

Abridged English version
1 Introduction
The understanding of the presently increasing greenhouse effects, and more generally, of climatic change effects on ecosystems requires a better understanding of physical, biochemical, and biological processes occurring in the oceans. Oceans contribute to global climate, they buffer atmospheric greenhouse gas, and they sustain some of the most productive ecosystems of the planet.
However, the functioning of food web and ecosystem functioning is still poorly understood, particularly at the top trophic level, where both climate and human activities act. The effects of climate changes on marine ecosystems can be investigated either directly through marine resources changes, or indirectly through changes occurring in top predator populations. For instance, some natural cyclic phenomena occur at a relatively short scale in time and space, compared to global change, but they have consequences of similar scales compared to those obtained by simulation of climate change.
2 Climatic anomalies and natural observatories
There is increasing literature on large-scale consequences of climatic changes (mainly El Niño Southern Oscillation, North Atlantic Oscillation, and Antarctic Circumpolar Wave) on ecosystems functioning.
El Niño is associated with large-scale changes in the atmosphere–ocean system in the Pacific Ocean, and its intensity is expressed using El Niño Southern Oscillation ENSO indices. El Niño is associated with negative atmospheric anomalies and the displacement of warm sea surface water on the western coasts of South America, where sea surface temperature anomalies can be up to 3–4 °C. This reduces and sometimes stops the coastal upwelling of South America. La Niña is the reverse phenomenon with abnormal cooling of sea surface waters. El Niño and La Niña alternate on a 6–8-year basis.
The North Atlantic Oscillation is similar to El Niño, but it occurs in the Atlantic Ocean. Positive anomalies are associated with strong westerlies shifted north and increase rains and temperatures in northern Europe while inducing dry anomalies in the Mediterranean Sea. Negative anomalies induce reverse situations. Positive and negative anomalies alternate on a decadal basis.
The Antarctic Circumpolar Wave is a complex system of coupled anomalies (two positive anomalies alternating with two negative anomalies) circulating easterly around the Southern Ocean within 8–10 years. It is interconnected to El Niño through troposphere vertical convection anomalies.
3 Marine ecosystems
3.1 North Atlantic Oscillation
The spatio-temporal dynamics of copepod populations was the first to be shown to be linked to the NAO. Four hypotheses were proposed to explain the observed correlation: a reduction of deep water volume in the Norwegian Sea where the population hibernate, a change of transport regime of the organisms from the Faeroe–Iceland channel to the North Sea, a change in food resource availability and/or a change in the competition equilibrium between species.
Similarly, the biology of small pelagic fish species such as the sardine and the herring makes them very sensitive to environmental fluctuations. Changes in their stocks have been shown to be linked to the NAO anomalies.
3.2 El Niño
The Pacific Ocean is the main site for tuna fisheries: 3.2 million cubic tons of tuna, mainly Skipjack tuna, are caught every year, i.e. about 70% of annual world tuna catch. Skipjack tuna catch per unit effort CPUE is highly correlated to ENSO indices and to the longitudinal position of the 29 °C surface isotherm. Tagging experiments have shown that Skipjack tunas perform long migrations over 1000 km. These studies show that Skipjack tunas respond to environmental conditions, as their movements are associated with the 29 °C isotherm displacements during El Niño–La Niña phenomena, as Skipjack tunas tend to stay in the most profitable waters in terms of temperature, oxygen content and food availability. These spatial shifts are consistent with existing temporal series showing a pick of Skipjack-tuna CPUE 6 to 12 months after El Niño phenomena. The temporal delay is consistent with the recruitment time for of Skipjack-tuna juveniles. However, relations between recruitment and ENSO may change among species and habitats.
3.3 Antarctic circumpolar wave
Penguins represent most of the consuming biomass in the Southern Ocean. King penguins are distributed in dense colonies around Antarctica and on sub-Antarctic islands. On Crozet Island, king penguin population is about 890 000 breeding pairs and are estimated to consume 745 000 cubic tons of mesopelagic Myctophid fish on the polar front. The polar front corresponds to the northern border of the Southern Ocean. King penguins have been shown to respond to annual surface anomalies of the polar front location and thermocline depth, but changing the duration of their foraging trips and the depth of their feeding dives.
