

1 Introduction
Dipnoi, or lungfishes, is a clade of Sarcopterygian fishes represented by a large number of fossil and six living species [1]. Dipnoans are recognized as a distinct group since the description of the fossil genus Ceratodus by Agassiz [2]. It was not until the beginning of the 19th century that the first living representatives of the dipnoan clade were discovered. The description of the South American Lungfish Lepidosiren paradoxa Fitzinger, 1837 and other living species, including the African Protopterus annectens (Owen, 1839) [3], triggered an intense debate on the nature of these animals. They were initially regarded as amphibians (or even closely related to reptiles) by some authors, while others considered them as fishes (see [4,5]). By the end of the 19th century their placement among fishes was widely accepted. Today, the Dipnoi are generally regarded as the sister group of tetrapods (e.g., [6]), but some studies conclude that the relationships among lungfishes, coelacanths, and tetrapods remain unresolved (e.g., [7]).
L. paradoxa was the first living dipnoan species to be discovered. It was described from material collected by Johann Natterer. During his expedition in Brazil (1817–1835) [8], he collected only two lungfish specimens [9–11]. Along with a large number of specimens of other groups, they were sent to the k.k. Naturalien-Cabinet in Vienna, Austria. One lungfish specimen had been collected in a creek close to the town of Borba (at that time in the Grão-Pará Province and now the state of Amazonas, Brazil) in the Madeira River drainage. The second specimen was found in a swamp close to “Vila-Nova”, on the left bank of the Amazon River (also in Amazonas) [10,11]. According to Natterer, this species was rare and unknown to most natives in these regions, as it is still the case today.
The specimens were briefly described by Leopold Fitzinger [9], who erected the name L. paradoxa, but did not include a detailed anatomical description. Possibly also as early as 1837, but after Fitzinger, Natterer published a description of the morphology, including a few details of the internal organs, which is more detailed than Fitzinger's description. The exact publication date of Natterer's article (1840 in the title page) is a matter of ongoing discussions, which we do not intend to enter here, because Natterer's is not the name-establishing article. A complete description was provided a few years later by Theodor Bischoff [11], based on the same specimens. Bischoff thoroughly described the skeleton, muscles, and nervous, respiratory, circulatory, digestive, reproductive, and urinary systems. The cross references in these three publications dealing with L. paradoxa leave no doubt that Natterer (probably in 1837) published after Fitzinger [9] and before Bischoff [10]. Bischoff [10] clearly stated that his description was based on the two specimens collected by Natterer and described by Fitzinger [9], and that these are the syntype of L. paradoxa. Unfortunately, none of the three authors involved gave any curatorial information such as collection numbers for the specimens. It is clear, however, that those specimens were kept in the k.k. Naturalien-Cabinet, which preceded the Naturhistorisches Museum Wien (NMW), and therefore should still be in that institution. Before this investigation, four lots of fishes, including L. paradoxa in the collection of the NMW were attributed to Natterer as the collector: NMW-50270 (now NMW-50270 pt. a), 16414 (now 50270 pt. b), 50271, and 90977 (in the following we use the presently valid inventory numbers.) Two of the NMW specimens are preserved in alcohol – an almost complete skeleton (NMW-50270 pt.a) (Fig. 1) and a complete specimen (NMW-50271). A third complete specimen is preserved dry (NMW-90977) and there are two large pieces of skin with scales (NMW 50270 pt.b). The presence of remains of three (if not four) specimens attributed to Natterer clearly contradicts the indubitable statement of all authors concerned – including the collector himself – that only two specimens had been collected. Some facts though, allow us to confidently identify one of the original specimens of Natterer (NMW-50270 pt.a), to putatively attribute the skin fragments (NMW-50270 pt.b) to this same type specimen, and to definitely exclude NMW-50271 and NMW-90977 from the type series.
