

Abridged English version
Recently, some scientists demonstrated coral reef fish larvae had swimming abilities sufficient to control their pattern of oceanic dispersion and their return to adult habitat. However, these swimming abilities will be useful, only if larvae can detect a suitable habitat at settlement stage, because it seems unlikely that successful settlement is solely a matter of chance. Thus, one of the great mysteries of coral reef fish ecology is how fish larvae locate the relatively rare patches of coral reef habitat on which they settle and ultimately reside as adults. The answer must lie partly in the sensory abilities of fish. Two research topics must be distinguished: (i) how do pelagic larvae recognise the island to colonise, and (ii) how do reef larvae recognise their settlement location? This manuscript examines the sensory abilities of coral reef fish larvae in the recognition of their location of settlement at Moorea Island (French Polynesia).
Larval recognition of settlement location can be based on the detection of conspecifics already settled or of types of habitat (e.g., shape of coral colony, odour of anemone) using emissions of visual, chemical and mechanical cues. These cues can be detected by five senses of fish: visual cues by sight, chemical cues by smell or taste, and mechanical cues (sound and vibratory waves) by hearing (sound is captured by otoliths) or vibratory sense (vibrations generated by movement of organisms in water are captured by lateral line). This study, carried out in aquaria and in situ, investigated the senses (sight, smell, taste, hearing, vibratory sense) used by larvae in the recognition of cues from habitat or conspecifics, and estimated the distance of detection of each sense.
The results in aquaria on 12 species demonstrated that 83% of species detected their settlement location due to the presence of conspecifics and not based on the type of habitat. Among the different senses tested, only sight, smell and vibratory sense were used in this detection. Thus among the 12 species sampled, sight is used seven times, smell seven times, vibratory sense five times, and taste and hearing never. Among these species, only two species (Parupeneus barberinus and Ctenochaetus striatus) used three senses (view, smell and vibratory senses), five (Myripristis pralinia, Lutjanus fulvus, Chromis viridis, Thalassoma sp. and Acanthurus triostegus) used two senses, three (Gymnothorax sp., Chrysiptera leucopoma and Rhinecanthus aculeatus) used one sense and two (Pomacentrus pavo and Canthigaster janthinoptera) apparently used no sense. The in situ experiment carried only out on Chromis viridis demonstrated that the distance of detection by sight was of 75 cm, and by smell was of 375 cm. This result has been validated by the location of settlement habitat. Indeed, when the distance of detection was less than 75 cm, larvae of Chromis viridis did not consider the direction of current in their choice of settlement habitat. By contrast, when the distance was greater than 75 cm, they only settled on habitat located upper current.
The hypothesis that fish had all senses functional at larvae stage is now accepted. Thus, for the two species that do not migrate toward conspecifics (Pomacentrus pavo and Canthigaster janthinoptera), the reasons could be due to no attraction of larvae in situ and/or stress of life in aquaria. This stress could inhibit the migratory behaviour of larvae even when larvae detect conspecifics, or the emission of chemical and mechanical cues from conspecifics. The hypothesis of no attraction of larvae in situ would mean that larvae detect their settlement location by other factors such as food resources, or the presence of predators.
The 10 species that detected conspecifics used only three senses: sight, smell and vibratory sense. Taste was not used due to the chemical composition and/or too weak concentration of pheromones produced by conspecifics. Indeed, some studies have demonstrated that smell was more sensitive than taste (which required a high concentration in pheromones). Other studies demonstrated that, according to the chemical composition of pheromones, fish used either smell or taste to detect an object (e.g., prey, predators, habitat). At present, the author tries to identify by High Performance Liquid Chromatography the nature of pheromones to test this hypothesis (Lecchini, in prep.). Hearing has not been used by reef larvae to detect conspecifics. Yet oceanic larvae use it to detect islands due to the sound of waves on reef crest of islands. A hypothesis would be that conspecifics do not generate sound in aquaria. Indeed, the sound generation that may be specific for each species is an active and voluntary process at the difference of emission of vibratory waves. The pelagic larvae can detect islands by sound because the waves always generate noise on reef crests. For reef larvae to recognise in situ conspecifics by hearing, the conspecifics should generate sound when larvae arrive on reef crests. Thus, there would be a temporal correlation between production of sound by conspecifics and the pattern of colonisation of larvae. This hypothesis is, however, difficult to accept. Overall, reef larvae did not use hearing to detect their location of settlement because conspecifics did not generate noise in aquaria.
The determination of which senses (sight, smell, vibratory sense) are used for the detection of settlement location could be due to a combination of different processes: imprinting an odour of parental habitat at birth, ‘training of conspecific’ in open water, and developmental state of sensory organs of each species. However, environmental conditions of larvae after colonisation stage are different from these of aquaria. In situ, larvae must not only detect the cues from conspecifics but also those from predators and congeners. Thus, future studies in aquaria should be validated by in situ experiments to test other environmental cues impacting the larvae during the settlement stage. Overall, sensory recognition of settlement location is still an open topic. The interface of research in behaviour and ecology is essential to explain or predict the processes that regulate responses of animals to aspects of their environment and of their distribution in space and time.
