was not sampled in Zaher et al. [16]). Our analyses, based on an expanded taxon sampling, recover Manolepis clustering with Pseudalsophis (PP 97%, BP 50%), and Conophis with Crisantophis (PP 95%, BP < 50%), thus we recover the tribe Conophiini as polyphyletic and the tribe Saphenophiini as paraphyletic. Moreover, if a new tribe was to be erected to allocate the genera Pseudalsophis Zaher et al., 2009, Saphenophis Myers, 1973 and Manolepis Cope, 1885, priority of the latter could be considered for naming the new taxon.
Acknowledgements
This work was funded by the Service de Systématique Moléculaire du Muséum National d’Histoire Naturelle. We thank all those persons who contributed tissue samples used in this study: Michel Blanc, Jonathan A. Campbell, Herndon Dowling, S. Blair Hedges, Andreas Hohmeister, Christian Marty, Linda Maxson, Mahmood Sasa, Jean-Pierre Vacher, and Wolfgang Wüster. DJG thanks Philippe Gaucher, Mark Wilkinson and Camp Patawa for help with fieldwork.
Appendix
List of taxa used for this work. Taxa in bold have been sequenced for this study. Abbreviations: JAC: Jonathan A. Campbell; SBH: S. Blair Hedges; AH: Andreas Hohmeister; MS: Mahmood Sasa (Grant for Vicerrectoria de Investigacion, Universidad de Costa Rica, Project 741-A1-021); JPV: Jean-Pierre Vacher; NV: Nicolas Vidal; WW: Wolfgang Wüster. FG: French Guiana; WI: West Indies; MW: field tag for specimen to be accessioned into the Natural History Museum, London.
Alsophis manselli (WI, AF158459, AF158528), Apostolepis assimilis (GQ457781, GQ457724), Apostolepis dimidiata (GQ457782, GQ457725), Arrhyton taeniatum (WI, AF158453, AF158522), Atractus albuquerquei (GQ457783, GQ457726), Atractus badius (FG, AF158425, AF158485), Atractus flammigerus (FG, AF158402, AF158471), Atractus schach (FG, AF158427, AF158486), Atractus trihedrurus (GQ457784, GQ457727), Atractus zidoki (FG, AF158426, AF158487), Boiruna maculata (GQ457785), Borikenophis portoricensis (WI, AF158448, AF158517), Caaeteboia amarali (GQ457807, GQ457747), Calamodontophis paucidens (GQ457786, GQ457728), Carphophis amoenus (AY577013, AY577022), Clelia bicolor (GQ457787, GQ457729), Clelia clelia (FG, AF158403, AF158472), Conophis lineatus (JAC, UTA R 46849, Aldea “El Arenal”, San Vincente, Zacapa, Guatemala), Contia tenuis (AY577021, AY577030), Crisantophis nevermanni (MS, Palo Verde National Park, Bagaces, Guanacaste, Costa Rica), Cubophis cantherigerus (WI, AF158405, AF158475), Diadophis punctatus (AF544765, AF544793), Dipsas catesbyi (Z46459, Z46496), Dipsas indica (FG, AF158421, AF158488), Dipsas indica (GQ457789, GQ457730), Dipsas neivai (GQ457790, GQ457731), Dipsas variegata (FG, AF158406, AF158476), Drepanoides anomalus (FG, AF158407, AF158477), Drepanoides anomalus (GQ457791, GQ457732), Echinantera melanostigma (SBH268510, unknown origin), Erythrolamprus aesculapii (FG, AF158462, AF158531), Erythrolamprus aesculapii (GQ457795, GQ457736), Elapomorphus quinquelineatus (GQ457794, GQ457735), Erythrolamprus mimus (MS, ICP 1105, Guapiles, Limon, Costa Rica), Farancia abacura (Z46491, Z46467), Farancia erytrogramma (AY577017, AY577026), Gomesophis brasiliensis (GQ457796, GQ457737), Grayia ornata (AF158434, AF158503), Helicops angulatus (FG, AF158408, AF158478), Helicops angulatus (GQ457797, GQ457738), Helicops gomesi (GQ457798, GQ457739), Helicops infrataeniatus (GQ457799, GQ457740), Helicops infrateniatus (GQ457800, GQ457741), Heterodon nasicus (GQ457801, AF158494), Heterodon platirhinos (AY577019, AY577028), Heterodon simus (AY577020, AY577029), Hydrodynastes bicinctus (FG, AF158430, AF158479), Hydrodynastes bicinctus (GQ457802, GQ457742), Hydrodynastes gigas (GQ457803, GQ457743), Hydrops triangularis (FG, AF158415, AF158499), Hydrops