4 Insular ecosystems
Climate changes also induce contrasted effects on geographically close areas. In western Europe, winter precipitations are positively correlated to the NAO. In Scotland, most of them occur as rain, and induce high heat loss in mammals such as Soay sheep and red deer. Positive NAO anomalies induce then high mortality in these species. In contrast, in Norway, positive NAO anomalies induce higher winter temperature, reducing snow cover, and thus increasing access to food for deer, and ultimately deer body condition.
5 Conclusion
The effects of climate on individuals and populations can be either direct through physiology (metabolism, reproduction) or indirect through the ecosystem, including prey-predator-competitor relationships.
Delayed effects are important in both marine and terrestrial ecosystems. The quality of a cohort produced a given year depends on climatic conditions during the year of their birth. Such a cohort effect has been described in codfish, but also in ungulates, in relation to NAO anomalies. Cohorts with relatively large individuals at birth have a more important contribution (qualitatively and quantitatively) than other cohorts to population dynamics.
Climatic changes tend to increase the frequency of extreme conditions that have usually ecological consequences of higher importance than mean fluctuations of the climate. For example, recent changes on environmental conditions at Kerguelen Island allowed the installation of new species such as the domestic fly when a given thermal threshold is achieved. However, this threshold is not achieved every year, avoiding the permanent installation of that species.
Climatic changes may affect organisms directly (through their physiology) and indirectly through primary and secondary producers. However, subjacent mechanisms and their consequences are usually complex, as the effects of environmental changes may depend on ecological factors.
Intrinsic and extrinsic mechanisms linked to the stochasticity of the environment typically interact in a complex manner at the scales of the populations and of the ecosystems. The understanding of these interactions is a challenge that can only be solved thanks to a consortium of ecologists, climatologists, oceanographers, statisticians and modellers. Simultaneous analysis of both ecological and oceanographic temporal series seems to be the key element for a better understanding of consequences of climatic changes on marine ecological processes. It is thus necessary and timely developing programs integrating mechanistic links between multi-scale environmental variability, individual behaviour, and population dynamics.
1 Introduction
Pour déterminer l'impact de l'augmentation de l'effet de serre, et plus généralement des changements climatiques, sur les écosystèmes, il est indispensable de mieux comprendre les phénomènes physiques, biogéochimiques et biologiques qui se déroulent au sein des océans. En effet, les océans participent à la régulation du climat par couplage avec l'atmosphère, contribuant en particulier aux phénomènes de puits de gaz à effet de serre, mais également aux transferts de chaleur entre les basses et hautes latitudes. Les océans comptent, par ailleurs, parmi les systèmes les plus productifs du globe. Or, ils font l'objet d'une exploitation croissante par les activités humaines.
Cependant, la compréhension des phénomènes marins constitue un challenge. Cela est lié, entre autres, à la difficulté d'accès aux grandes profondeurs et à certaines régions océaniques, ainsi qu'à la complexité et à la variabilité des phénomènes impliqués. Ainsi, il aura fallu attendre les années 1980 et le développement de la télémétrie satellitale pour avoir une vue globale des océans et de leur variabilité. Depuis, l'océanographie physique a progressé de telle sorte qu'il est aujourd'hui possible de prédire de manière réaliste la circulation océanique et l'état de la mer, ainsi que leurs changements. Plus récemment, la biogéochimie a fait des progrès rapides grâce à des modèles bio-géochimiques dynamiques couplés, qui permettent de relier la physique océanique et la dynamique des populations phytoplanctoniques. Toutefois, on est encore loin de comprendre exactement le fonctionnement des réseaux trophiques, en particulier au niveau des échelons supérieurs qu'affectent en dernier lieu les changements climatiques, mais également les activités humaines. Il est donc encore difficile de proposer des modèles réalistes de la dynamique des populations et des écosystèmes marins en prenant en compte ces changements.
L'impact des changements climatiques sur les écosystèmes marins peut être mis en évidence de façon directe d'après l'évolution des ressources marines. Il peut également être abordé en étudiant les changements au niveau des populations de prédateurs marins, dont certains se reproduisent à terre, et des espèces terrestres dont les apports nutritifs sont essentiellement d'origine marine. Il existe, en effet, dans la nature des phénomènes climatiques cycliques qui agissent à des échelles de temps et d'espace relativement petites comparées aux changements climatiques globaux, mais dont les conséquences sont du même ordre de grandeur que celles décrites par les modèles de changements du climat.
