

Abridged English version
The night monkeys, genus Aotus, are the only nocturnal monkeys among neotropical primates. The taxonomy of this genus is unclear. Considered as monospecific until 1983, the genus is now divided into two groups, named ‘grey neck’ and ‘red neck’, respectively. These groups differ in the pelage, in morphology, in distribution (found north and south of the Amazon, respectively), but also chromosomally. Nine species are nowadays recognized, but the taxonomic history of Aotus is not yet fully understood.
Nights monkeys are WHO-recommended primates for the study of malaria, first because they are the only cebids susceptible to infection, but also because this susceptibility is variable between species, probably due to karyotypic variations. Thus, they are intensively studied as a research model for human malaria Plasmodium falciparum and Plasmodium vivax. A good knowledge of the Aotus taxonomy is therefore necessary.
Taxa are usually defined using genetic and phenotypic data too. The differentiation of species mainly occurred in night monkeys by genetic drift and separation of species via rivers, which play the role of geographical barriers.
Two populations of Kuhl's night monkeys (Aotus infulatus), one from each bank of the Tocantins River in Brazilian Amazonia, show remarkable genetic differences. The aim of this paper is to test the phenotypic difference of these populations by studying the shape of the skull. Some taxonomic conclusions could ensue from these morphological results.
The cranial morphology of 28 specimens of Aotus from the Amazonian Basin, including specimens of Aotus infulatus from both banks of the Rio Tocantins, was studied using geometrical morphometrics in three dimensions. A Procrustes fitting was performed and Principal-Component Analysis and Discriminant Analysis were computed. Statistics attest to a significant distinction in the cranial morphology of the two ‘gray-neck’ and ‘red-neck’ groups. This distinction is significantly correlated with cranial size differences, as described in previous studies. Both populations of Khul's monkeys show notable cranial differences in the shape of the neurocranium, in the zygomatic arches, and in the palate, but also in the relative size of the orbits. Although these differences are not statistically significant, they are true differences and should be taken into account for further studies. However, we cannot suggest a taxonomic differentiation of these populations (for example, into two sub-species). As morphometric and genetic results are highly concordant, they could indicate that this species is undergoing recent rapid evolutive changes.
A parasitological study attests that, contrary to what was supposed, Aotus infulatus is susceptible to malaria. As mentioned previously, susceptibility to malaria parasites is variable between taxa. The specimens analyzed in this study were from the banks of Rio Tocantins, but with no detailed localization. As morphometric and genetic results suggest an ongoing differentiation of the populations from each bank of the river, a difference in susceptibility to malaria between these populations is suspected, which could be confirmed by further studies.
1 Introduction
Les douroucoulis sont les seuls primates nocturnes du Nouveau Monde. Ces singes sont étudiés pour leur système visuel (par exemple, [1,2]), leur écologie [3–5], leur reproduction [6], mais aussi pour leur capacité à résister à la malaria Plasmodium falciparum, capacité variable selon les taxons [7]. Une application médicale à l'homme nécessite ainsi de bien définir la taxinomie et de bien comprendre l'histoire évolutive de ce genre.
Le genre Aotus, défini en 1811 par Illiger, a longtemps été considéré comme monospécifique. Ainsi, des différences locales suivant différents morphoclines ont été reconnues au sein de l'espèce unique Aotus trivirgatus [8,9], et ce n'est qu'à partir de 1983 [10], avec la définition de neuf espèces valides, dont deux composées de deux sous-espèces, que le genre devient polyspécifique. Hershkovitz [10] combine des données de couleur du pelage (notamment du cou), du caryotype, de la morphologie crânienne et postcrânienne et de la biogéographie pour distinguer deux grands groupes d'espèces : le groupe des « cou gris » est composé des taxons Aotus lemurinus (A. l. lemurinus et A. l. griseimembra), A. brumbacki, A. trivirgatus et A. vociferans, répartis au nord des rios Amazone, Solimões et Marañón. Le second groupe, des « cou rouge », est composé des espèces Aotus miconax, A. nancymaae, A. nigriceps, A. azarae (A. a. azarae et A. a. boliviensis) et A. infulatus, répartis au sud des cours d'eau nommés ci-dessus. Hershkovitz [10], puis d'autres auteurs (par exemple, [7,11–14]) s'intéressent à la susceptibilité de ces espèces à la malaria. Le groupe des « cou gris » est sensible au parasite, alors que celui des « cou rouge » est décrit comme plus résistant.