1 Introduction
À la suite de l'attouchement d'une tige de céréale et des vibrations qu'il entraı̂ne, le puceron vert de l'avoine (Sitobion avenae) se laisse immédiatement tomber au sol en une réaction de fuite devant un prédateur [1]. La fuite ou le camouflage, dès qu'est perçu un prédateur, l'affirmation de son rang social, la détection puis la localisation de la source alimentaire appropriée, du partenaire sexuel ou du site d'installation, sont autant de manifestations tangibles de l'adaptation des organismes à leur écosystème. Elles sont fondées sur l'information et supposent la sollicitation quasi-constante d'un équipement sensoriel qui discrimine, enregistre, compare, ordonne et coordonne les informations reçues. Afin que cette analyse soit efficace, le comportement des organismes doit être coordonné dans le temps et contrôlé dans l'espace [2]. Tout déplacement de quelques centimètres ou de milliers de kilomètres nécessite donc, de la part des animaux, un comportement orienté. Certains auteurs ont récemment démontré que les larves de poissons coralliens avaient des capacités natatoires suffisantes pour contrôler leur patron de dispersion dans l'océan, ainsi que leur retour jusqu'à l'habitat parental [3–5]. Cependant, comme le succès de l'installation dans l'habitat parental n'est pas uniquement dû à la chance [6], les capacités natatoires des larves ne leurs seront utiles que si elles détectent le récif à coloniser. Ainsi, l'une des grandes énigmes de l'écologie des poissons coralliens est de savoir comment les larves localisent les relativement rares habitats coralliens sur lesquels elles peuvent s'installer. La réponse doit se trouver dans l'univers sensoriel des poissons [7]. Il semble important de distinguer deux axes de recherche dans cette thématique de la reconnaissance sensorielle : (i) comment les larves océaniques détectent-elles l'ı̂le à coloniser et (ii) comment les larves colonisantes détectent-elles l'habitat d'installation ? De plus en plus d'études s'orientent actuellement sur la reconnaissance sensorielle de l'ı̂le à coloniser [5,8,9]. Cet article traite des capacités sensorielles des larves de poissons coralliens dans la détection de leur habitat d'installation.
Les poissons coralliens ont généralement un cycle de vie complexe, avec une phase larvaire océanique et pélagique, de 10 à 120 jours selon les espèces, suivie d'une phase récifale relativement sédentaire pour les juvéniles et les adultes [10]. Durant la phase océanique, les larves s'éloignent plus ou moins loin de leur ı̂le natale grâce aux courants [11] et/ou à une dispersion active [4]. Ensuite, elles retournent vers une ı̂le corallienne (d'origine ou non) pour continuer leur développement en juvéniles, puis en adultes. Les larves colonisent cette ı̂le de nuit et au niveau de la crête récifale (phase de colonisation, [12]). Dans les heures qui suivent cette colonisation, les larves en cours de métamorphose choisissent un habitat (phase d'installation, [13]) selon principalement deux critères : les caractéristiques structurales de l'habitat et les interactions intra- et inter-spécifiques avec les poissons déjà installés [14–16]. Cette phase d'installation a été largement étudiée chez les poissons coralliens [6], mais peu d'informations sont disponibles sur les réponses comportementales des larves aux signaux sensoriels attracteurs du milieu [17]. Ces signaux qui permettent aux larves de détecter leur lieu d'installation, peuvent être émis par l'habitat corallien (forme spécifique d'une colonie corallienne ou phéromone spécifique d'une anémone) ou par les conspécifiques (poissons au stade juvénile ou adulte de la même espèce que les larves colonisantes, déjà présents sur le lieu d'installation). Sweatman [14] a étudié in situ la reconnaissance olfactive des lieux d'installation chez deux espèces de Pomacentridae (Dascyllus aruanus et Dascyllus reticulatus). Il a utilisé un système de pompes qui collectaient l'eau de colonies coralliennes contenant des conspécifiques et l'injectaient sur des colonies coralliennes dépourvues de poissons. Il a ainsi démontré que les colonies coralliennes sur lesquelles l'eau était injectée présentaient une installation en juvéniles de Dascyllus aruanus et de Dascyllus reticulatus supérieure à celle des colonies dépourvues de ce système. Booth [18] a démontré en aquarium que la vue jouait un rôle dans le choix du lieu d'installation pour Dascyllus albisella (Pomacentridae). Enfin, Elliott et al. [19] ont démontré in situ que les larves d'Amphiprion (poisson clown) détectaient par l'odeur et jusqu'à une distance de 8 m, la présence d'anémones. Seules ces trois études ont traité de la reconnaissance sensorielle des lieux d'installation chez les poissons coralliens.
L'objectif de mon étude était d'estimer par des expériences en aquarium et in situ les capacités sensorielles des larves de poissons coralliens lors de la recherche du lieu d'installation de leur espèce. Cette recherche peut donc correspondre à la détection des conspécifiques ou d'un type d'habitat par leurs émissions de signaux visuel (forme d'une colonie corallienne), chimique (odeur d'une anémone) et mécanique (ondes vibratoires ou sonores des poissons). Ces signaux pourront être reconnus par cinq sens des larves : le signal visuel par la vue, le signal chimique par l'odorat (les narines sont responsables de ce sens) et par le goût (des cellules sensorielles au niveau de la bouche permettent cette reconnaissance), et le signal mécanique (ondes sonores ou vibratoires) par l'ouı̈e (le son est capté au niveau des otolithes) et par la perception des ondes vibratoires (les vibrations sont captées par la ligne latérale et sont produites par tout mouvement d'organismes dans l'eau). Il est à noter que la perception du champ électrique est un sens présent chez certaines espèces de poissons [7,20], mais ces espèces sont absentes de notre zone d'étude de Moorea, en Polynésie française [21].
2 Matériel et méthodes
2.1 Modèle biologique de l'étude
Aucune liste d'espèces cibles n'a été préalablement définie pour cette étude. Elle dépend des captures quotidiennes effectuées par des filets de crête [12]. Ces filets permettent la capture des larves de poissons juste avant qu'elles ne pénètrent dans le lagon pour s'installer puisque les filets sont posés sur la crête récifale de la côte nord de Moorea. Les conspécifiques, utilisés comme émetteurs de signaux, sont des juvéniles qui ont été capturés aux filets de crête et maintenus entre 15 et 21 jours dans des aquariums.
2.2 Protocole d'étude des expériences sensorielles en laboratoire
2.2.1 Description et principe du système expérimental
L'objectif de l'expérience est de tester séparément les trois signaux (signal visuel, chimique et mécanique) afin de savoir si les larves peuvent les utiliser dans la détection des lieux d'installation. Cette expérience n'ayant jamais été réalisée auparavant, il a fallu concevoir un système expérimental particulier (Fig. 1). Ce système est constitué d'un aquarium à triple choix (A, B ou C), comprenant cinq compartiments (A, B, C, D, E), auquel sont ajoutés, de part et d'autre, deux aquariums à un seul compartiment (appelés bacs).
