

Abridged English version
Palaeoecological investigations are of vital importance for understanding past vegetation and climate changes, especially for the Mediterranean area where a drastic increasing aridity has been observed during the last decades. From Morocco and Tunisia some studies are available, but they are extremely rare from Algeria. We present here the pollen analyses of two littoral marshes from north-east Algeria, as a first attempt of gathering new data.
The sites studied
The two marshes of Bourdim (8° 15′ 14′′ E, 36° 48′ 12′′ N, 13 m a.s.l.) and Garaat El-Ouez (8° 20′ E, 36° 49′ 06′′ N, 45 m a.s.l.) belong to the large El-Kala littoral wetland complex close to the Tunisian border; they lie on the Miocene sandstones and Pleistocene deposits. The local bioclimate is “thermo-mediterranean”, with mean annual rainfall being above 700 mm (944 mm at El-Kala); the dry season lasting from May to September and the mean annual temperature being 17.9 °C. Although dense cork oak forests are still present in the region with deciduous Quercus faginea forests at wet places and higher altitudes, the natural vegetation around the sites which is most probably composed of open wood stands with Quercus suber and Quercus coccifera, mattorals with Erica arborea, meadows, is damaged by intensive agriculture and pastoralism. The marshes themselves are occupied by different aquatic to halophytic formations including more or less disturbed Alnus glutinosa fens.
Methods
The cores were obtained with a Russian corer in 1998. The simplified lithostratigraphies of Bourdim (660 cm) and Garaat El-Ouez (480 cm) appear at the left of the pollen diagrams. After pollen extraction, 67 spectra were established for Bourdim and 48 for Garaat El-Ouez, based on minimum of 300 upland pollen grains. Pollen grains of aquatic taxa, pteridophytes and Salix, Alnus are excluded from the pollen sums in the percentage calculations. Eight 14C dates were obtained from bulk sediments. The incoherencies between some of them make difficult the establishment of a robust chronological framework.
The diagrams, vegetation history and discussion
The Bourdim sequence provided a detailed record of Late Holocene vegetation dynamics. This record reflects open surrounding vegetation with a dominant E. arborea mattoral which is most probably a result of the destruction of a former cork oak forest. This advanced degradation suggests a very recent age, as the pollen data from the Tunisian sites never show a strong forest decline before the Roman times. This consideration leads us to accept the 14C age of 1255 ± 75 uncal B.P. at 590–595 cm and reject the two ages of 2855 ± 51 uncal BP at 635 cm and 1977 ± 34 uncal BP at 345 cm. Contrarily, if the two last ages are accepted, one must admit that either the forest has been destroyed as soon as the Bronze age or at some places the late Holocene “natural” vegetation could have been composed of a dense E. arborea mattoral as was evidenced in Corsica. The only major change observed in the diagram is local: the replacement of an open (permanent) water ecosystem by an Alnus/Salix fen (transition of Bour-d to Bour-e pollen zone). Additional 14C ages are needed to establish a firm chronology and to date the exact time of this water table lowering. The Garaat El-Ouez sequence is divided into 5 pollen zones (Gar-a to Gar-c). Pollen evidences and 14C ages clearly point out to a hiatus at 60/70 cm between a late Holocene peat and organic sediments accumulated at the very end of the last Pleniglacial. The Holocene section gives the image of disturbed oak forests mixed with an E. arborea mattoral, as at Bourdim.
For the first time in Algeria, the vegetation during the last glacial period is described. All along the glacial part of the sequence, pine (most probably Pinus halepensis) is the dominant tree; the abundance of Poaceae associated with different steppe taxa suggests a forest-steppe. Pine is extremely rare in the Holocene pollen sequences from Tunisia. In Algeria, the pollen sequence of Oum el-Khaled, close to Tebessa suggests the dominance of P. halepensis stands mixed with Quercus ilex since early Holocene until 4000 B.P., when it is replaced by a Stipa steppe. The P. halepensis open forests observed today in the Aures Mountains could constitute a possible analogue landscape. Cedrus pollen is represented by a continuous curve (frequently above 5 %) during this period suggesting that the tree was present in the region. Abies is regularly present. As in Dar Fatma in Tunisia, our record demonstrates that, during the last glaciation, Abies pinsapo (as Cedrus) was largely disseminated in NW Maghreb. The presence of different thermophilous trees suggests a dry but relatively mild climate at that time.