Hernández-Camacho et Cooper [15] précisent la taxonomie en définissant en 1976 la nouvelle sous-espèce Aotus lemurinus zonalis ; Ramirez-Cerquera fait de même en 1983 [16] avec l'espèce Aotus hershkovitzi, membre des « cou gris » et proche de l'espèce Aotus lemurinus [17]. Ford [11] analyse les données biométriques des crânes et des dents, mais aussi les données de coloration de pelage, à l'aide de méthodes multivariées dans le but de réactualiser la taxinomie du genre. Au regard de ses résultats issus d'analyses en composantes principales, il décide de simplifier la taxinomie en regroupant les espèces Aotus vociferans, Aotus lemurinus et Aotus brumbacki et en incluant Aotus azarae boliviensis au sein de l'espèce Aotus infulatus. Il propose également qu'Aotus brumbacki soit une sous-espèce de l'espèce Aotus lemurinus (ne pouvant pas distinguer les variations de couleur du pelage). Finalement, Groves, en 2001 [18] propose une nouvelle révision de la taxinomie et distingue l'espèce Aotus lemurinus d'Aotus vociferans par la couleur du pelage et par le nombre de chromosomes. Celui-ci conserve ces deux taxons au rang d'espèces et considère également qu'Aotus infulatus, A. azarae et A. boliviensis forment un complexe, dont de nombreux caractères sont communs, et en fait trois sous-espèces de l'espèce Aotus azarae. Ainsi, il apparaît que la taxinomie de ce genre est récente et reste discutée.
L'histoire évolutive du groupe est également très informative, puisque plusieurs auteurs [17–19] mentionnent que, chez les douroucoulis, les fleuves auraient constitué des barrières géographiques importantes, qui auraient séparé dans un premier temps les populations, puis les taxons. Ainsi, les douroucoulis se seraient différenciés essentiellement par dérive génétique (dispersion/isolation). La radiation serait originaire de Colombie ou du Pérou (au sein de l'aire de répartition d'Aotus vociferans) et la radiation se serait faite successivement de part et d'autre des rios Amazone/Marañon, Negro, Orénoque, Trombetas, Tapajós/Madeira/Madre de Dios.
Schneider – [20], puis Sampaio – [21], étudient les protéines du sang et l'enzyme anhydrase carbonique chez des individus de l'espèce Aotus infulatus provenant des rives droite et gauche du rio Tocantins. Les différences entre ces deux populations sont remarquables et de telles différenciations moléculaires ont une importance capitale dans le contexte évolutif évoqué précédemment.
Si des différences morphologiques ont été observées et décrites entre les différents taxons du genre, une étude morphologique n'a jamais été menée sur les individus de l'espèce Aotus infulatus des rives droite et gauche du rio Tocantins. Connaissant les différences génétiques décrites par Schneider – et Sampaio –, on peut se poser la question d'une différenciation morphologique entre ces populations et des conséquences taxinomiques qu'elle engendrerait.
La forme du crâne est très informative. Elle résulte de l'embryogenèse, reflète diverses adaptations du régime alimentaire et de la locomotion. Elle est également le témoin du dimorphisme sexuel secondaire chez les espèces dimorphiques.
Les méthodes de morphométrie géométrique permettent la modélisation d'objets biologiques (dans leur ensemble) en deux, ou trois dimensions, mais aussi la quantification des différences de conformation entre ces objets. De manière générale, les résultats de ce type de méthodes sont représentés à l'aide de diagrammes spatiaux construits à l'aval de méthodes d'analyses multivariées. Ces méthodes de morphométrie géométrique sont souvent appliquées en biogéosciences afin de comparer la conformation d'organismes, aux divers niveaux taxinomiques (études inter- ou intraspécifiques). Elles constituent également un outil très pertinent dans l'étude de la différenciation morphologique en rapport avec la différenciation géographique [22–26]. Ces méthodes s'avèrent donc tout à fait appropriées pour mener une étude morphologique crânienne à un niveau interpopulationnel.
2 Matériel et méthodes
2.1 Matériel
L'échantillon est composé de 28 crânes de spécimens appartenant à quatre taxons du genre Aotus (Tableau 1). Ces spécimens ont été étudiés au Museu Paraense Emílio Goeldi (Belém, Pará, Brésil) et au Muséum national d'histoire naturelle (Paris, France) et proviennent de différentes localités du Brésil et d'Équateur (Fig. 1). Cet échantillon peut sembler de petite taille, mais le nombre de spécimens disponible pour le taxon Aotus infulatus est faible, et ce particulièrement lorsque l'on travaille sur le crâne (voir les échantillons analysés par Thorington et Vorek en 1976 et par Hershkovitz en 1983). Ainsi, à titre d'échange, l'American Museum of Natural History de New York et le National Museum of Natural History de Washington ont été visités afin d'augmenter le nombre de spécimens, mais aucun crâne d'Aotus infulatus n'est disponible dans ces collections. D'autre part, le but de ce travail étant d'étudier la morphologie crânienne des populations des rives droite et gauche du rio Tocantins, seuls quelques taxons des deux grands groupes de douroucoulis ( « cou gris » et « cou rouge ») sont analysés. Ils permettent de replacer les individus de l'espèce Aotus infulatus dans le cadre morphologique crânien du genre. Les spécimens des deux populations de l'espèce Aotus infulatus provenant des rives droite et gauche du rio Tocantins ont été principalement collectés entre 1985 et 1986 dans la région de Tucuruí lors de la construction d'un barrage hydroélectrique (Fig. 2).