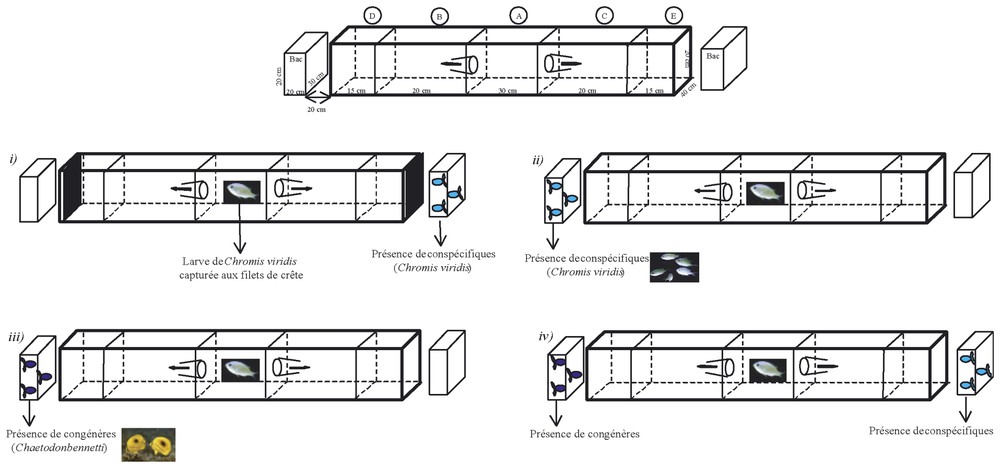
Description de l'aquarium et protocole expérimental de l'étude sensorielle permettant de mettre en évidence une attraction spécifique des larves vers les conspécifiques. Exemple dans la reconnaissance visuelle des conspécifiques par les larves de Chromis viridis. Les larves sont introduites dans le compartiment central (noté A) et peuvent y rester ou migrer dans les deux compartiments adjacents (notés B et C) grâce à des entonnoirs (système d'anti-retour). Les deux autres compartiments situés à l'extrémité (notés D et E) sont isolés des précédents par une paroi en verre. À cet aquarium à cinq compartiments, deux autres aquariums à un compartiment (appelés bacs) sont ajoutés de part et d'autre. Le principe de l'expérience est d'introduire des larves dans le compartiment central et d'observer dans quel compartiment elles se situent (A, B ou C) deux minutes après.
Pour tester le signal visuel, les conspécifiques ou un bloc de corail (représentant l'habitat corallien d'installation) sont placés dans les bacs. Puisque les bacs et l'aquarium ne sont pas en contact et reposent sur des plaques de polystyrène, les ondes mécaniques et les molécules chimiques ne peuvent pas se propager jusqu'aux larves, qui sont placées dans le compartiment central de l'aquarium [22]. Dans cette configuration, le signal visuel est seul responsable de la migration des larves.
Le signal mécanique est uniquement testé à partir des conspécifiques. Ces derniers sont introduits dans le compartiment D ou E (Fig. 1). Les vitres qui séparent les compartiments D de B et E de C sont obscurcies par une plaque de plastique opaque afin d'éliminer le signal visuel. De plus, les molécules chimiques ne peuvent pas diffuser entre ces compartiments. Dans cette configuration, le signal mécanique est seul responsable de la migration des larves. Si la migration a lieu, elle peut être déclenchée par l'audition des larves ou par leur perception des ondes vibratoires. Afin de ne tester que l'audition, les larves sont alors immergées durant 24 h dans des bains à streptomycine (44 mg l−1, [23]). La streptomycine a pour fonction de bloquer temporairement les cellules sensorielles de la ligne latérale [24,25].
Pour tester le signal chimique, le compartiment B ou C est rempli indépendamment des deux autres compartiments (une plaque de plastique est positionnée devant l'entonnoir correspondant afin de l'obturer) avec de l'eau dans laquelle des conspécifiques (cinq conspécifiques dans trois litres d'eau) ou un bloc corallien (colonie corallienne sphérique de 6 cm de rayon dans trois litres d'eau) ont été immergés pendant 6 h. Lorsque les larves sont introduites dans le compartiment central (rempli avec de l'eau pure), la plaque de plastique est soulevée et les molécules chimiques peuvent diffuser. Si la migration a lieu, elle peut être déclenchée par l'olfaction ou la gustation. Afin de ne tester que la gustation, les narines des larves sont temporairement obstruées par l'ajout de vaseline [25].
Afin que seuls les signaux sensoriels testés soient responsables de la migration des larves, l'expérience a été réalisée dans une pièce du laboratoire isolée de la lumière extérieure (pièce calfeutrée) et des bruits extérieurs (expérience menée de nuit). Cette expérience a été cependant réalisée en présence de lumière intérieure, avec une répartition homogène des néons dans la pièce. L'eau des aquariums est prélevée dans le lagon grâce à un système de pompes. Cette eau est alors décantée dans des bacs puis subit une filtration sur un bac à sable (pores à 50 μm). Ensuite, elle est stérilisée par rayonnements ultraviolets. Cette eau relativement pure (sans micro-organisme et sans matière organique) est ainsi renouvelée à chaque expérience.
2.2.2 Traitement statistique
L'aquarium à cinq compartiments a été conçu afin d'étudier dans quel compartiment (A, B ou C) se situent les larves deux minutes après leur introduction dans l'aquarium. Il faut donc comparer une distribution théorique (répartition homogène des larves dans les trois compartiments) à une distribution observée. Le test du chi2 a été choisi [26]. Pour appliquer ce test, l'expérience doit être réalisée avec plus de 15 larves (80 % des effectifs théoriques ) pour chaque signal testé. De plus, les réponses des larves doivent être indépendantes les unes des autres (pas d'effet de groupe). Une larve par aquarium doit donc être utilisée (10 aquariums et 20 bacs ont été construits pour l'étude). Ainsi, 20 larves sont testées individuellement dans la reconnaissance de chaque signal (parfois 16 larves sont utilisées à cause de captures insuffisantes). Les trois signaux des conspécifiques ou les deux signaux des blocs coralliens sont testés durant la même nuit respectivement à partir d'un lot de 60 ou de 40 larves capturées la nuit précédente par des filets de crête. L'étude a été réalisée de février à mai 2002, période durant laquelle trois expériences ont pu être effectuées, sur 12 espèces.