As a conclusion, our data confirm that NW Maghreb played the role of a glacial refugium for thermomesophilous trees. Although the set of 14C dates available today does not allow a precise chronological discussion, we show the great potential of east Algerian wet zones for the reconstruction of past vegetation and climate. The great age differences in the infilling of our two study sites requires a systematic exploration of all the potential sites, and a multidisciplinary palaeoecological investigation of them to better understand their origins and setting as well as a more detailed reconstruction of the past vegetation, landscape and climate of the area.
1 Introduction
Le réchauffement climatique et ses conséquences suscitent un grand nombre d’interrogations. Modifications des écosystèmes, sécheresses, érosion, événements météorologiques soudains ou encore exodes de populations sont autant de défis qu’il faudra surmonter si l’augmentation des températures à l’échelle planétaire se poursuit.
Parmi les régions du globe les plus menacées, la zone méditerranéenne se situe dans des contextes bioclimatique et biogéographique particulièrement vulnérants : elle se trouve à la confluence du climat aride nord-africain et du climat tempéré humide central-européen, subissant des interactions complexes entre une influence nord-atlantique, et des influences subtropicales, voire tropicales [1]. Une grande fragilité de la région méditerranéenne à toute variation significative du climat mondial résulte de cet équilibre ténu [2,3]. De plus, la dégradation et la fragmentation récurrentes des espaces naturels par l’action anthropique y est très ancienne. Ainsi, il subsiste l’épineuse question d’un déterminisme climatique et/ou anthropique des changements paléoenvironnementaux dont témoignent les enregistrements paléoécologiques sur le pourtour méditerranéen, et ce, en dépit des nombreuses études consacrées à ce sujet [4–7].
L’enjeu est crucial pour le Maghreb méditerranéen. Cette région compte parmi les priorités mondiales de conservation en tant que « hotspot » de biodiversité [8] en raison d’un fort taux d’endémisme végétal, d’une grande richesse taxonomique et d’une pression humaine élevée [9].
Le Maghreb est également aux premières loges de la menace climatique. Plusieurs études prospectives concluent à une prochaine aridification [10] voire une désertification de l’ensemble de la frange sud-méditerranéenne [11]. Dans ce contexte, la connaissance des dynamiques passées des écosystèmes en Afrique du Nord reste primordiale car les données paléoécologiques :
- • fournissent des éléments décisifs pour la compréhension des changements écologiques à long terme en relation avec des épisodes climatiques passés ;
- • peuvent mettre en évidence le rôle de l’homme et de ses pratiques agropastorales dans la mise en place et la dégradation des paysages nord-africains depuis le Mésolithique ;
- • permettent également d’éprouver la solidité de modèles prospectifs sur des pas de temps supérieurs à ceux livrés par des séries de données instrumentales [12].
L’histoire de la végétation, du climat et de l’occupation humaine au Maghreb reste cependant très largement méconnue en raison d’un nombre très insuffisant d’études palynologiques, le Maroc est certainement une région la mieux documentée. Les travaux de Ballouche [13], Ballouche et Damblon [14], Bernard et Reille [15], Cheddadi et al. [16], Damblon [17], El-Hamouti et al. [18], Lamb et al. [19], Reille [20,21] ont permis de retracer les grandes étapes des dynamiques végétales tardiglaciaires et holocènes, avec cependant de grandes lacunes temporelles et géographiques. Pour la Tunisie, citons Ben Tiba [22,23] et Ben Tiba et Reille [24], Damblon et Van den Berghen [25], Stambouli [26], Stambouli-Essassi [27] dont les contributions majeures à la connaissance du passé écologique de la région soulignent également des lacunes fondamentales.