Composition de l'échantillon étudié
| Taxons étudiés | Mâles | Femelles | Total |
| Groupe des « cou gris » | |||
| Aotus vociferans | 3 | 1 | 4 |
| Aotus trivirgatus | 7 | 4 | 11 |
| Groupe des « cou rouge » | |||
| Aotus nigriceps | 1 | 2 | 3 |
| Aotus infulatus (rive gauche du rio Tocantins) | 2 | 3 | 5 |
| Aotus infulatus (rive droite du rio Tocantins) | 3 | 2 | 5 |
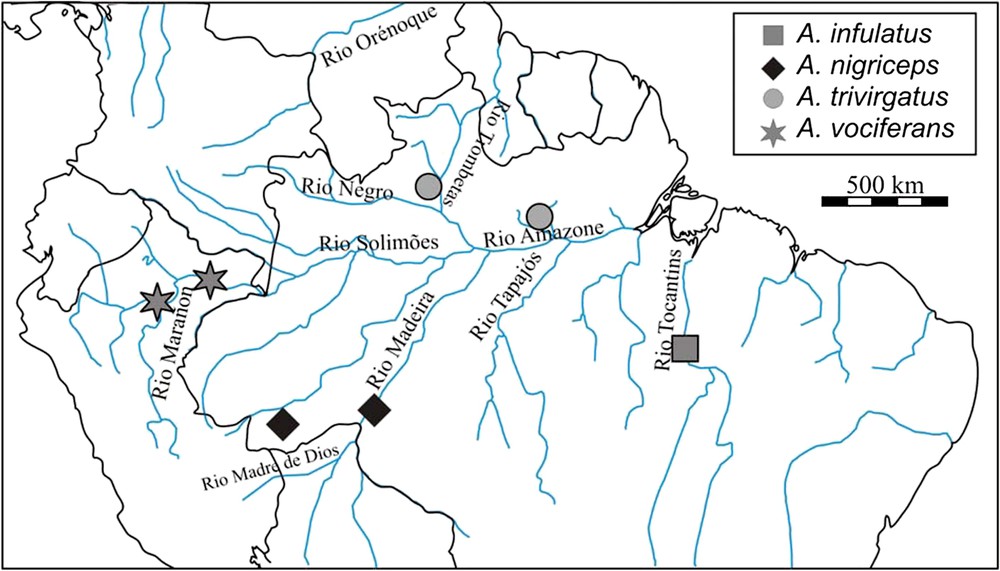
Localisation géographique de la provenance des spécimens qui composent l'échantillon. Les douroucoulis appartenant au groupe des « cou gris » sont répartis au nord des rios Amazone/Solimões/Marañon, alors que ceux du groupe des « cou rouge » sont répartis au sud. Les différents fleuves indiqués jouent ou ont joué un rôle actif dans la distinction des espèces par dispersion/isolation au cours de l'histoire évolutive du genre.
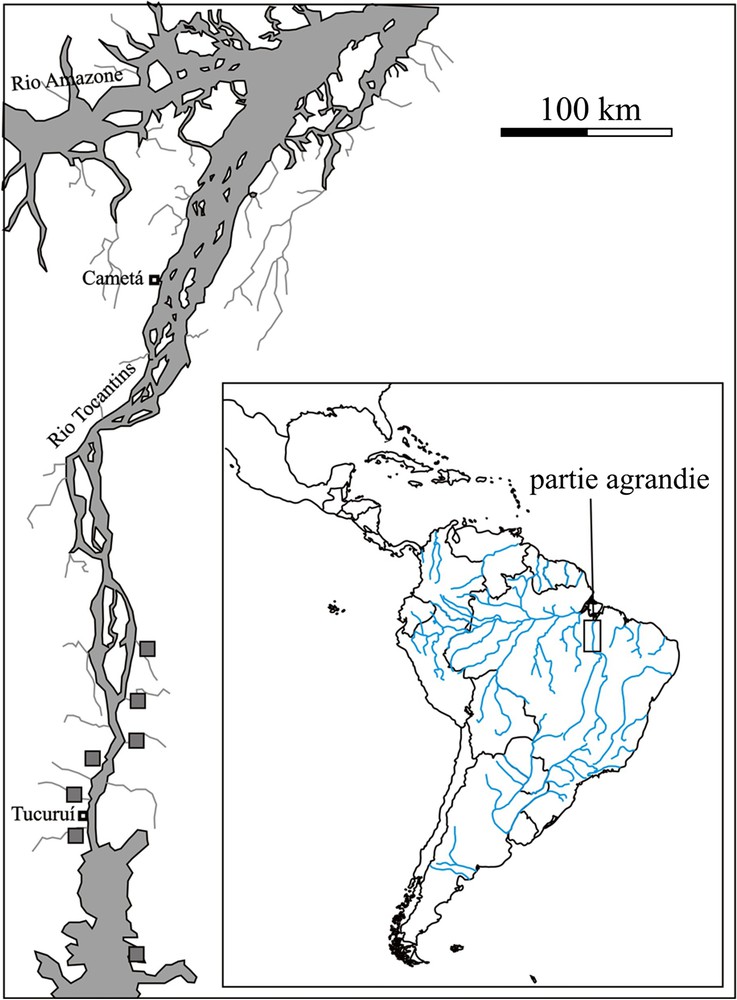
Localisation géographique de la provenance des spécimens d'Aotus infulatus des populations des rives droite et gauche du rio Tocantins.