2.2.3 Expérience 1 : détection des signaux visuels, chimiques et mécaniques émis par les conspécifiques
Le deuxième impératif de l'étude, après l'impératif statistique, est de démontrer que l'attraction est spécifique. Les larves doivent reconnaı̂tre les signaux émis par les juvéniles de leur espèce et non ceux émis par une quelconque espèce. Pour démontrer cette attraction spécifique, le protocole d'étude est le suivant (Fig. 1).
(i) Expérience de vérification de l'isolation de l'aquarium. Des conspécifiques (au nombre de cinq) sont placés dans un des deux bacs. Une plaque de plastique opaque est positionnée entre ce bac et l'aquarium afin d'inhiber la diffusion du signal visuel. Une plaque identique est placée entre l'autre bac et l'aquarium. Les larves sont alors introduites dans les compartiments centraux des aquariums, et le compartiment dans lequel est présente chaque larve après deux minutes est noté. Un test du chi2 est réalisé afin de savoir si la distribution observée est identique à la distribution théorique (6,66 larves par compartiment) ou si les 20 larves sont majoritairement restées dans les compartiments centraux (pas de migration). Si tel est le cas, aucun signal n'est émis des conspécifiques jusqu'aux larves (signal électrique, magnétique, etc.). L'étude peut continuer.
(ii) Expérience de détection des conspécifiques (exemple du signal visuel). Les conspécifiques (au nombre de cinq) sont placés dans un des deux bacs. Les larves sont alors introduites dans les compartiments centraux des aquariums, et le compartiment dans lequel est présente chaque larve après deux minutes est noté. Un test du chi2 est réalisé pour démontrer l'attraction vers les conspécifiques.
(iii) Expérience de non répulsion vis-à-vis des congénères (exemple du signal mécanique). Les congénères sont des larves capturées aux filets de crête, maintenues en aquarium entre 15 et 21 jours et appartenant à une autre espèce que celle des larves testées. Les congénères sont placés (au nombre de cinq) dans un des deux compartiments D ou E. Les larves sont alors introduites dans les compartiments centraux des aquariums, et le compartiment dans lequel est présente chaque larve après deux minutes est noté. Un test du chi2 est réalisé. L'espèce de congénère est gardée dans la suite de l'étude si et seulement si aucune répulsion significative de la part des larves n'est observée, c'est-à-dire que les larves ne se situent pas majoritairement dans le compartiment (B ou C) situé à l'opposé du compartiment contenant les congénères.
(iv) Expérience de détection spécifique des conspécifiques (exemple du signal chimique). Le compartiment B est rempli avec de l'eau ayant contenu des conspécifiques, et le compartiment C avec de l'eau ayant contenu des congénères. Le compartiment central est rempli avec de l'eau pure. Les larves sont alors introduites dans les compartiments centraux des aquariums et l'observateur note le compartiment dans lequel est présente chaque larve deux minutes après avoir retiré les plaques de plastique servant d'isolant entre les compartiments A, B et C. Un test du chi2 est réalisé afin de savoir si les larves détectent spécifiquement les conspécifiques.
Ce protocole expérimental est utilisé pour les trois signaux. Si la détection spécifique est significative pour le signal chimique ou mécanique, il faut alors séparer l'olfaction de la gustation et l'audition de la perception des ondes vibratoires. L'expérience (iv) est réalisée de nouveau, sauf que les narines des larves sont obstruées par l'ajout de vaseline ou que leur ligne latérale est inhibée par la streptomycine.
2.2.4 Expérience 2 : détection des signaux visuel et chimique émis par les blocs coralliens
L'objectif de l'expérience est identique à celui de l'expérience 1, sauf que les signaux sont émis par un bloc corallien (colonie corallienne vivante ou morte sphérique de 6 cm de rayon, ou débris coralliens formant un amas sphérique de 6 cm de rayon). Ces blocs sont récoltés dans le lagon et correspondent aux habitats d'installation des larves testées. Le protocole expérimental est identique à celui de l'expérience 1, sauf que le signal mécanique n'est pas testé.
2.2.5 Expérience 3 : migration préférentielle des larves vers les signaux des conspécifiques ou vers ceux des blocs coralliens
L'objectif de l'expérience est de savoir si les larves sont attirées préférentiellement par les signaux des conspécifiques ou par ceux des blocs coralliens. Le protocole expérimental est identique à celui de l'expérience 1, sauf que les congénères sont remplacés par un bloc corallien.
2.3 Protocole d'étude de l'expérience sensorielle in situ
L'étude est réalisée sur un fond sableux en bordure de plage (sur la côte nord de Moorea), dépourvu de tout organisme (exceptée l'endofaune), d'une profondeur de 60 cm et avec un courant faible. L'étude a été effectuée de nuit au cours d'une semaine de pleine lune (avril 2002) afin de pouvoir suivre sous l'eau, les larves sans lumière artificielle. Sur ce banc de sable, un cercle est tracé, dont le centre est matérialisé par un piquet (Fig. 2). À ce piquet est attachée une boı̂te en plastique transparente, dans laquelle chaque larve testée est introduite. Au bout d'une minute, la boı̂te est ouverte et la larve peut migrer. À la périphérie du cercle, 12 arcs de cercle de même dimension sont matérialisés par des piquets, six sont occupés par des colonies coralliennes vivantes de Porites rus sur lesquelles des conspécifiques sont installés (entre 40 et 60 par arc de cercle) et les six autres sont inoccupés. Cette expérience in situ a pour objectif d'estimer la distance de détection des sens en comptant le nombre de larves qui s'installent sur un arc de cercle occupé (ou sortent du cercle) selon la longueur des arcs de cercle.