2 Anomalies climatiques, observatoires naturels
Les premières études se sont typiquement intéressées aux conséquences écologiques des conditions environnementales locales telles que la température, les précipitations, et la couverture neigeuse. Ces études omettaient le caractère holistique du système climatique. Cependant, un intérêt croissant est porté aux conséquences de la variabilité climatique sur le fonctionnement des écosystèmes à de grandes échelles d'espace et de temps, en particulier les phénomènes El Niño, l'oscillation Nord-Atlantique et l'onde circumpolaire antarctique. Ces phénomènes interviennent dans les grands changements climatiques et affectent l'ensemble des compartiments de l'écosystème planétaire.
2.1 El Niño
Le phénomène El Niño est associé à des changements à grande échelle du fonctionnement du système atmosphère–océan centré sur le Pacifique. L'intensité du phénomène est exprimée par un indice calculé à partir des différences de pression atmosphérique des deux côtés de l'équateur (indice des oscillations australes, El Niño Southern Oscillations, ENSO en anglais [21]). Dans les conditions normales, le bassin occidental du Pacifique équatorial présente une masse d'eau qui compte parmi les plus chaudes du globe. Un événement El Niño est associé à une anomalie atmosphérique négative, et à un réchauffement des eaux de surface équatoriales qui s'étend de la ligne de changement de date jusqu'aux côtes de l'Amérique du Sud. Les anomalies thermiques associées à El Niño peuvent atteindre +3 à +4 °C et provoquent l'affaiblissement, voire la disparition du système de remontée d'eaux profondes sur les côtes d'Amérique du Sud. La Niña est le phénomène inverse, correspondant à un refroidissement anormal des eaux de surface dans la partie orientale. El Niño et La Nina se succèdent dans le temps avec une cyclicité de 6 à 8 ans (Fig. 1).
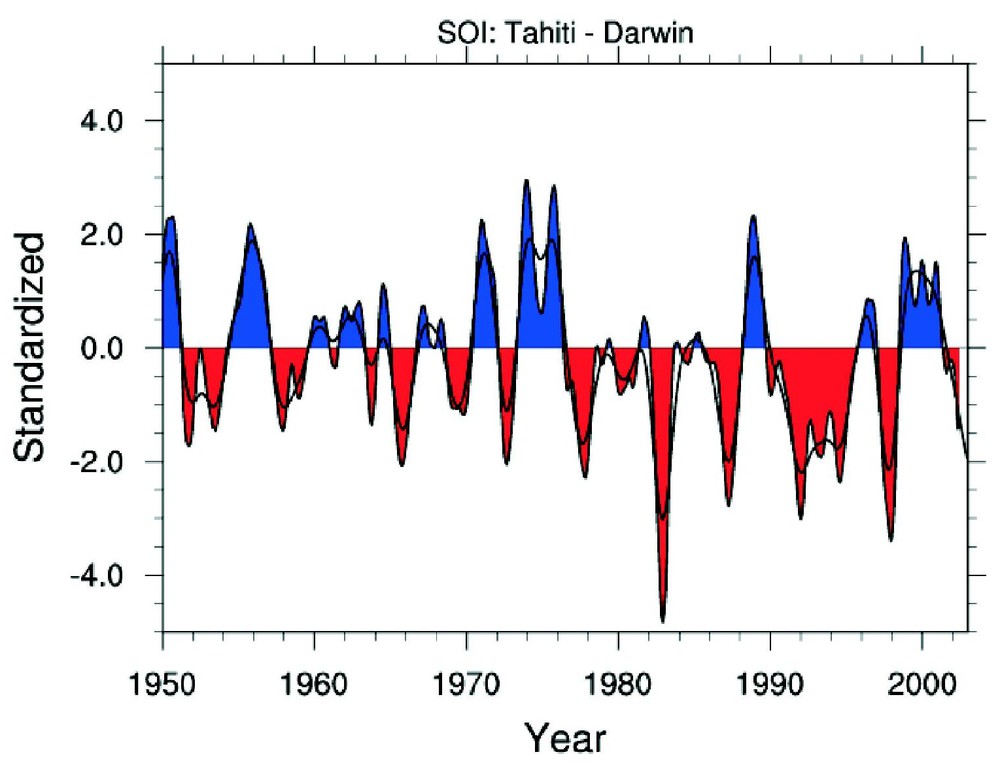

Variation temporelle de l'indice ENSO de 1950 à 2003 (en haut, d'après http://www.cgd.ucar.edu/cas/catalog/climind/soi.html) et de l'ONA de 1864 à 2003 (en bas, d'après http://www.cgd.ucar.edu/~jhurrell/noa.html).