2.2 Méthodes
L'essor des méthodes de morphométrie date des trois dernières décennies. Même si les travaux de D'Arcy Thompson [27] en ont posé les bases au début du XXe siècle, ces méthodes ont pris un essor considérable avec le développement des outils de traitement statistique et de calcul informatique. Ainsi, l'évolution de l'aptitude des microordinateurs a été accompagnée de nombreuses réflexions méthodologiques et applications (par exemple, [28–39]). Elles visent à modéliser la forme des objets naturels et à mesurer leurs différences. Une nouvelle étape correspond à la définition conceptuelle de la « conformation » et de la « taille », ainsi que de leur distinction [40], qui a mené à la fameuse équation :
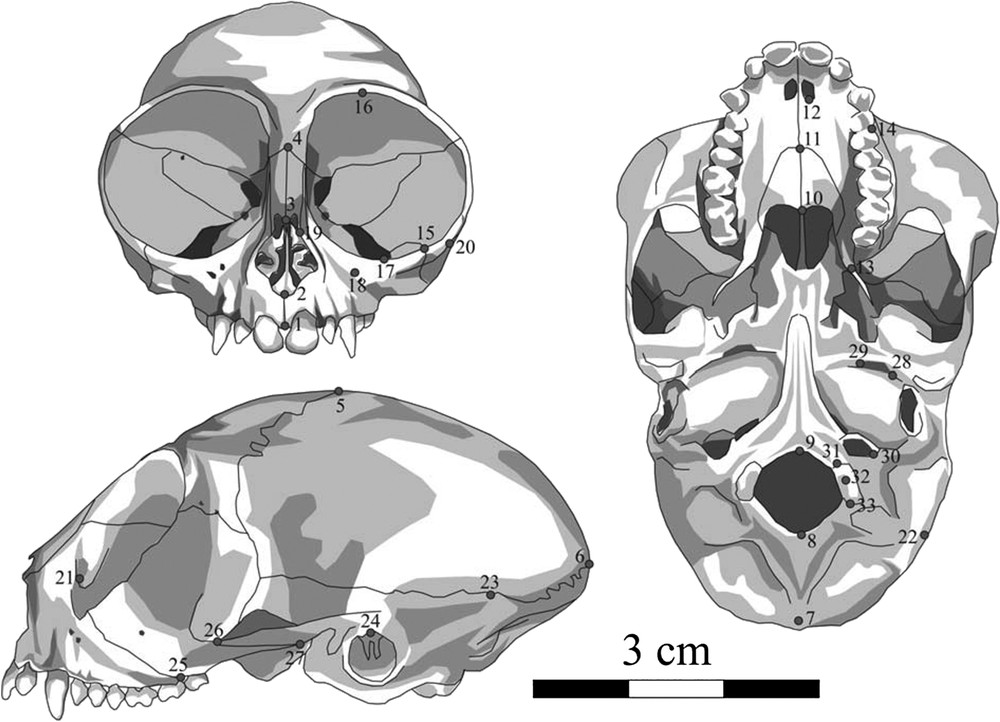
Identification des 33 points homologues définis sur le crâne et utilisés pour l'étude morphométrique. Par convention, les points sont pris sur le plan sagittal et sur le côté gauche du crâne. Ceci permet d'éviter l'effet d'une éventuelle asymétrie fluctuante.
Les méthodes Procrustes avec ajustement généralisé aux moindres carrés ont été utilisées pour analyser les données [29,34,42,43]. Après avoir testé leur normalité par un test de Kolgomorov–Smirnov, les résidus Procrustes ont été utilisés comme variables dans une analyse multivariée (ACP) ; les deux axes principaux, qui résument respectivement 82,1 et 16,2% de la variance totale, ont été retenus afin de construire l'espace morphologique (Fig. 4).