Enfin, en Algérie, seuls les travaux de Ritchie [28], de Salamani [29,30] et de Benslama [31] ont abordé la question de la dynamique holocène de la végétation. Les connaissances sur l’histoire de la végétation et de l’action de l’Homme pendant l’Holocène en Algérie sont par ailleurs très limitées. La séquence holocène d’Oum el-Khaled (Monts de Nementcha) témoigne d’épisodes récents de déboisement et de pratiques agropastorales pendant la période subatlantique [28]. Ces événements ne sont malheureusement pas datés. En Grande Kabylie, des traces de la mise en culture de l’olivier et de céréales sont enregistrées tardivement dans les séquences de La Châtaigneraie et de Tagma étudiées par Salamani [29] entre 810 ± 180 B.P. et 350 ± 200 uncal. B.P., soit très postérieurement à la néolithisation.
La présente étude de deux enregistrements paléoécologiques prélevés dans le nord-est algérien (région d’El-Kala) représente une contribution inédite à l’histoire nord-africaine de la végétation. Le présent travail propose de reconstituer les dynamiques écologiques passées de la région et d’évaluer les impacts respectifs des activités humaines et du climat des 15 derniers millénaires. Au-delà de cette problématique de première importance, le manuscrit aborde également la place et la signification biogéographiques d’espèces ligneuses clés comme Cedrus ou encore Myrtus, dont le maintien est mis en péril par la pression anthropique croissante et le changement climatique.
2 Les sites étudiés et leur contexte
2.1 Cadre géographique et géomorphologique
La région d’El-Kala est une des zones du nord-est de l’Algérie les plus riches en biodiversité et soumise à une protection, avec la mise en place depuis 1983 d’un Parc National (d’une superficie de 76 438 hectares) et inscrite en tant que réserve de la Biosphère par l’UNESCO depuis 1990. La région est limitée à l’est par la frontière algéro-tunisienne, au nord par la mer, à l’ouest par le Cap Rosa, au sud par les contreforts du djebel El Ghorra.
Notre étude a porté sur deux marais littoraux, Bourdim et Garaat El-Ouez, appartenant à un vaste complexe humide séparé de la Méditerranée par des cordons dunaires, délimité au sud et à l’est par la terminaison orientale de l’Atlas Tellien (Fig. 1). Ces zones humides, organisées en mosaïques d’écosystèmes forestiers, lagunaires, et parfois marins, reposent sur des grès et argiles de Numidie miocènes et des dépôts quaternaires dans des zones subsidentes [32]. L’alimentation en eau de ces zones humides est assurée par la nappe dunaire, le débordement de l’oued Kebir et par le ruissellement des eaux de surface sur les argiles imperméables (créant des ravines à écoulements intermittents ou « chabats »).
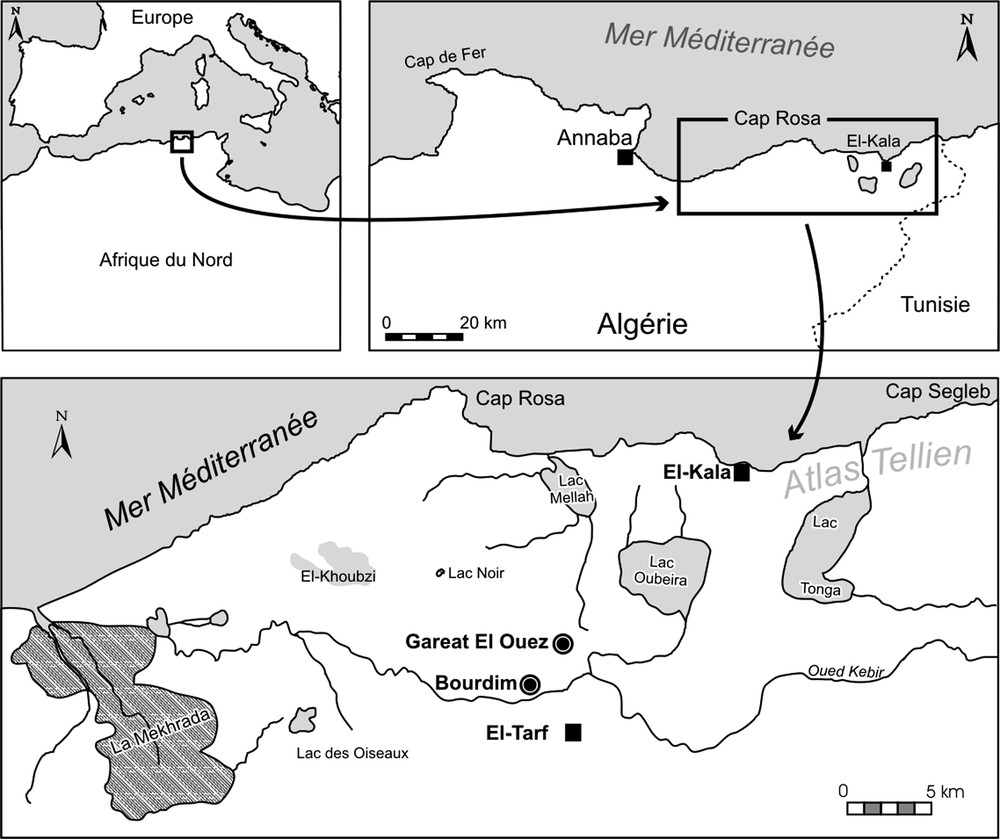
Localisation des sites étudiés.