triangularis (GQ457804, GQ457744), Ialtris dorsalis (WI, AF158456, AF158525), Imantodes cenchoa (FG, AF158429, AF158495), Imantodes cenchoa (GQ457805, GQ457745), Imantodes lentiferus (FG, AF158463, AF158473), Leptodeira annulata (FG, AF158404, AF158473), Leptodeira septentrionalis (WW, Old Northern Highway, 14.3 km N. Sand Hill, Orange Walk, Belize), Liophis breviceps (FG, AF158464, AF158533), Liophis epinephelus (MS, Barrio Lujan, San José, Costa Rica), Liophis jaegeri (GQ457809, GQ457749), Liophis juliae (WI, AF158445, AF158514), Liophis miliaris (FG, AF158409, AF158480), Liophis reginae (FG, AF158433, AF158501), Liophis typhlus (FG, AF158410, AF158481), Liophis typhlus (GQ457811, GQ457751), Lygophis elegantissimus (GQ457808, GQ457748), Lygophis meridionalis (GQ457810, GQ457750), Manolepis putnami (JAC, Mexico, Guerrero, Ejido de Bahia, 16.95759N/99.94158W, JAC 22044), Micrurus surinamensis (FG, AF544770, AF544799), Natrix natrix (AF158461, AF158530), Ninia atrata (GQ457814), Nothopsis rugosus (MS, ASL 493, Guayacan, Siquirres, Limon, Costa Rica), Oxyrhopus clathratus (GQ457815, GQ457754), Oxyrhopus formosus (FG, AF158411, AF158482), Oxyrhopus melanogenys (FG, AF158422, AF158489), Oxyrhopus petola (AH, Costa Rica), Oxyrhopus petola (WW, Zamora, Chinchipe Province, Ecuador), Oxyrhopus rhombifer (NV, Paraguay), Oxyrhopus rhombifer (GQ457816, GQ457755), Phalotris lemniscatus (GQ457817, GQ457756), Phalotris nasutus (GQ457818, GQ457757), Philodryas aestiva (GQ457819, GQ457758), Philodryas agassizi (GQ457823, GQ457762), Philodryas baroni (Argentina, AF158469, AF158534), Philodryas mattogrossensis (GQ457820, GQ457759), Philodryas olfersi (Brazil, AF158417, AF158484), Philodryas patagoniensis (GQ457821, GQ457760), Philodryas psammophidea (AH, Paraguay), Philodryas viridissima (FG, AF158419, AF158474), Phimophis guerini (GQ457822, GQ457761), Pseudalsophis elegans (Chili, AF158401, AF158470), Pseudoboa coronata (FG, AF158412, AF158483), Pseudoboa coronata (GQ457824, GQ457763), Pseudoboa neuwiedii (FG, AF158423, AF158490), Pseudoboa nigra (GQ457825, GQ457764), Pseudoeryx plicatilis (FG, AF158418, AF158500), Pseudotomodon trigonatus (GQ457827, GQ457766), Psomophis genimaculatus (GQ457828, GQ457767), Psomophis joberti (GQ457829, GQ457768), Ptychophis flavovirgatus (GQ457830, GQ457769), Rhabdophis subminiatus (AF544776, AF544805), Rhinobothryum lentiginosum (FG, AF158465, AF158535), Sibon nebulata (FG, AF544777, AF544806), Sibynomorphus garmani (GQ457831, GQ457770), Sibynomorphus mikanii (GQ457832, GQ457771), Siphlophis cervinus (FG, AF158466, AF158536), Siphlophis compressus (FG, AF158467, AF158537), Siphlophis pulcher (GQ457834, GQ457773), Tachymenis peruviana (JAC, Peru, Cusco, 1 km N. of Kusilluchayoc, UTA R 51461), Tachymenis peruviana (GQ457835, GQ457774), Taeniophallus nicagus (FG, AF158414, AF158502), Taeniophallus brevirostris (GQ457793, GQ457734), Thamnodynastes lanei (GQ457836, GQ457775), Thamnodynastes pallidus (FG, AF158420, AF158492), Thamnodynastes rutilus (GQ457837, GQ457776), Thamnodynastes strigilis (SBH 268509, Argentina), Tomodon dorsatus (GQ457838, GQ457777), Tretanorhinus variabilis (WI, AF158460, AF158529), Trimetopon gracile (MS, RV, Las Nubes de Coronado, San José, Costa Rica), Tropidodryas striaticeps (GQ457838, GQ457778), Umbrivaga pygmaea (MW 5778, Patawa, French Guiana), Xenodon dorbignyi (GQ457812, GQ457752), Xenodon histricus (GQ457813, GQ457753), Xenodon merremii (GQ457840), Xenodon neuwiedii (GQ457841, GQ457779), Xenodon semicinctus (NV, Paraguay), Xenodon severus (Z46474, Z46449), Xenodon werneri (FG, AF158468, AF158538), Xenopholis scalaris (JPV, French Guiana), Xenoxybelis argenteus (FG, AF158413, AF158493).