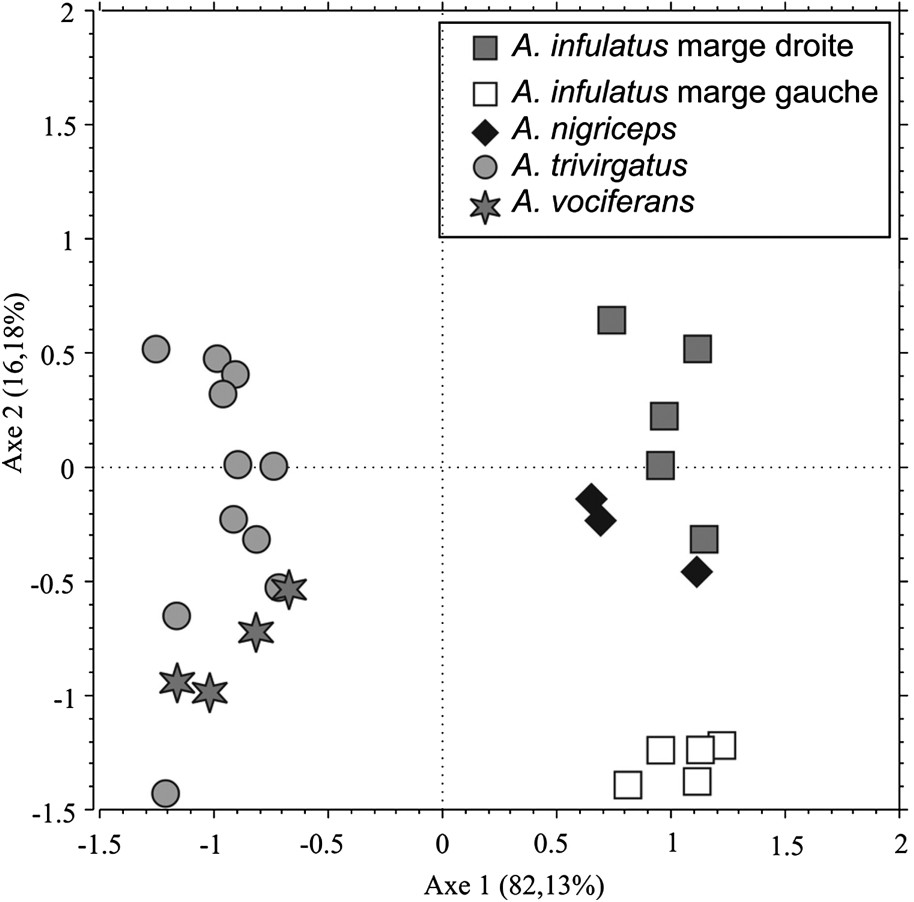
Présentation du premier plan principal Axe 1/Axe 2, qui résume 98,31% de la variance totale. L'Axe 1 est significativement corrélé à la taille des spécimens (taille mesurée du crâne). Sur l'Axe 1 se différencient les morphologies crâniennes du groupe des « cou gris » et des « cou rouge ». L'Axe 2 n'est pas significativement corrélé à la taille. Sur ce second axe, deux sous-groupes morphologiques se distinguent, décrivant la différenciation de la morphologie crânienne des deux populations de l'espèce Aotus infulatus de part et d'autre du rio Tocantins.
Il est à noter que le rapport entre le nombre de mâles et de femelles n'est pas identique pour chaque taxon de l'échantillon. Le dimorphisme sexuel est peu perceptible chez les douroucoulis (par exemple, [11,18,19]). L'influence du dimorphisme sexuel a été testée par une analyse discriminante sur les scores des deux premiers axes principaux. La distance de Mahalanobis () entre mâles et femelles, ainsi que les valeurs F du test de Fisher et les valeurs des probabilités associées, ont été calculées pour chaque taxon. Les mâles et les femelles ne présentent pas de différences morphologiques significatives et le dimorphisme sexuel intervient peu dans la distinction de la morphologie crânienne des spécimens. La proportion de mâles et de femelles composant l'échantillon des taxons peut donc ne pas être strictement identique, car le dimorphisme sexuel n'influence pas significativement la morphologie crânienne. Les analyses effectuées sur notre échantillon ne seront donc pas biaisées par cette différence de proportion.
Afin de tester statistiquement la significativité des différences morphologiques entre les taxons, une analyse discriminante a été réalisée sur les scores de l'ACP. Les distances de Mahalanobis, ainsi que les valeurs des tests F et les probabilités associées, ont été calculées et sont présentées dans le Tableau 2.