Le marais de Bourdim (8° 15′ 14′′E, 36°48′ 12′′ N, altitude entre 10 et 16 m) occupe la vallée au nord ouest de Bouteldja (Fig. 1) ; sa superficie varie de 2 à 15 ha (lors des grandes eaux).
Garaat El-Ouez (8° 20′ E, 36° 49′ 06′′ N, altitude 45 m) s’est développé dans une dépression sur un replat des grès et argiles de Numidie (Fig. 1). Cette succession de marais, (d’une superficie d’environ 10 ha) est étagée entre 40 et 50 m d’altitude.
2.2 Climat et végétation actuelle
La région est soumise à un climat méditerranéen dont la saison sèche dure au maximum quatre mois (mi-mai à mi-septembre), mais les précipitations moyennes annuelles sont partout supérieures à 700 mm/an, augmentant d’ouest en est et avec l’altitude. Ainsi, on rencontre dans la région des bioclimats méditerranéens humides (717 mm/an à Bouteldja, 944 mm/an à El-Kala) à perhumide (1200 mm/an à El-Aioun, alt. : 700 m ; 1500 mm/an à El-Ghoura, alt. : 1202 m).
Dans la région d’El Kala, l’humidité atmosphérique moyenne annuelle est de l’ordre de 72 % la température moyenne annuelle est de 17,9 °C avec une amplitude moyenne de 6,75 °C.
Ce climat est particulièrement favorable au développement d’une végétation forestière : 72,25 % de la superficie de la région d’El Kala est composée de forêts, 9 % de terres agricoles, 7 % de lacs, et enfin 11 % de ripisylves et marais. Le couvert végétal est largement dominé par une subéraie à sous-bois d’Erica arborea L. et de Myrtus communis L., alors que les formations dunaires sont occupées par Quercus coccifera L.
L’aulnaie représente également un petit écosystème fragile et original de la région puisqu’il est principalement alimenté par les eaux des crues des oueds en période hivernale. Ces formations d’aulnaies comportent des arbres de grande taille (20 m de hauteur en moyenne) abritant une forte diversité en oiseaux. Cette aulnaie est actuellement dégradée et en forte régression.
Des séries de végétation ont été décrites pour la région [33], et nous ne présentons que la végétation environnant les deux sites d’étude dans le Tableau 1, classée en trois groupements distincts :
- • une végétation aquatique, constituée de Nymphaea alba, Callitriche palustris, Ranunculus aquatilis, Polygonum salicifolium et Salvinia natans ;
- • une végétation amphibie avec Scirpus lacustris, Phragmites australis, Typha latifolia, Juncus capitatus ;
- • une végétation mésophile, représentée par des prairies à Orminis mixta, Mentha rotundifolia, Trifolium maritimum ;
- • une végétation plus hélophile, représentée par l’aulnaie (Alnus glutinosa) associée à Salix pedicellata, Erica scoparia, Rubus ulmifolius, Vitis vinifera, Athyrium filix-femina, Osmunda regalis, Laurus nobilis.