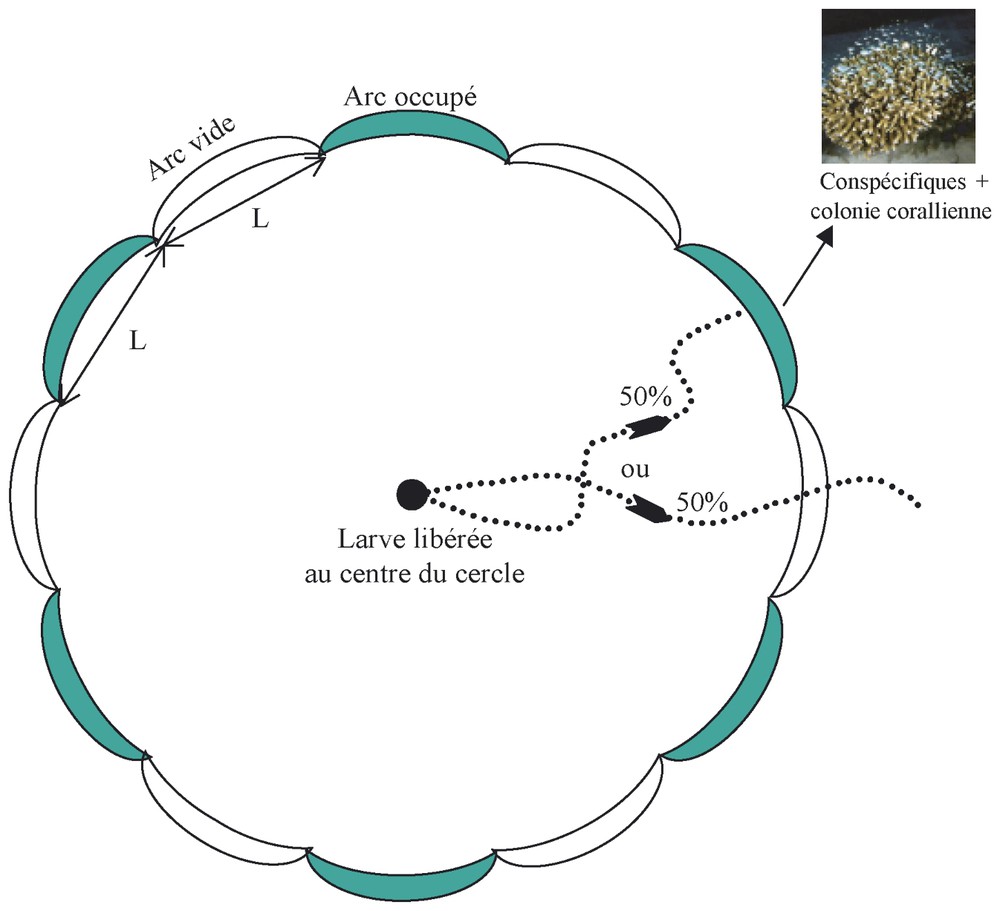
Principe de l'expérience in situ afin d'estimer la distance de détection des sens responsables de la reconnaissance des lieux d'installation. Un cercle est tracé sur fond sableux ; son centre est matérialisé par un piquet (rond noir). Ce cercle est découpé en 12 arcs de cercle de même dimension (L) dont six sont occupés par des colonies coralliennes vivantes de Porites rus et des conspécifiques (arcs de cercle foncés) et six sont vides (arcs de cercle transparents). La post-larve libérée au centre de ce cercle à une probabilité égale à 50% de s'installer sur les arcs de cercle occupés en absence d'attraction sensorielle. Différentes longueurs d'arc de cercle sont ainsi testées.
Différentes longueurs d'arc de cercle sont testées. La première longueur est de 50 cm. À cette longueur, les habitats coralliens, constitués d'une à trois colonies coralliennes (plus les conspécifiques), occupent la totalité de la longueur de l'arc de cercle. Si l'attraction sur les arcs de cercle occupés est significative (test du chi2), alors la distance de détection du sens testé est comprise entre 0 et 25 cm. La seconde longueur testée est de 1 m. À cette longueur, les habitats coralliens occupent toujours une longueur de 50 cm et les 25 cm laissés libres de chaque côté de l'habitat correspondent à la distance maximale de détection précédemment testée. Si l'attraction est significative, alors la distance de détection du sens est comprise entre 25 et 75 cm. L'étude est ainsi poursuivie avec des longueurs de 2 m (distance de détection comprise entre 75 et 175 cm) et de 4 m (distance de détection comprise entre 175 et 375 cm).
Comme l'étude est réalisée in situ, les différents signaux issus des lieux d'installation (conspécifiques+habitat) ne peuvent être individualisés. Les larves sont alors testées dans cinq états physiologiques : (i) les larves possèdent tous leurs sens, (ii) la vue est inhibée par l'ajout de sparadraps sur les yeux (le sparadrap est collé minutieusement sur les joues afin de ne pas abı̂mer les yeux des poissons), (iii) l'odorat est inhibé par l'ajout de vaseline, (iv) la perception des ondes vibratoires est inhibée par l'ajout de streptomycine et (v) tous les sens sont inhibés, sauf le goût et l'ouı̈e. L'inhibition des sens est effectuée quatre heures avant l'expérience afin que les larves « s'adaptent » à leur état physiologique (toutes les larves sont ensuite recapturées et maintenues quelques jours en aquarium avant d'être relâchées). Ainsi, pour chaque longueur, 16 larves de chaque état physiologique sont testées individuellement durant une même nuit. Deux réplicats sont effectués par longueur afin de réduire la variabilité pouvant provenir des conditions environnementales du milieu et de la condition physiologique des larves. Le nombre de larves qui s'installent sur les arcs de cercle occupés est noté. Aucune prédation n'est observée durant l'étude. Un test du chi2, ajusté par les corrections de Yates [26], est réalisé sur chaque réplicat de chaque longueur de chaque état physiologique avec, comme distribution théorique, une probabilité de 50 % que les larves sortent du cercle. Cette expérience n'a pu être réalisée que sur une seule espèce, Chromis viridis.