Valeurs des coefficients F de Fisher (demi-matrice supérieure) et des probabilités p associées (demi-matrice inférieure), calculées sur les distances de Mahalanobis () après analyse discriminante sur les scores des deux axes principaux de l'analyse en composantes principales. Les valeurs significatives au seuil de 5% sont indiquées en gras
| Aotus infulatus | Aotus infulatus | Aotus nigriceps | Aotus trivirgatus | Aotus vociferans | |
| (rive droite) | (rive gauche) | ||||
| Aotus infulatus (rive droite) | 2,13 | 0,23 | 17, 46 | 13, 85 | |
| Aotus infulatus (rive gauche) | 0,127305 | 1,40 | 19, 55 | 14, 22 | |
| Aotus nigriceps | 0,875376 | 0,271293 | 9, 66 | 8, 72 | |
| Aotus trivirgatus | 0,000006 | 0,000003 | 0,000331 | 0,31 | |
| Aotus vociferans | 0,000033 | 0,000028 | 0,000594 | 0,818775 |
3 Résultats
Les spécimens sont projetés dans le plan principal Axe 1/Axe 2 (Fig. 4). Deux groupes morphologiques se distinguent sur l'Axe 1. Le premier, dont la morphologie est décrite par des valeurs comprises entre −1,5 et −0,5, correspond aux spécimens des taxons Aotus trivirgatus et Aotus vociferans. Un second groupe, dont la morphologie est décrite par des valeurs comprises entre 0,5 et 1,5, correspond aux spécimens des taxons Aotus nigriceps et Aotus infulatus. Ces deux ensembles morphologiques correspondent aux groupes des « cou gris » (pour le premier) et des « cou rouge » (pour le second) décris par Hershkovitz. Il est à noter que la distinction morphologique de ces deux groupes est observable sur l'Axe 1 et que cet axe est significativement corrélé à la taille (, ), les spécimens du groupe des « cou gris » étant en moyenne plus petits que ceux du groupe des « cou rouge ». Il existe une relation allométrique entre ces deux groupes, mais, puisque la valeur du coefficient de corrélation est faible, elle n'explique pas entièrement, même si cette allométrie est significative, la distinction morphologique des groupes.
La contribution des variables sur chacun des axes permet d'identifier les différences morphologiques entre ces deux groupes. Ainsi, ces différences concernent essentiellement la taille des orbites (relativement plus grandes chez les petites espèces que chez les grandes). Cette dissemblance peut néanmoins n'être qu'un effet de la comparaison de conformations lors de la description de structures dont la taille varie peu, alors que la taille générale du crâne augmente. De plus, les arcades zygomatiques des individus du groupe des « cou gris » sont plus fines que celles des individus du groupe des « cou rouge ». Enfin, la disposition des points de la base du crâne et du palais est différente, traduisant un palais plus haut chez les individus « cou gris » que chez les « cou rouge ».
Qu'en est-il, à présent, de la disparité (différence morphologique) au sein même des groupes ? Les spécimens des taxons Aotus vociferans et Aotus trivirgatus, qui composent le groupe des « cou gris », présentent des morphologies proches, puisque les zones qu'occupent ces groupes au sein de l'espace morphologique se chevauchent. La seule utilisation de l'espace morphologique ne permet pas une distinction des deux taxons. Le test de Fisher calculé entre ces deux taxons indique que les différences morphologiques ne sont pas significatives au seuil de 5%.
Le groupe des « cou rouge » est composé des taxons Aotus infulatus et Aotus nigriceps. Des sous-groupes morphologiques se distinguent au sein de ce groupe ; cette distinction s'effectue sur l'Axe 2 (qui n'est pas significativement corrélé à la taille). D'une part, les spécimens de la population de la rive gauche du rio Tocantins de l'espèce Aotus infulatus se regroupent autour des valeurs 0,7 à 1,3 sur l'Axe 1 et −1,5 à −1,2 sur l'Axe 2. D'autre part, les spécimens de la population de la rive droite se différencient clairement de ceux de la rive gauche, puisqu'ils se repartissent entre les valeurs −0,4 et 0,7 sur l'Axe 2. La part de l'espace morphologique qu'occupe chacune des populations de l'espèce Aotus infulatus est distincte, traduisant des différences de morphologie crânienne décrites par l'Axe 2. Ainsi, au regard des contributions des variables sur cet axe, le neurocrâne est plus développé antéro-postérieurement chez les individus provenant de la rive gauche du fleuve que chez ceux de la rive droite. De même, les individus de la rive droite possèdent des arcades zygomatiques plus fortes et plus hautes que ceux de la rive gauche. Enfin, il existe aussi de faibles différences concernant la forme des orbites ainsi que la position de l'ouverture nasale. Il est également à noter que les spécimens de la population provenant de la rive gauche du fleuve présentent une morphologie crânienne assez proche (la dispersion du nuage de points de ce sous-groupe morphologique au sein de l'espace morphologique est faible). Il semble que la variabilité interindividuelle de la morphologie crânienne soit plus forte chez les spécimens provenant de la rive droite du fleuve. Il convient d'être prudent quant à l'interprétation de ce dernier résultat, qui est fortement dépendant de l'échantillonnage.
Les spécimens de l'espèce Aotus nigriceps présentent des morphologies crâniennes décrites par les valeurs comprises entre −0,1 et −0,5 sur l'Axe 2. La part de l'espace morphologique qu'occupe ce taxon recoupe celle qu'occupent les spécimens de la population de la rive droite du rio Tocantins de l'espèce Aotus infulatus. Les morphologies crâniennes sont proches entre ces deux groupes. Les valeurs de F et les probabilités associées, calculées à partir des distances de Mahalanobis () sont présentées dans le Tableau 2. Il apparaît que les populations de l'espèce Aotus infulatus des rives droite et gauche du rio Tocantins ne sont pas statistiquement différentes. Ainsi, si une réelle différenciation existe (elle est mise en avant par l'occupation de l'espace morphologique), cette différenciation n'est pas significative. La morphologie crânienne des spécimens de ces populations ne peut donc être considérée, au regard de nos résultats, comme significativement distincte. Il faut cependant noter que la valeur de F pour ces deux populations est la plus élevée des valeurs non significatives, inversement aux probabilités associées.