Description des principales formations forestières à proximité des sites étudiés.
| Sites | Marais de Bourdim | Garaat-El-Ouez |
| Végétation locale | ||
| Subéraie mixte dégradée à Quercus coccifera et à Halimium halimifolium Magnocariçaie à Cladium mariscus, Juncus effusus, Carex elata et Nypheae alba Aulnaie à Alnus glutinosa Frênaie humide à Fraxinus angustifolia Subéraie-zénaie et cocciféraie dunaire |
Subéraie dégradée Aulnaie dégradée Magnocariçaie à Cladium mariscus et Juncus effusus |
2.3 Prélèvements et traitements
Les sondages ont été réalisés à l’aide d’un carottier russe [34]. Le site de Bourdim a livré un profil de 660 cm et Garaat El-Ouez un profil de 480 cm. Les lithologies simplifiées des profils de Bourdim et de Garaat El-Ouez sont respectivement présentées dans les Fig. 2 et 3.
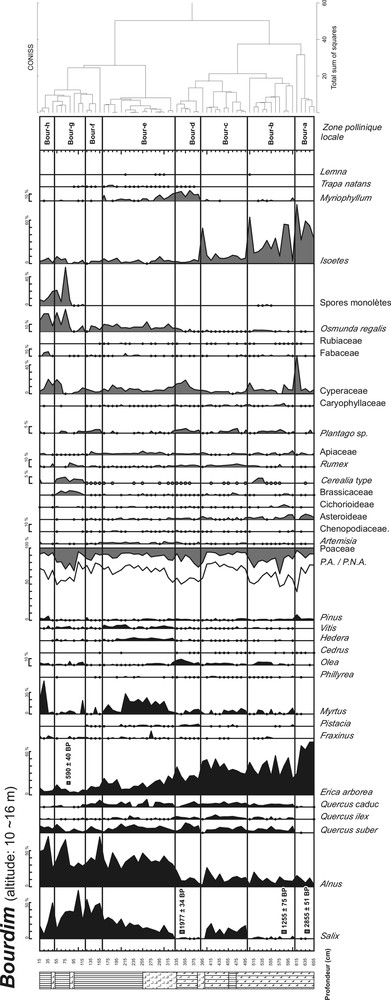
Diagramme pollinique et lithologie (légende figure suivante) simplifiés du site de Bourdim.
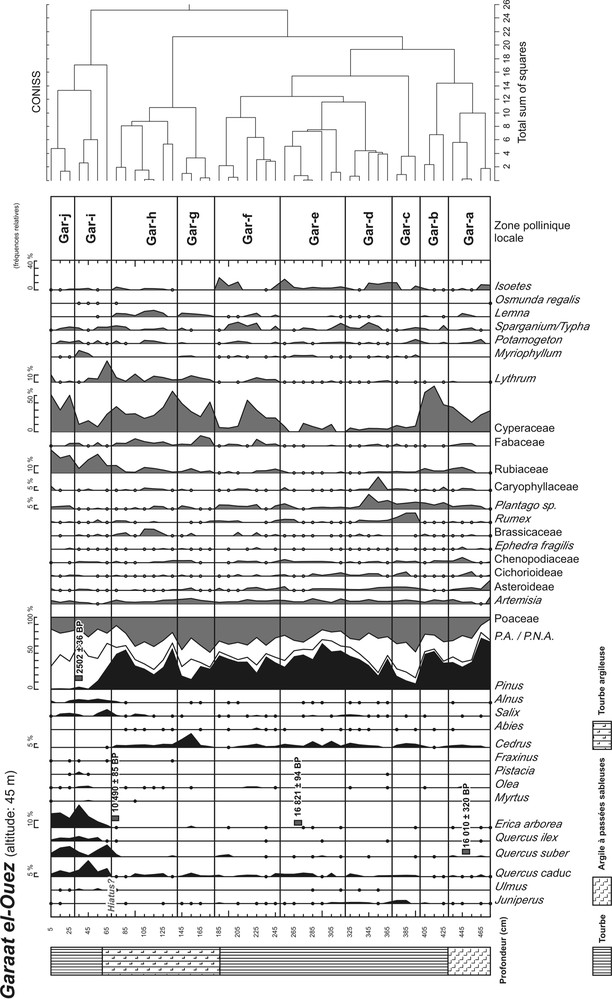
Diagramme pollinique et lithologie simplifiés du profil de Garaat El-Ouez.