Temporal changes of ENSO index from 1950 to 2003 (top, from http://www.cgd.ucar.edu/cas/catalog/climind/soi.html) and NOA index from 1864 to 2003 (bottom, from http://www.cgd.ucar.edu/~jhurrell/noa.html).
2.2 L'oscillation Nord-Atlantique
L'oscillation Nord-Atlantique (ONA, North Atlantic Oscillation, NAO en anglais [11]) est l'homologue atlantique de El Niño. C'est une altération des masses d'air situées entre l'anticyclone des Açores et le centre dépressionnaire situé au-dessus de l'Islande. L'ONA contrôle l'intensité des vents d'ouest entre 40°N et 60°N sur l'Atlantique Nord. Les anomalies positives de l'ONA sont associées à des vents d'ouest renforcés et décalés vers le nord, qui augmentent les précipitations et les températures sur l'Europe du Nord et les états du Sud-Est des États-Unis d'Amérique, ainsi que des anomalies sèches sur la mer Méditerranée. Des conditions quasi-opposées apparaissent pendant les anomalies négatives. Les anomalies positive/négative se succèdent dans le temps, avec une cyclicité décadaire (Fig. 1).
2.3 L'onde circumpolaire antarctique
L'onde circumpolaire antarctique (OCA, Antarctic Circumpolar Wave, ACW en anglais) a récemment été décrite comme étant un système d'anomalies de surface atmosphériques (pression, vent) et océaniques (température de surface, couverture de glace de mer) circulant d'ouest en est autour de l'océan Austral [23]. L'océan Austral est le seul domaine océanique encerclant le globe : il permet ainsi des interconnections entre les autres océans du globe, et joue un rôle capital dans la régulation du climat et dans la propagation des anomalies climatiques. L'OCA est un système d'anomalies couplées (deux anomalies positives alternant avec deux anomalies négatives) faisant donc le tour du continent Antarctique en 8 à 10 ans. L'OCA trouverait une de ses origines dans le bassin occidental du Pacifique subtropical, et semble être connectée au phénomène El Niño, décrit précédemment, grâce à des anomalies de convection verticale troposphérique [20].
3 Écosystèmes marins
3.1 L'oscillation Nord-Atlantique
Les rapports de campagnes en mer remontant au Xe siècle signalaient que des périodes propices à la pêche alternaient avec des périodes de disette. Ces mêmes rapports indiquaient également que le succès de la pêche suivait une dynamique spatiale asynchrone.
La dynamique spatio-temporelle des populations zoo-planctoniques de copépodes Calanus finmarchicus et C. helgolandicus a été parmi les premières à être mises en relation avec l'ONA [8,22]. Quatre mécanismes ont été proposés pour expliquer cette corrélation : une réduction du volume des eaux profondes de la mer de Norvège où les populations hivernent, un changement de régime du transport des organismes du chenal des ı̂les Féroé–Shetland vers la mer du Nord, une variation de la disponibilité trophique via la production phyto-planctonique et une modification de l'équilibre des relations de compétition entre les deux espèces [19]. Les jeunes stades de copépodes sont la source principale de nourriture pour les alevins de nombreuses espèces de poissons dans l'Atlantique nord, et ils régulent ainsi le recrutement des stocks de poissons.