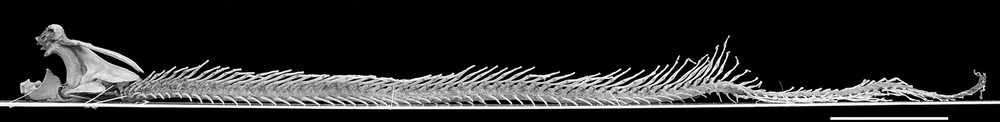
Lepidosiren paradoxa, specimen NMW-50720 pt.a, left side view. Scale bar equals 10 cm.
Lepidosiren paradoxa, spécimen NMW-50720 pt.a, vue latérale gauche. Barre d’échelle égal à 10 cm.
2 The type series
Fitzinger [9] supplied insufficient information to help with the identification of the type specimens of L. paradoxa. On the other hand, Natterer [10] and Bischoff [11] provide identical and quite exact measurements (down to approximately 2 mm) for the two specimens: “3 Fuss 9 Linien” and “1 Fuss 10 Zoll” (3 feet and 9 lines, and 1 foot and 10 inch, respectively). Using the Viennese measuring system then practiced, these measurements correspond to 96.65 and 57.86 cm, respectively (1 Fuss = 0.316081 m, 1 Zoll = 0.02634 m, 1 Linie = 0.002195 m) [12]. This poses problems in trying to attribute type status to any of the Viennese specimens: none of the three complete specimens conforms to these sizes, only the skin parts (NMW-50270 pt.b) might arguably belong to a specimen of approximately 97 cm length. However, it is by no means clear that Natterer, although Viennese, actually applied Viennese measuring standards in this case. Although his article precedes that of Bischoff [11], which is clear from Bischoff's referral to it, it was Bischoff who, in 1837, examined the two specimens in Vienna before Natterer's publication. It seems more than reasonable to assume that Bischoff measured the specimens at that time. Being based in Heidelberg (Germany), where he taught anatomy at the university, he would probably have used Heidelberg standards, quite different from those of Vienna. Also, Natterer – being more of a collector than an anatomist – might very well just have adopted the measurements of the scientist Bischoff for his subsequent publication. This reasoning, seemingly much of a speculation at first sight, is nevertheless strongly supported by the facts described below.
The Heidelberg foot measures only 88% of the Viennese foot [12]. The total lengths (TL) of the two type specimens therefore would be 85.05 and 50.92 cm. Consequently, the TL of the specimens could be either 96.65 and 57.86 (Viennese measure) or 85.05 and 50.92 cm (Heidelberg measure).
The specimen NMW-90977 has 69.8 cm TL, a value fitting neither “Viennese” nor “Heidelberg” lengths. Therefore, it cannot be the type. NMW-50270 pt.b consists of two large fragments of skin with scales, which measure approximately 71 and 57 cm. Those could be the skin of the left and the right side of the larger specimen, but not of the smaller specimen, because even the smallest of the skin fragments is too large to be the skin of the smaller specimen. Both fragments are incomplete, lacking at least the portion that covered the skull and part of the caudal fin, consequently they must have come from a specimen much larger than 57.86 or, obviously, 50.92 cm for the smaller specimen. Whether one or indeed both of the two fragments belong to the larger type specimen, cannot be decided based on unequivocal evidence. There is, however, a considerable probability, that this is the case: we know that Bischoff had been obliged to conserve the skin “as completely as possible” in the course of dissecting the specimen [11].
The specimens NMW-50270 pt.a and NMW-50271 measure approximately 85 and 86 cm, respectively. That is close to the exact measurements reported for the larger Natterer specimen, Heidelberg standard assumed (85.05 cm). This makes them both candidates for being the larger type specimen simply because their respective actual sizes fit the reported TL reasonably well. However, since both have almost the same length, if one is the type, the other consequently is not. Despite the presence of Natterer's name also on the label of NMW-50271, the analysis of Bischoff's work permits us to clearly identify the skeleton (NMW-50270 pt.a, Fig. 1) as belonging to the larger specimen of the type series.