Les sédiments de nature organique ont été traités par la méthode dite classique [35] et les échantillons argileux par flottation sur la liqueur de Thoulet [36]. La détermination des grains de pollen a été effectuée au microscope optique, sous objectif à immersion d’huile à l’aide de collections et d’un atlas pollinique de référence [37]. Les effectifs sont exprimés en fréquences relatives (pourcentages par rapport à la somme pollinique totale, spores exclues) et représentés graphiquement sous forme de diagrammes.
2.4 Diagrammes polliniques : Zonation
Les diagrammes de Bourdim (67 spectres) et de Garaat El-Ouez (48 spectres) sont respectivement présentés dans les Fig. 2 et 3. Un minimum de 300 grains de pollen a été compté dans chaque niveau afin d’assurer la représentativité statistique de chaque spectre analysé. Les diagrammes polliniques en fréquences relatives présentent l’évolution des principaux taxons rencontrés lors de l’analyse. Les effectifs des Ptéridophytes et de plantes aquatiques ont été exclus de la somme pollinique. De même, les forts effectifs observés d’Alnus de Salix et de Cyperaceae (surabondants localement) minorent la représentation des autres taxons dans la pluie pollinique ; leurs effectifs ont donc également été exclus de la somme pollinique totale afin d’améliorer la représentation graphique des autres taxons polliniques.
Quatre datations 14C ont été obtenues sur tourbe et sédiment (pas de macrorestes) pour chaque site au Centre de Datation par le Radiocarbone de Villeurbanne (Université Lyon I). Le Tableau 2 présente les âges obtenus. La calibration à 2 σ a été effectuée grâce au logiciel Calib 6 [38] en accord avec la courbe de calibration intcal09 [39].
Datations 14C obtenues pour les deux profils algériens.
| Site | Niveaux datés (cm) | Réf. Lab. | Matériel daté | Âge 14C non calibré (années B.P.) | Âges 14C calibrés à 2 σ (années B.P.) (cal B.P.) |
| Bourdim | 78–82 | Ly-9273 | Tourbe | 590 ± 40 | 535–654 |
| 345–350 | AA50110 | Tourbe | 1977 ± 34 | 1865–1997 | |
| 585–590 | Ly-9272 | Tourbe | 1255 ± 75 | 1051–1301 | |
| 635–640 | AA50112 | Tourbe | 2855 ± 51 | 3088–3158 | |
| Garaat El-Ouez | 35–40 | AA50105 | Tourbe | 2502 ± 36 | 2458–2739 |
| 75–80 | Ly-9275 | Tourbe | 10 490 ± 85 | 12 109–12 598 | |
| 265–270 | AA50108 | Tourbe | 16 821 ± 94 | 19 775–20 296 | |
| 445–450 | Ly-9274 | Sédiment argileux | 16 010 ± 320 | 19 666–19 867 |
La zonation du diagramme a été établie : (1) grâce à une analyse préliminaire par clustering du logiciel CONISS [40] ; et, (2) en tenant compte des variations notables de fréquences des taxons forestiers et herbacés majeurs. Une zone pollinique est définie par une série d’assemblages polliniques de niveaux voisins qui ne montrent pas de différence majeure entre eux. L’étape de la zonation est primordiale car elle permet d’agencer la description du diagramme pollinique, et par conséquent l’histoire de la végétation retracée. Les Tableaux 3 et 4 résument les critères biostratigraphiques et les assemblages polliniques qui ont permis de définir chaque zone pollinique des deux diagrammes. En l’absence de cadre palynostratigraphique de référence pour la région, aucune chronozone n’a pu être définie avec certitude.