La biologie des petites espèces pélagiques de poissons comme la sardine Sardina pilchardus, Sardinops sagax et le hareng Clupea harengus les rendent très sensibles aux fluctuations environnementales. On sait aujourd'hui que les fluctuations des stocks apparaissent sur une échelle de temps décadaire, résultant des différents régimes de vents dominants associés aux anomalies de l'ONA. Bien que les populations de sardines et de harengs soient géographiquement distantes de plusieurs milliers de kilomètres, la covariance de leur dynamique indique bien leur relation aux changements environnementaux et climatiques [1].
3.2 El Niño
Au large de l'Amérique du Sud, un fort système de remontée d'eaux profondes, associé au courant de Humboldt, induit une production biologique exceptionnellement élevée [4]. Au début des années 1970, les pêcheries péruviennes ciblant l'anchois étaient les plus productives du monde, avec des prises quotidiennes atteignant 180 000 tonnes. Toute l'industrie s'effondra en 1972, á la suite d'un phénomène El Niño induisant un déplacement des masses d'eau superficielles le long des côtes sud américaines [5]. Toutefois, une chute du recrutement durant l'année précédente suggère que l'effondrement des captures résultait à la fois d'une surexploitation et de conditions environnementales défavorables [7].
L'océan Pacifique est aujourd'hui le site principal des pêcheries thonières : 3,2 millions de tonnes de thons, principalement la bonite, Katsuwonus pelamis, sont prélevés annuellement, soit environ 70 % des captures annuelles mondiales de thonidés [12]. La bonite se distribue principalement dans la couche d'eau de mélange superficielle des bassins équatorial et subtropical. La flotte américaine réalise ses captures de surface dans la bande équatoriale [5°N–5°S], c'est-à-dire dans les eaux chaudes et à faible production primaire du Pacifique ouest.
La capture de bonites par unité d'effort (CPUE) est fortement corrélée aux oscillations ENSO et à la position méridienne de l'isotherme de surface 29 °C, qui correspond à la bordure orientale du bassin d'eau chaude mentionné précédemment (Fig. 2). Des expériences de marquage effectuées pendant des périodes El Niño et La Niña ont montré que les bonites effectuent des déplacements sur plusieurs milliers de kilomètres. Ces migrations sont associées aux mouvements de l'isotherme de surface 29 °C (Fig. 3) [14,16]. (i) Lors de situations normales, la masse d'eau froide s'étend jusqu'à la longitude 180° ; la divergence équatoriale bien établie est la source d'une forte production primaire. Dans le Pacifique ouest, la production primaire de la couche de mélange superficielle est relativement faible, du fait d'une forte stratification verticale de la colonne d'eau. La divergence équatoriale se superpose au flux zonal dirigé vers l'ouest pour créer un décalage spatial latitudinal (vers le sud) et longitudinal (vers l'ouest) des communautés planctoniques. (ii) Lors du développement d'un phénomène El Niño, le système se déplace vers l'est : la masse d'eau chaude d'ouest s'étend vers le Pacifique central, l'intensité de la divergence équatoriale diminue et la langue d'eau froide se retire vers l'est. La production secondaire, initialement dans la langue d'eau froide, se déplace dans la masse d'eau chaude pour se mélanger à la production secondaire provenant du Pacifique ouest. Sous l'influence des courants, la biomasse est agrégée dans une large bande zonale associée à la convergence le long de la bordure est de la masse d'eau chaude en expansion. La forte stratification verticale de sub-surface constitue des conditions favorables à l'alimentation de surface des bonites, qui pourraient expliquer les déplacements des populations en relation avec les déplacements de la zone de convergence. Le déplacement de la cellule de convection atmosphérique vers le Pacifique central modifie le régime des vents et engendre une remontée d'eau profonde sur la côte nord de la Nouvelle-Guinée, associée à une forte production primaire dans le secteur ouest du Pacifique. (iii) Lorsque le déplacement vers l'est s'arrête pour s'inverser, la bordure est de la masse d'eau chaude devient moins attractive pour les bonites. Le déplacement vers l'ouest du système {masse d'eau chaude/langue d'eau froide} limite un éventuel enrichissement des eaux de surface du secteur est de la masse d'eau chaude par la biomasse de la langue d'eau froide. La diminution de l'intensité de la divergence équatoriale liée à El Niño est associée à une diminution de la production secondaire avec un décalage temporel de quelques semaines (pour le zooplancton) à quelques mois (micronecton), alors que les côtes de Nouvelle-Guinée, enrichies durant El Niño, continuent de s'enrichir en zooplancton et micronecton jusqu'au retour à la situation initiale. Ainsi, l'attractivité de la zone de Nouvelle-Guinée (en terme d'habitat pour les thons) tend à augmenter, alors qu'elle diminue sur la bordure est de la masse d'eau chaude. Ce scénario pourrait expliquer les déplacements des thons vers l'ouest à cette période (Fig. 3 [14,16]). De plus, le probable enrichissement en zooplancton du Pacifique ouest lors de El Niño constitue une situation très favorable à la survie et au développement larvaire. Alors que le phénomène El Niño a des conséquences catastrophiques dans le Pacifique est, il pourrait donc faciliter le recrutement et le succès de pêche aux thons 6 à 12 mois plus tard dans le Pacifique ouest.