Bischoff, already in 1837, after having seen both specimens (their intestines were only partly preserved) was allowed to analyze and dissect the larger specimen in Heidelberg, at the condition that he left the complete skeleton intact. The skin should also be conserved as completely as possible. He had finished this work and his manuscript by the beginning of 1840. It is unclear, however, whether his illustrations are based partly on both specimens or on just one of them. At least all the figures of the skeleton of L. paradoxa presented by Bischoff are clearly based on one fish only, i.e. the larger type specimen (cf. NMW-50270 pt.a). Other drawings, depicting soft tissues, cannot be readily attributed to a specific specimen. However, there is little doubt that they too belong to the same specimen, given that Bischoff only had one fish for thorough investigation at his disposal in Heidelberg, though he had examined both (we do not know how thoroughly) three years earlier in Vienna. One or both specimens had previously (1837?) been lightly dissected and consequently was incomplete, but Bischoff clearly states that the heart was still present in both specimens, as were the digestive system as well as sexual and urinary organs. He also specifies that both specimens were females.
The lot NMW-50270 pt.a consists of an almost complete skeleton, presently lacking the ceratohyals, subopercles, pectoral, and pelvic girdles and fins. The Meckel's and the nasal cartilages – usually lost during preparation – are preserved. This confirms Bischoff's text, mentioning their presence. Analyzing Bischoff's drawings, it becomes clear that the pectoral and pelvic girdles had been separated during preparation, since those structures are shown independently, and do not appear in his illustration of the reconstructed complete body. The whereabouts of those missing structures are unknown.
The skull roof is entirely composed of a large frontoparietal bone and of the styliform supraorbitals. In many specimens of L. paradoxa there is an unossified area behind the point of articulation between the supraorbital and the frontoparietal. In NMW-50270 pt.a, that is not the case: this region is completely ossified, probably because the specimen is a large adult. In front of the frontoparietal is the almost triangular dermethmoid bone, which is fractured but complete. The nasal cartilages are still preserved on either side of the dermethmoid, and close to the anterior end of this bone is the pair of vomerian teeth. The pterygoid is located below the frontoparietal, and in its anterior end supports the upper tooth plate. The ligaments connecting the pterygoid and the frontoparietal on both sides are preserved on the surface of contact between these bones. Completing the side of the skull is the squamosal, which covers the lower projection of the frontoparietal and the posterior part of the pterygoid. At the anterior extremity of the squamosal is the condyle for articulating with the lower jaw. The styliform opercle contacts the squamosal behind this condyle.
The basicranium is completely composed of the parasphenoid of almost lozenge shape and a cartilage or ligament, which occupies the unossified area anterior to this bone. On each side of the rear portion of the skull is a globular cartilage, which fills an unossified area and makes contact internally with the parasphenoid, laterally with the squamosal and dorsally with the frontoparietal. These globular cartilages normally house the otoliths. Since these cartilages are complete on both sides, the otoliths cannot be seen, and is probably the reason why Bischoff did not mention them in his description, but there is no reason to assume that they are not preserved. In the occipital region there is only a pair of exoccipitals and the large and tubular cranial ribs, which are connected by ligaments to those bones.
The lower jaw is preserved and comprises a large pair of prearticular bones, with high coronoid processes, a pair of triangular angular bones, and the remains of Meckel's cartilage. The lower tooth plate, with three cusps, is placed at the anterior end of the prearticular. The cusps of the left lower tooth plate are considerably more worn than those of the right lower tooth plate. On each side of the mandibular symphysis and below the lower tooth plates is located a thin anterior portion of the Meckel's cartilage.
The axial skeleton is well preserved. There are no vertebral arcocentra. Instead of that, the vertebral elements are supported by the notochord. As described by Bischoff, there are 55 pairs of ribs, followed by 25 hemal elements. The dorsal part of the axial skeleton is composed of 83 neural elements and, as stated above, the apendicular skeleton is missing.