Description des zones polliniques définies pour le profil de Bourdim.
| Zone pollinique | Description de la végétation |
| Bour-h | Régression des Poaceae au profit des éléments de la subéraie |
| Retour de Myrtus | |
| Fin des courbes continues de Cerealia et des Brassicaceae | |
| Bour-g | Diminution du rapport pollen arboréen/pollen non arboréen PA/PNA |
| Courbe continue de Cerealia | |
| Courbe continue des Brassicaceae | |
| Augmentation des taux de Poaceae | |
| Bour-f | Maintien d’Alnus et de Salix |
| Régression de Myrtus | |
| Redéploiement des taux d’Erica | |
| Fin de courbes continues de Vitis et d’Hedera | |
| Bour-e | Fort développement d’Alnus associé à Salix ; recrudescence d’Osmunda regalis |
| Maintien de Quercus suber | |
| Déclin d’Erica | |
| Optimum de Myrtus | |
| Courbes continues de Vitis et d’Hedera | |
| Bour-d | Optimum de Quercus suber |
| Optimum d’Olea | |
| Courbe significative de Myriophyllum et occurrences de Trapa natans | |
| Chute transitoire d’Erica | |
| Bour-c | Expansion de Salix |
| Accroissement des taux de Quercus suber et de Myrtus | |
| Maintien d’Erica | |
| Augmentation du rapport PA/PNA | |
| Bour-b | Accroissement des taux de Poaceae |
| Recul modéré d’Erica | |
| Présence de Pistacia et Olea | |
| Courbe significative de Myrtus | |
| Courbe continue de Quercus suber et Quercus caduc | |
| Bour-a | Développement optimal d’Erica arborea |
| Absence quasi totale de Quercus | |
| Abondance d’Isoetes |
Description des zones polliniques définies pour le profil de Garaat-El-Ouez.
| Zone pollinique | Description de la végétation |
| Gar-j | Chute du rapport PA/PNA |
| Augmentation des taux de Rubiaceae et de Cyperaceae | |
| Courbe continue de Plantago sp. | |
| Gar-i | Déclin de Pinus |
| Courbes continues de Quercus caduc, Quercus suber, Quercus ilex et d’Erica arborea | |
| Fin de courbe continue de Cedrus | |
| Optimum de Lythrum | |
| Gar-h | Déclin de Cedrus |
| Optimum transitoire de Pinus | |
| Phase à Brassicaceae | |
| Gar-g | Optimum de Cedrus |
| Augmentation des taux de Cyperaceae | |
| Courbes continues des Fabaceae et des Rubiaceae | |
| Gar-f | Fin de courbe de Quercus caduc |
| Phase à Plantago sp. Dominant | |
| Courbe continue des Cyperaceae | |
| Gar-e | Optimum du rapport PA/PNA |
| Courbe significative de Quercus caduc | |
| Augmentation des taux de Cedrus | |
| Gar-d | Augmentation du rapport PA/PNA |
| Optimum de Plantago sp. et des Caryophyllaceae | |
| Optimum transitoire de Cedrus | |
| Gar-c | Déclin de Pinus |
| Courbes continues de Cedrus et de Quercus caduc | |
| Optimum de Rumex | |
| Chute des Cyperaceae | |
| Gar-b | Optimum des Cyperaceae |
| Augmentation des taux de Pinus | |
| Déclin transitoire des Poaceae | |
| Gar-a | Forts taux de Pinus (70 %), puis déclin |
| Augmentation des taux de Poaceae et d’Artemisia |
3 Le site de Bourdim : une histoire holocène originale
3.1 Chronologie
Le profil pollinique de Bourdim reflète une image détaillée mais relativement récente de l’histoire de la végétation régionale. Les quatre datations obtenues sont incohérentes puisque la continuité stratigraphique n’est pas respectée par celles-ci. Si l’âge 1255 ± 75 B.P. est exclu, la chronologie semble plus cohérente. Néanmoins, en l’absence d’éléments plus objectifs, il semble arbitraire d’invalider cette date plutôt qu’une autre. Cela pose par ailleurs le problème de possibles remaniements sédimentaires. Le contexte sédimentaire de ces marais littoraux est favorable à des phénomènes de pollutions organiques (apports marins, remaniements de dépôts anciens, bioturbation, piétinement dans le marais). Cependant, les dynamiques végétales retracées dans le diagramme ne semblent pas hachées ou interrompues, ce qui suggère l’absence de remaniements sédimentaires. On peut raisonnablement penser que les incohérences des dates sont imputables à de probables pollutions par de la matière organique remaniée. L’interprétation du diagramme holocène de Bourdim ne peut donc pas s’appuyer sur un contrôle chronologique absolu solide.