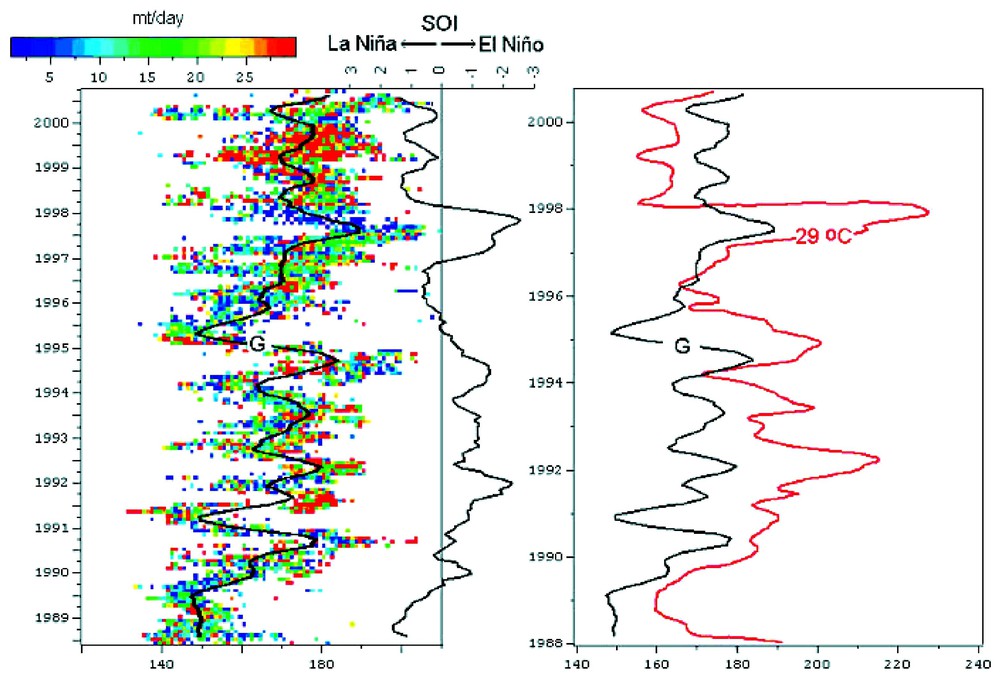
Variations temporelles (axe vertical) et longitudinales (axe horizontal) des captures par unité d'effort CPUE mensuelles moyennes (indiquées par la courbe noire, notée G) de bonites par les senneurs américains dans le Pacifique ouest équatorial dans la bande [5°N–5°S], en relation avec les oscillations ENSO (SOI) et la position longitudinale de l'isotherme de surface 29 °C. Chaque variable est lissée par une moyenne mobile sur 5 mois (d'après [16]).
Skipjack tuna CPUE of the United States purse seine fleet in the Western Equatorial Pacific Ocean between 5°N and 5°S. Temporal and longitudinal changes in mean CPUE (G solid curve) in relation to Southern Oscillation Index (SOI) and longitudinal position of the 29 °C surface isotherm (from [16]).