3 Résultats
3.1 Expériences sensorielles en laboratoire
L'expérience 1 de l'étude en laboratoire a été réalisée sur 12 espèces de poissons coralliens réparties en neuf familles (Tableau 1). Seuls les résultats de la détection spécifique des conspécifiques (présence simultanée des conspécifiques et des congénères) sont présentés pour chaque sens (Tableau 1). Cependant, pour chaque larve testée, l'aquarium était parfaitement isolé (expérience i) et les congénères n'ont pas provoqué de répulsion significative des larves (expérience iii). De plus, si la détection spécifique (expérience iv : présence simultanée de conspécifiques et de congénères) était significative, la détection des conspécifiques (expérience ii : présence uniquement de conspécifiques) l'était aussi et vice-versa. Ainsi, aucune variabilité n'a été observée entre les différentes étapes du protocole expérimental. En utilisant le test du chi2 à un seuil de probabilité de 5 %, la détection devient significative à partir de 12 larves (pour 20 larves testées) ou de 10 larves (pour 16 larves testées) migrant dans le compartiment d'où provient le signal sensoriel. Sur les 12 espèces étudiées, la vue et l'odorat ont été utilisés sept fois, le sens vibratoire cinq fois et le goût et l'ouı̈e jamais. Sur ces 12 espèces, deux (Parupeneus barberinus et Ctenochaetus striatus) utilisent trois sens (vue, odorat et sens vibratoire), cinq (Myripristis pralinia, Lutjanus fulvus, Chromis viridis, Thalassoma sp. et Acanthurus triostegus) en utilisent deux, trois (Gymnothorax sp., Chrysiptera leucopoma et Rhinecanthus aculeatus) en utilisent un et deux (Pomacentrus pavo et Canthigaster janthinoptera) n'en utilisent aucun.
Études sensorielles en aquarium réalisées sur 12 espèces de poissons coralliens. Les données correspondent au nombre de larves par compartiment et lorsque l'attraction est significative, elles sont mises en gras (test du chi2 à 5%). Les lettres « nt » signifient non testé, car la détection chimique ou mécanique des conspécifiques, ou celle de l'habitat, était non significative. Dans l'expérience 1, l'attraction vers les conspécifiques correspond au compartiment B et celle vers les congénères au compartiment C. Pour l'expérience 2, l'attraction vers leur habitat d'installation (identique à celui observé in situ à Moorea) correspond au compartiment B et celle vers un autre habitat au compartiment C. Leur habitat d'installation est cité avec la forme de la colonie et son état – vivant ou mort. Pour Pomacentrus pavo, l'espèce de corail est Porites lobata, myrmidonensis ou australiensis. Pour Thalassoma sp., c'est Pocillopora verrucosa ou meandrin. Dans l'expérience 3, l'attraction vers les conspécifiques correspond au compartiment B et celle vers l'habitat au compartiment C
Dans l'expérience, aucune des cinq espèces étudiées n'est attirée par le signal visuel ou chimique des habitats d'installation. L'expérience 3 confirme que les signaux attracteurs (visuel et chimique) proviennent des conspécifiques et non des blocs coralliens. Il est à noter que les expériences 1 et 3, réalisées avec des larves issues de captures décalées dans le temps (durée de l'étude : quatre mois) montrent une absence de variabilité dans les capacités sensorielles des larves. En effet, l'utilisation des sens pour une même espèce est homogène entre les deux expériences. Seule, Chromis viridis présente une variabilité, puisque la vue n'est pas utilisée dans l'expérience 1, tandis que dans l'expérience 3, elle l'est.
3.2 Expérience sensorielle in situ
L'étude a été réalisée sur Chromis viridis, dont les larves détectent en laboratoire les conspécifiques par l'odorat, le sens vibratoire et parfois la vue. In situ, les larves, avec tous leurs sens fonctionnels, détectent significativement les lieux d'installation (conspécifiques et/ou habitats coralliens) jusqu'à une distance d'au moins 375 cm, distance la plus grande testée (Fig. 3). Lorsque le goût et l'ouı̈e sont les seuls sens fonctionnels, il n'y a pas d'installation significative, quelle que soit la distance de détection. À l'inverse, l'inhibition de la ligne latérale n'induit jamais une installation non significative. Les larves détectent donc les lieux d'installation par la vue et/ou l'odorat.
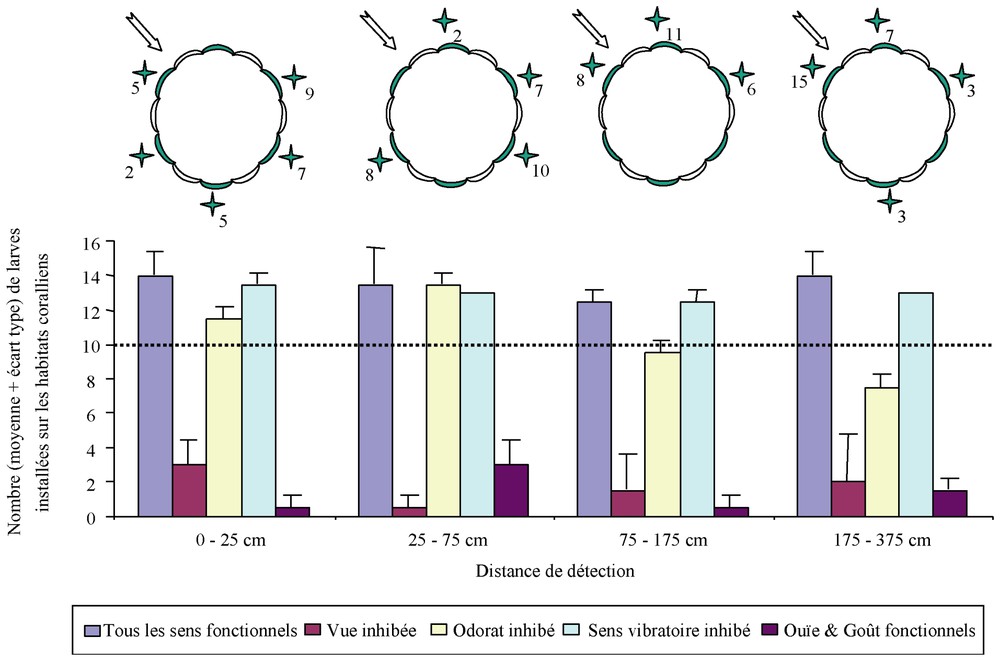
Moyenne et écart type du nombre de larves s'installant sur les arcs de cercle occupés selon la distance de détection et l'état physiologique des larves de Chromis viridis. Le trait horizontal à la valeur de 10 représente le seuil à partir duquel l'installation est significative (test du chi2 à 5%). Le choix du site d'installation est représenté par des étoiles avec le nombre total (deux réplicats) de larves installées avec tous leurs sens. La flèche indique le sens du courant.