Information drawn from Bischoff's text and especially his illustrations allow us to identify the skeleton NMW-50270 pt.a as belonging to the larger specimen collected by Natterer. Most remarkable is a fracture on the dermethmoid, described by Bischoff as “a kind of irregular suture or scar, which I do not believe to be natural, but brought about by accident only” ([11], p. 9). Because of this fracture, the bone appears with an unnatural angle of almost 90° in its median portion. This same fracture, as well as the angle of the dermethmoid, is represented in Bischoff's drawings (Figs. 2 and 3). This by itself is sufficient to identify NMW-50270 pt.a beyond any doubt as the depicted specimen, but additional evidence also supports this.
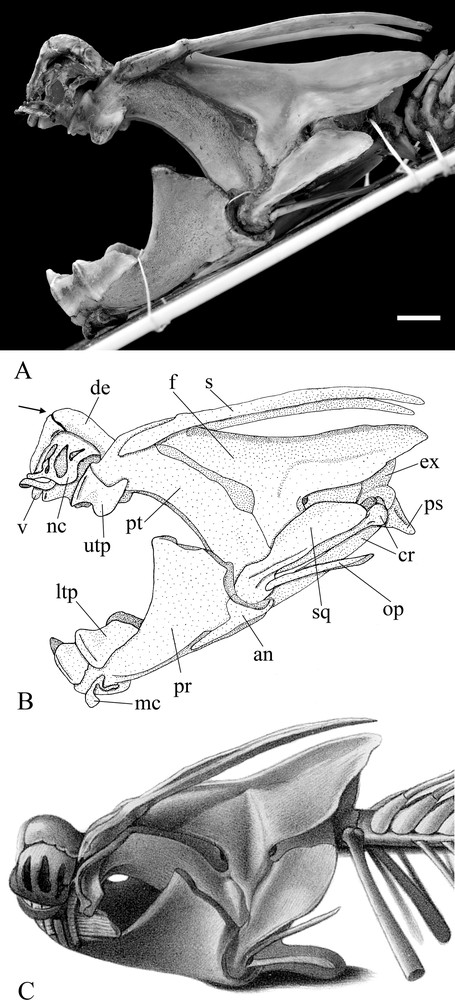
Lepidosiren paradoxa, specimen NMW-50720 pt.a, left side view of the skull. Photo (a), drawing (b), and Bischoff's (1840) reconstruction (c). Abbreviations: an: angular; cr: cranial rib; de: dermethmoid; ex: exoccipital; f: frontal; ltp: lower tooth plate; mc: Meckel's cartilage; nc: nasal cartilage; op: opercle; pr: prearticular; ps: parasphenoid; pt: pterigoid; s: supraorbital; sq: squamosal; upt: upper tooth plate; v: vomerian tooth. Arrow indicates the fracture on the dermethmoid. Scale bar equals 10 mm.
Lepidosiren paradoxa, spécimen NMW-50720 pt.a, crâne en vue latérale gauche. Photo (a), dessin (b) et reconstruction de Bischoff (1840) (c). Abréviations : an : angular ; cr : côte crânienne ; de : dermoethmoïde ; ex : exoccipital ; f : frontal ; ltp : plaque dentaire inférieure ; mc : cartilage de Meckel ; nc : cartilage nasal ; op : opercule ; pr : préarticulaire ; ps : parasphénoïde ; pt : pterigoïde ; s : supraorbitaire ; sq : squamosal ; upt : plaque dentaire supérieure ; v : dent vomerian. La fleche indique la fracture du dermoethmoïde. Barre d’échelle égal à 10 cm.

Lepidosiren paradoxa, specimen NMW-50720 pt.a, dorsal view of the skull. Photo (a), drawing (b), and Bischoff's (1840) reconstruction (c). Abbreviations: de: dermethmoid; f: frontal; lc: labial cartilage; ltp: lower tooth plate; nc: nasal cartilage; op: opercle; pr: prearticular; pt: pterigoid; s: supraorbital; sq: squamosal; upt: upper tooth plate. Arrows indicate the fracture on the dermethmoid and marks on the skull. Scale bar equals 10 mm.