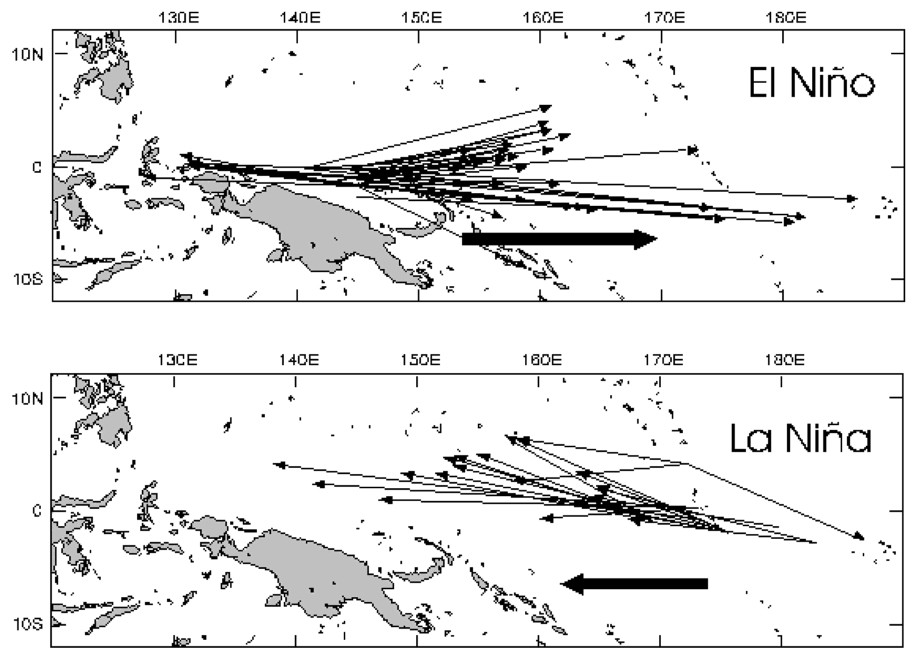
Déplacements individuels (une flèche représente un animal marqué) de bonites marquées lors d'un épisode El Niño (en haut) et d'un épisode La Niña (en bas, d'après [16]). Les flèches épaisses indiquent la direction principale des déplacements.
Skipjack tuna movements during El Niño (top) and La Niña (bottom) events (each individual movement is shown by one individual arrow). Bold arrows indicate the global direction (from [16]).
En plus de ces causes extrinsèques, les fluctuations des CPUE de bonites sont probablement liées aux fluctuations des stocks, c'est-à-dire du recrutement. L'étude des séries temporelles des CPUE montre en effet qu'un pic d'abondance apparaı̂t 6 à 12 mois après un épisode El Niño. Sachant que la première classe de taille des captures de bonites dans le Pacifique ouest correspond à 35 cm, soit un âge de 9 mois, ce schéma est cohérent avec l'hypothèse décrite précédemment d'un effet positif du phénomène El Niño sur le recrutement de bonites. La masse d'eau chaude du Pacifique ouest étant la zone de frai principale des bonites, l'augmentation des productions phyto- et zoo-planctoniques durant l'épisode El Niño contribuerait à un meilleur taux de survie des juvéniles. Le décalage temporel de 6 à 12 mois entre le pic de production primaire et celui du micronecton correspond au temps nécessaire pour le recrutement des juvéniles dans les pêches [14].
Les relations existant entre le recrutement et les oscillations ENSO varient selon les espèces et selon leurs habitats. El Niño induit des conditions favorables au recrutement de la bonite dans le Pacifique ouest, mais défavorables à celui des albacores Thunnus alalunga [15], comme cela a également été décrit pour les anchois du Pérou.
3.3 L'onde circumpolaire antarctique
Les manchots représentent l'essentiel de la biomasse consommante des écosystèmes de l'océan Austral [6]. Les manchots royaux Aptenodytes patagonicus se regroupent en colonies pouvant atteindre plusieurs millions d'individus tout autour du continent Antarctique et sur les ı̂les subantarctiques. Aux ı̂les Crozet, secteur austral de l'océan Indien, où la population est la plus grande du monde (890 000 couples reproducteurs), les manchots royaux consomment principalement des poissons méso-pélagiques de la classe des Myctophidés (745 000 tonnes par an [9]) sur le front polaire antarctique [2,10]. Le front polaire, situé au sud des ı̂les Crozet, détermine la limite nord de l'océan Antarctique. Toutefois, les anomalies de surface annuelles influencent la position du front polaire et la profondeur de la thermocline : par exemple, un réchauffement des eaux de surfaces augmente la distance au site d'alimentation et provoque l'enfoncement de la thermocline. Les manchots royaux sont alors contraints d'adapter la durée de leur voyage alimentaire en mer, ainsi que leur effort de plongée afin d'assurer leur reproduction (Bost, non publié, et [13]).