Pour une distance inférieure à 75 cm, l'installation est significative même si l'odorat est inhibé, tandis qu'elle devient non significative si la vue est inhibée. Pour une distance supérieure à 75 cm, l'inhibition de l'odorat induit aussi une installation non significative des larves. L'absence de détection olfactive pour une distance inférieure à 75 cm proviendrait de l'inhibition de la vue par l'ajout de sparadrap. Ce moyen d'inhibition gênerait tellement les larves (obs. pers.) qu'elles perdraient leur comportement migratoire vers le signal chimique qu'elles détecteraient. Ainsi, cette étude démontre que la vue et l'odorat sont les deux sens utilisés par les larves de Chromis viridis dans la détection de leur lieu d'installation. La vue ne permet de détecter ces lieux que jusqu'à une distance de 75 cm (à des distances supérieures, la vue ne permet plus une installation significative lorsque l'odorat est inhibé), tandis que l'odorat le permet jusqu'à une distance d'au moins 375 cm (l'absence d'installation significative, lorsque la vue est inhibée, étant due à l'ajout du sparadrap). Cette conclusion est confirmée par le choix du lieu d'installation (Fig. 3). Lorsque la distance de détection est inférieure à 75 cm, les larves ne prennent pas en compte le sens du courant (répartition au hasard sur les différents arcs de cercle). En revanche, lorsque la distance est supérieure à 75 cm, elles s'installent principalement sur les arcs de cercle situés en amont du courant. Ceci suggère l'utilisation de l'odorat pour localiser la source émettrice de signaux.
4 Discussion
4.1 Limites de l'étude
Les limites de l'étude concernent essentiellement les expériences réalisées en aquarium. Je ne peux nier que des biais comportementaux doivent exister, comme notamment le stress des larves et des conspécifiques dans les aquariums. Ce stress se traduirait par l'inhibition du comportement migratoire des larves ou par la modification des signaux chimiques et mécaniques des conspécifiques. Brown et Godin [27] ont démontré que les conspécifiques soumis à un stress émettaient non plus des molécules attractives, mais des molécules répulsives. En outre, avec le protocole expérimental utilisé, un résultat négatif (pas d'attraction significative) ne permet pas de conclure que les larves n'ont pas les capacités sensorielles pour détecter leur lieu d'installation. Elles peuvent ne pas migrer, tandis qu'elles détectent les signaux émis par les conspécifiques ou l'habitat. Le fait de réaliser ces expériences en aquarium a néanmoins permis de travailler dans un mésocosme clos et contrôlable, où seul le sens ou le signal sensoriel testé est responsable de la migration des larves. À l'inverse, lors des expériences in situ, le goût et l'ouı̈e n'ont pas pu être testés individuellement et l'inhibition de la vue par l'ajout de vernis à ongles a très certainement modifié le comportement migratoire des larves. En aquarium, la diffusion du signal visuel est simplement inhibée par l'ajout d'une plaque de plastique opaque entre l'aquarium et le bac. Ainsi, ces expériences en aquarium ont permis de tester séparément la capacité de détection de cinq sens des poissons, et ceci sur 12 espèces.
4.2 Capacités sensorielles des larves dans la détection des conspécifiques
Parmi les 12 espèces étudiées dans l'expérience 1, 10 migrent vers les conspécifiques et deux ne migrent pas. L'hypothèse que les poissons aient tous leurs sens fonctionnels au stade larvaire semble être maintenant admise [7]. Pour les deux espèces qui ne migrent pas (Canthigaster janthinoptera et Pomacentrus pavo), ceci proviendrait du stress de la vie en aquarium et/ou d'une non-attraction des larves in situ. Cette dernière hypothèse signifierait que les larves, soit s'installent indifféremment sur une zone du récif [28,29], soit sélectionnent leur lieu d'installation selon les caractéristiques structurales de l'habitat [19,30], ou encore selon d'autres facteurs environnementaux, tels que les disponibilités alimentaires de l'habitat et l'absence de prédateurs ou de congénères [31,32].
Parmi les 10 espèces qui migrent, sept utilisent la vue pour reconnaı̂tre les conspécifiques. La vue est un sens bien développé chez les larves de poissons coralliens [7]. En outre, les expériences en aquarium ont été réalisées à la lumière et à courte distance. Dans l'expérience in situ, la distance de détection de la vue est inférieure à 75 cm. Néanmoins, l'estimation de cette distance est très variable selon la clarté de l'eau, la densité des massifs coralliens et la luminosité lunaire. Wottom [20] l'a estimé expérimentalement à 40 m chez les poissons (en général) et Leis et Carson-Ewart [33] l'ont estimé in situ à 10 m sur les larves de Plectopomus leopardus (Serranidae). La vue serait donc a priori le sens permettant de vérifier à courte distance que le lieu d'installation détecté est le « bon ». En effet, la densité des coraux limitant le champ de vision sur de grandes distances et la détection des lieux d'installation se faisant la nuit, la vue ne peut être le sens majeur de la détection à grande distance.
Le signal chimique des conspécifiques est détecté par sept espèces, mais l'odorat est seul responsable de cette détection. La non-utilisation de la fonction gustative pourrait être due à l'ajout de la vaseline sur les narines des larves. La vaseline aurait une odeur suffisamment forte pour empêcher les larves de détecter les conspécifiques par le goût (obs. pers.). De plus, l'odorat est souvent décrit comme un sens plus sensible que le goût, nécessitant des concentrations en phéromones élevées [34]. Cependant, Hellstrom et Doving [35] ont démontré que, selon la nature des phéromones, les poissons utilisaient l'olfaction ou la gustation dans la reconnaissance d'une proie ou d'un habitat. Ainsi, une étude est en cours pour identifier, par chromatographie liquide haute performance, la nature de ces phéromones (Lecchini, in prép.).