Les tests statistiques ne permettent pas non plus de distinguer morphologiquement les spécimens de l'espèce Aotus nigriceps de ceux de l'espèce Aotus infulatus. Il est intéressant de noter que les spécimens de l'espèce Aotus infulatus provenant de la rive droite du rio Tocantins sont plus proches morphologiquement de ceux de l'espèce Aotus nigriceps que ceux provenant de la rive gauche.
Ces conclusions sont très positives en ce qui concerne le potentiel discriminatoire de la méthode globale utilisée pour cette étude. En effet, elle permet de mettre en avant des différences morphologiques très fines, apportant ainsi de nouveaux arguments morphologiques et proposant également une visualisation des résultats. Il est à noter que les tests statistiques utilisés permettent de séparer significativement les morphologies crâniennes des deux grands groupes de douroucoulis ( « cou gris » et « cou rouge »), mais qu'au sein de ces groupes la distinction morphologique n'est pas significative.
4 Discussion
Les résultats morphométriques de notre étude confirment l'existence de deux groupes morphologiques correspondant aux deux groupes taxinomiques d'Hershkovitz. Ainsi, les « cou gris » et les « cou rouge » possèdent une morphologie crânienne statistiquement différente. Ces différences entre les deux groupes morphologiques ont été identifiées, lors de notre étude, comme les résultats de variation des conformations liées à la taille (allométrie), en accord avec ce qui est proposé par les précédents auteurs. Ford [11] propose un schéma évolutif tiré des conclusions d'Hershkovitz [10], mais aussi de son analyse multivariée des données craniodentaires et des couleur de pelage, complétant les connaissances chromosomiques et de l'étude des protéines du sang. Dans ce schéma, le groupe des « cou rouge » dérive de celui des « cou gris », et les différences morphologiques entre les taxons concernent essentiellement l'augmentation de la taille du crâne chez Aotus nancymaae et Aotus infulatus, mais également l'augmentation en taille de l'appareil masticateur chez Aotus trivirgatus et Aotus infulatus. Les conclusions morphométriques de cette étude confirment et complètent ces résultats ; il semble que les individus du groupe des « cou rouge » présentent un crâne plus grand et plus robuste (augmentation en hauteur des arcades zygomatiques) que celui des « cou gris ».
La différenciation des populations des rives droite et gauche du rio Tocantins au sein de l'espèce Aotus infulatus a été proposée par Schneider – et Sampaio – [20,21] au regard des fréquences alléliques (CA2*9) et de la variance interlocus (dix loci d'enzymes des érythrocytes et deux loci des protéines du sérum). Cependant, ces différences restent faibles et ne sont pas statistiquement significatives. Ceci indiquerait une séparation récente, voire en cours, de ces populations. Les résultats de notre étude morphométrique viennent confirmer les conclusions génétiques de Schneider – et de Sampaio – Ainsi, il existe des différences entre les morphologies crâniennes de ces deux populations, qui concernent essentiellement la forme du neurocrâne et le développement en hauteur des arcades zygomatiques, ne sont pas significativement corrélées à la taille et ne décrivent pas la séparation morphologique des dimorphes. Dans le cas de ces deux populations de l'espèce Aotus infulatus, la distinction génétique, tout comme la distinction morphologique, n'est pas statistiquement significative. Ainsi, même si de réelles différences ont été observées, décrites et quantifiées, elles ne permettent pas de justifier une séparation taxinomique de ces populations.
Lors d'études cytogénétiques, Pieczarka – [44–46] insistent sur les faibles différences entre les espèces du groupe des « cou rouge ». Ainsi, A. nancymaae et A. azarae présentent le même degré de similarité que deux populations de la même espèce (A. infulatus provenant des rives droite et gauche du rio Tocantins). De même, ces auteurs considèrent que les différences entre Aotus infulatus et Aotus azarae boliviensis ne sont pas suffisamment importantes pour qu'on puisse envisager une spéciation. Comment interpréter cette conclusion maintenant qu'une distinction génétique, mais également morphologique, de ces deux populations a été mise en avant ? Les concepts de populations, d'espèces et de sous-espèces sont remis en cause. Colin Groves [47] se penche sur le problème de la définition de ces entités taxinomiques. Cet auteur rappelle qu'une sous-espèce correspond à un fragment d'espèce, c'est-à-dire à une population géographiquement distincte d'une autre population de l'espèce. Les apports de la génétique et de la phylogéographie sont très importants dans la reconnaissance de taxons et certains concepts d'espèces sont basés sur ces données (par exemple, le principe de concordance proposé par Avise [48]). Cependant, Groves insiste sur le fait que ces données génétiques ne permettent pas à elles seules de conclure quant à la taxinomie et qu'il est absolument nécessaire d'avoir recours aux observations phénotypiques.