4 Écosystèmes continentaux
Les changements climatiques induisent également des effets contrastés sur des régions géographiques à échelle spatiale relativement faible. En Europe occidentale, les précipitations hivernales sont positivement corrélées à l'ONA. En Écosse, la majeure partie de ces précipitations tombe sous forme de pluies, qui causent de fortes déperditions de chaleur chez les mammifères, tels que les moutons Soay Ovis aries et cerfs Cervus elaphus. Les anomalies positives de l'ONA induisent donc une forte mortalité dans ces populations. En Norvège, les températures hivernales relativement plus élevées qui résultent des anomalies positives de l'ONA réduisent le couvert neigeux, améliorant l'accès à la ressource pour les cervidés [3]. À l'inverse de ce qui est observé en Écosse, les anomalies positives de l'ONA augmentent ainsi la survie et la condition corporelle des ongulés de Norvège [17,18].
5 Conclusions
Les exemples présentés ici montrent l'évidence de certaines relations entre la dynamique des écosystèmes marins et les changements environnementaux, pour certains associés à des changements climatiques, mais également le manque d'éléments nécessaires à une compréhension exacte des mécanismes impliqués dans ces phénomènes. L'effet du climat sur les individus et les populations peut être direct, via la physiologie des animaux (métabolisme et reproduction), ou indirect, via l'écosystème, incluant les relations proies–prédateurs–compétiteurs.
- (i) Les effets retardés du climat sont importants pour les écosystèmes marins, et ont également de fortes répercussions sur les écosystèmes terrestres. La qualité de la cohorte produite une année donnée dépend des conditions climatiques au cours de l'année de sa naissance. Un tel effet cohorte a été décrit chez les morues, mais également chez les ongulés, et a été mis en relation avec les anomalies de l'ONA. De plus, les cohortes composées d'individus relativement gros dès leur naissance ont une survie et une contribution quantitative et qualitative plus importante que les autres cohortes à la dynamique des populations.
- (ii) Les changements climatiques tendent à augmenter la fréquence des conditions extrêmes, qui ont souvent des conséquences écologiques plus importantes que les fluctuations moyennes du climat. Par exemple, les modifications récentes des conditions climatiques des ı̂les Kerguelen ont permis l'établissement de nouvelles espèces, telles que la mouche domestique Calliphora vicina, qui s'installe au-delà d'un seuil thermique de 611 degrés-jours annuels. Toutefois, ces conditions seuil ne se présentent pas chaque année et ne permettent pas l'installation définitive de l'espèce (Y. Frenot, comm. pers).
- (iii) Les changements climatiques peuvent affecter les organismes directement (par leur physiologie) ou indirectement via les producteurs primaires et secondaires. Toutefois, les mécanismes sous-jacents et leurs conséquences sont souvent complexes, car les effets de ces changements environnementaux peuvent dépendre de facteurs écologiques.
Les mécanismes intrinsèques et extrinsèques liés à la pêcherie de l'environnement interagissent typiquement de manière complexe à l'échelle des populations et des écosystèmes. La compréhension de ces interactions est un challenge dont la résolution implique un consortium d'écologistes, climatologistes, océanographes, statisticiens et modélisateurs. Ainsi, l'analyse simultanée des séries temporelles écologiques et océanographiques est un élément clé de la compréhension des conséquences des variations climatiques sur les mécanismes écologiques marins. Il est donc nécessaire de développer des programmes intégrant les liens mécanistiques existant entre la variabilité environnementale multi-échelle, le comportement individuel et la dynamique des populations. De tels programmes offriraient des résultats dont les implications iraient au-delà de la seule écologie.
Remerciements
Les auteurs remercient C.A. Bost, Y. Frenot et P. Lehodey pour leurs communications personnelles.
IYG bénéficiait d'un financement Marie Curie du programme de la Communauté européenne, n° HPMF-CT-2000-00816.