La perception des ondes vibratoires a déclenché la migration de cinq espèces. Ce résultat démontre, pour la première fois, que les juvéniles de certaines espèces émettent des ondes mécaniques propres à chaque espèce et que les larves peuvent les détecter. Néanmoins, cette mise en évidence a été réalisée en aquarium. Or, in situ, le sens vibratoire des larves de Chromis viridis n'a pas permis de détecter les lieux d'installation. Le rôle de ce sens dans la reconnaissance des lieux d'installation est donc à nuancer. De plus, in situ, la nature et la diffusion des ondes mécaniques seront stoppées ou fortement modifiées au contact de chaque obstacle.
L'audition n'a pas été utilisée par les larves pour détecter les conspécifiques. Or, les larves océaniques l'utilisent pour détecter l'ı̂le à coloniser, grâce au bruit des vagues sur la crête récifale et/ou au « chœur nocturne » du lagon [8,9]. Le chœur nocturne est produit par le grincement des « dents » des crevettes, des poissons et des oursins sur les coraux [36]. Pour expliquer la non-utilisation de l'audition, une hypothèse serait que les conspécifiques ne produisent pas de son dans l'aquarium. En effet, le son est un processus actif et volontaire, à la différence de l'émission des ondes vibratoires. Les larves dans l'océan détectent l'ı̂le à coloniser grâce au son car les vagues produisent en permanence du bruit sur la crête récifale et le chœur nocturne est aussi permanent. Pour que les larves reconnaissent in situ les conspécifiques par l'ouı̈e, il faudrait que ces derniers produisent du son lorsque les larves arrivent sur la crête récifale. Il y aurait alors un couplage temporel entre la production de sons par les conspécifiques et la phase de colonisation. Cette hypothèse est difficilement acceptable. En conclusion, l'ouı̈e ne joue aucun rôle dans la détection des lieux d'installation car, tout simplement, les conspécifiques ne produisent pas de son dans les aquariums (ceci aurait pu être vérifié avec des micros immergés).
4.3 Reconnaissance des lieux d'installation par la présence des conspécifiques et non par celle de l'habitat corallien
Les expériences en aquarium ont démontré que les larves reconnaissaient leur lieu d'installation par la présence de conspécifiques et non par celle de l'habitat. Ce résultat est différent de celui d'Elliott et al. [19], pour qui les larves d'Amphiprion étaient attirées par l'odeur des anémones et non par celle des conspécifiques. La présente étude n'étant réalisée que sur cinq espèces et la symbiose Amphiprion–anémone étant bien particulière, il n'est pas possible de conclure quant au rôle attracteur ou non de l'habitat corallien. Cependant, ces études soulèvent le problème du déterminisme de la reconnaissance des lieux d'installation. En effet, la détection de certains stimuli et la réaction qu'ils induisent paraissent souvent innées, mais parfois semblent être acquises au cours de la vie [1]. La question est donc de savoir si la reconnaissance des lieux d'installation est innée ou est acquise durant la phase océanique des larves.
Miyagawa [30] et Arvedlund et al. [37] ont émis l'hypothèse que certaines espèces d'Amphiprion, au stade embryon, s'imprégnaient de l'odeur des anémones hôtes. Cette hypothèse de l'imprégnation a déjà été suggérée pour le retour des juvéniles de saumons dans leur rivière de naissance [38] et pour la reconnaissance « mère–enfant » à la naissance chez le mouton [39]. Si cette hypothèse est retenue chez les poissons coralliens, cela signifierait que dans l'étude d'Elliott et al. [19], les larves ne se sont imprégnées que de l'odeur des anémones et non de celle des parents vivant sur ces anémones. Dans la présente étude, cela signifierait que les larves (des espèces à œufs démersaux) ne se sont imprégnées que de l'odeur des conspécifiques. Cependant, les larves détectent aussi les conspécifiques par la vue et la perception des ondes vibratoires. Il y a donc obligatoirement un apprentissage de la reconnaissance des conspécifiques lors de la phase océanique. Durant cette phase, les larves vivent généralement en groupes [40], ce qui permet d'apprendre à reconnaı̂tre les individus de la même espèce puisqu'ils ont la même forme, les mêmes capacités natatoires, les mêmes exigences alimentaires, etc. Cet apprentissage facilitera ensuite la reconnaissance visuelle, olfactive et mécanique des conspécifiques lors de la phase d'installation.
5 Conclusion
Ces expériences sur les capacités sensorielles des larves à l'installation ont démontré que les poissons coralliens choisissaient leur lieu d'installation grâce à la présence de conspécifiques et non à cause des caractéristiques de l'habitat. Parmi les différents sens fonctionnels des larves, seuls la vue, l'odorat et la perception des ondes vibratoires sont utilisés dans la reconnaissance des conspécifiques. Le déterminisme de l'utilisation de ces sens proviendrait d'une combinaison de différents processus : imprégnation de l'odeur du lieu parental à la naissance, apprentissage du conspécifique dans le milieu pélagique et degré de développement des organes sensoriels de chaque espèce. Cependant, les conditions environnementales dans lesquelles se retrouvent les larves après la colonisation du récif sont différentes de celles des aquariums. In situ, les larves doivent non seulement détecter les signaux émis par les lieux d'installation, mais aussi ceux émis par les prédateurs et les congénères. Il est donc évidant que les études en laboratoire, si elles sont nécessaires au début, devront être remplacées dans l'avenir par des études in situ afin de prendre en compte l'ensemble des informations que reçoivent les larves au moment de choisir un lieu d'installation.
Remerciements
L'auteur remercie James Algret et Yannick Chancerelle pour leur participation aux expériences à Moorea et tous ceux qui ont contribué à améliorer la qualité du manuscrit (René Galzin, Pascale Panunzzio, Serge Planes et Jean-Pierre Pointier). L'auteur remercie aussi Vincent Dufour, directeur de l'entreprise Aqua-Fish Technology, qui a fourni les filets de crête permettant la capture des larves. Enfin, l'auteur remercie la fondation d'entreprise Total pour son aide financière dans la construction des aquariums.