La difficulté pour identifier les espèces de ce genre ne semble pas seulement due à des problèmes de définition et de taxinomie. Comme suggéré précédemment, certains groupes de douroucoulis sont probablement sujets à de récents et rapides changements évolutifs impliquant une grande variabilité intraspécifique et une faible variabilité interspécifique. Ceci induit une forte similarité entre les espèces (qui a été mise en avant par notre analyse morphométrique et des tests statistiques non significatifs au sein de chacun des deux grands groupes de douroucoulis), qui implique une complexification de la définition des taxons. Il apparaît qu'à ce jour la taxinomie de ce genre n'est pas encore clairement résolue.
Peu de travaux ont été publiés concernant la différenciation de sous-espèces chez les singes du Nouveau Monde. Les travaux d'Hershkovitz [49] de Cheverud et Moore [50], de Kohn – [51] et de Peres – [52] sur les espèces et sous-espèces de tamarins peuvent néanmoins être utilisés à titre de comparaison. Chez les tamarins, les spéciations se sont faites majoritairement de façon non adaptative, avec une forte contribution de la dérive génétique. Les fleuves auraient isolés les populations et les différences morphologiques entre les taxons résulteraient principalement de différences génétiques plutôt que d'une adaptation à un environnement précis. Ayres [53] remarque que de nombreux primates néotropicaux sont limités, dans leur distribution, par des fleuves et les différenciations ; les spéciations se font la plupart du temps le long d'une rive. Ceci est d'autant plus vrai lorsque la zone de distribution est éloignée de la source du fleuve, c'est-à-dire que la largeur du fleuve augmente [52]. Peu de choses sont connues quant à la capacité d'Aotus infulatus à franchir ce type de barrière géographique ; Schneider – [20] considèrent qu'au regard de l'hétérogénéité génétique (confirmée par nos résultats morphométriques) entre les deux populations et compte tenu de la largeur du rio Tocantins au niveau du barrage de Tucuruí (1300 m de largeur), le fleuve se comporte comme une barrière géographique effective. Ceci est d'autant plus valable que cette zone a subi une déforestation intense liée à l'élevage du bétail, depuis plus d'un siècle. Cette déforestation a augmenté la zone à franchir pour que les douroucoulis puissent passer d'une rive à l'autre, ce qui, selon Peres –, constitue une barrière bien plus infranchissable que le fleuve lui-même [52].
Les douroucoulis sont de bons sujets pour les recherches sur la malaria et plusieurs espèces résistent aux infections par les parasites humains Plasmodium falciparum et Plasmodium vivax. Cette susceptibilité à la malaria est variable selon les taxons et probablement selon le caryotype [12]. Aotus azarae boliviensis est connu pour être très sujet à la malaria, alors qu'Aotus infulatus semble particulièrement résistant. Cependant, en 2000, Carvalho – [13] testent la susceptibilité à la malaria de cinq spécimens de l'espèce Aotus infulatus. Ces auteurs concluent que cette espèce n'est pas particulièrement résistante aux parasites et qu'elle constitue un excellent modèle pour la recherche médicale. Ce résultat est en lien direct avec ceux de notre étude. En effet, les spécimens étudiés sont tirés d'un échantillon ayant été collecté en 1984 lors de l'inondation de la zone amont au barrage de Tucuruí, mais ces spécimens proviennent à la fois des rives droite et gauche du rio Tocantins. La provenance exacte des cinq individus étudiés n'est pas précisée, mais on peut alors se poser la question d'une éventuelle différence de résistance aux parasites de la malaria entre les populations des deux rives. Des études complémentaires sont indispensables afin d'éclaircir ce point ; il apparaît que les résultats de notre étude pourraient s'avérer très informatifs dans le cadre de futures applications médicales.
Remerciements
Cet article constitue une contribution de l'UMR CNRS 5561 « Biogéosciences », Dijon (équipe « Forme, évolution, diversité ») et du laboratoire de l'École pratique des hautes études « Paléobiodiversité et Préhistoire », Dijon. L'auteur remercie le musée Paraense Emílio Goeldi (Belém, Brésil), et en particulier le Dr José de Souza e Silva Júnior pour l'accès aux collections, le Dr Maria Paula Cruz Schneider (université fédérale du Pará, Belém, Brésil) pour son aide et son soutient lors du travail à Belém, ainsi que le Dr Marc Godinot (EPHE, Paris) et le Pr. Gabriel Marroig (université de São Paulo, São Paulo, Brésil) pour les conseils qu'il lui a donnés. L'auteur remercie également l'Institut de paléontologie humaine de Paris pour le prêt du microscribe et Carmela Château Smith pour la relecture de la version abrégée en anglais.