Lepidosiren paradoxa, spécimen NMW-50720 pt.a, crâne en vue dorsale. Photo (a), dessin (b) et reconstruction de Bischoff (1840) (c). Abréviations : de : dermethmoïde ; f : frontal ; lc : cartilage labiale ; ltp : plaque dentaire inférieure ; nc : cartilage nasal ; op : opercule ; pr : préarticulaire ; pt : pterigoïde ; s : supraorbitaire ; sq : squamosal ; upt : plaque dentaire supérieure. Les flèches indiquent la fracture du dermoethmoïde et des marques sur le crâne. Barre d’échelle égal à 10 cm.
There are several peculiarities depicted in Bischoff's reconstruction (Fig. 3), which correspond to skeleton NMW-50270 pt.a, such as a small deepening on the dorsal surface of the left supraorbital and a protuberance close to the posterior end, on the left margin of the frontoparietal. Another example is the rare presence of some cartilages, which are normally lost during preparation (cf., nasal and Meckel's cartilages). Finally, there is the wear of the left lower tooth plate, faithfully represented by Bischoff.
The sum of these pieces of evidence, together with the size of the specimen correspond almost exactly to the one given in the description – when Heidelberg measuring standards are considered –, leaves no doubt that NMW-50270 pt.a is indeed the skeleton of the larger syntype which Fitzinger had at his disposal when naming the species. Since NMW-50270 pt.a has almost the same size as NMW-50271, the latter cannot be the other syntype, nor can it, indeed, have been collected by Natterer. Interpreting the available sources, this lot (whose original label is not preserved) must be regarded now as having been mislabeled.
Concerning the two fragments of skin (formerly NMW-16414, now NMW 50270 pt.b), we suggest that probably they also are part of the larger type specimen. Although there is no definite evidence to confirm this, the following reasoning supports this view: The mere and extremely rare fact that skin fragments are present at all (the NMW only counts a full hand of skin fragments among several hundred thousands of specimens), and the circumstance that this presence could be expected because of Bischoff's explicit statement of having been obliged to keep the skin intact, are evidence that these fragments belong to the same specimen as the skeleton NMW-50270 pt.a. Supported by and in accordance with article 72.4.1.1 of the ICZN [13] (any evidence can be taken into account for determining the type series), these skin fragments can be regarded as putative parts of the larger syntype specimen. Because of this and in order to stress provenance from the same specimen as NMW 50270 pt.a, the former lot NMW-16414 has been renumbered and is now inventoried as NMW 50270 pt. b (ex NMW-16414).
As stated above, NMW-90977 does not fit the measurements of either syntype and consequently is not part of the type series. The locality of this specimen cannot be determined, as Natterer does not mention which specimen was collected in which of the two indicated localities. It is clear, however, that this specimen comes from the Amazon Basin, and its locality is either a creek near Borba in the Madeira River drainage or a swamp close to “Vila-Nova”, in the Amazon River drainage.
3 Conclusion
L. paradoxa was the first living lungfish species to be described. Its discovery, followed by the description of the other living dipnoans, helped to gain a better understanding of the origin of tetrapods and the transition of vertebrates from water to land. Although the provenance of the specimen described here and collected by Natterer cannot be identified down to the Madeira River or the Amazonas River, NMW-50270 pt.a unequivocally is the skeleton of one of the original syntypes used by Fitzinger in the description of L. paradoxa; and NMW-50270 pt.b is putatively attributed to the same specimen.
Acknowledgments
We sincerely thank Ernst Mikschi for allowing us to examine the specimens under his care and David Martill, Agnès Dettäi, Wilson Costa, and the reviewers for suggestions on the manuscript. LPM and PMB acknowledge the support of CNPq and CAPES (Brazilian Government).