3.2 Dynamiques végétales
Erica arborea est représenté par des taux importants dans la première moitié de l’enregistrement pollinique (supérieurs à 50 % dans la zone Bour-a). Ses fréquences relatives ne diminuent significativement qu’à partir de la zone pollinique d. Les taxons de la subéraie, bien que présents, sont faiblement représentés quantitativement. On pourrait penser que des subéraies clairsemées occupaient les abords du site, avec en sous-bois la dominance d’E. arborea. Ben Tiba [23] a démontré en Kroumirie que Quercus suber était un modeste pollinisateur : l’étude de la pluie pollinique a mis en évidence que sa représentation ne dépassait guère 20 % au sein d’une suberaie alors qu’E. arborea – souvent présent en sous-bois – se caractérisait par une forte production pollinique.
Il paraît évident que les environs du site de Bourdim étaient dominés par une suberaie mixte dont le sous-bois devait être massivement composé par E. arborea pendant la première moitié de l’enregistrement pollinique. Toujours en Tunisie, sur le site du Djebel El Ghorra, la bruyère arborescente a certainement joué un rôle majeur de substitution dans les maquis dégradés [22] ; il est également probable qu’à Bourdim, une dégradation du maquis – antérieure au début de l’enregistrement pollinique – ait favorisé l’essor d’Erica au sein de la suberaie.
En Corse, des formations à bruyères arborescentes semblent avoir représenté un écosystème stable au cours de l’optimum climatique holocène [41]. Il n’est pas exclu qu’en Algérie, E. arborea ait pu former un véritable couvert forestier pendant l’Atlantique (classiquement daté en Europe méditerranéenne entre 8000 et 5500 B.P. uncal B.P.). La base du diagramme témoignerait alors de l’extrême fin de cet optimum. En l’absence de chronologie fiable et d’enregistrements polliniques plus anciens, cette hypothèse demeure néanmoins discutable.
Dans la zone pollinique Bour-d, on observe un accroissement marqué des taux de Poaceae et d’Olea, associé au recul du matorral, avec la diminution des fréquences d’E. arborea : cela pourrait indiquer une intensification de la pression humaine (pastoralisme et culture d’Olea).
Puis Myrtus semble prendre un rôle important dans la dynamique forestière de cette suberaie mixte, sans doute progressivement dégradée. Dans la zone pollinique Bour-e, on observe une nette expansion du myrte alors que les taux d’Erica diminuent. Ces deux essences (caractéristiques de la subéraie de la région d’El-Kala) sont certainement entrées en concurrence : l’optimum de Myrtus (documenté dans la zone pollinique Bour-e) témoignerait t’il d’une transition climatique humide ? Cet accroissement des taux de Myrtus concorde avec le début de la courbe continue d’Osmunda regalis, et l’augmentation significative des fréquences polliniques d’Alnus et de Salix : il semble donc que le développement d’une myrtaie corresponde à la mise en place d’une végétation hygrophile autour du site. Il est d’ailleurs probable qu’une ceinture forestière à Myrtus, Alnus et Salix, (avec O. regalis en sous-bois et sur le marais) se soit installée à Bourdim. Les courbes continues d’Hedera et de Vitis témoignent également de ce proche environnement forestier humide autour du site, composé par des aulnaies littorales comparables à celles observées de nos jours.
À partir de la zone pollinique Bour-g, les taux de pollen d’arbres chutent significativement avec l’augmentation des Poaceae, associées aux Brassicaceae, à Rumex et à Cerealia. Ce dernier est représenté avec Rumex sous la forme d’une courbe continue, indiquant certainement un accroissement de la pression anthropique. La recrudescence d’activités agropastorales dans les environs du site a probablement affecté le couvert forestier car tous les taxons ligneux marquent un recul, à l’exception de Q. suber.
En dépit de son épaisseur importante (6,55 m), la séquence de Bourdim ne décrit qu’une histoire brève (moins de trois millénaires), mais précieuse compte tenu de l’absence de données paléoécologiques pour la région. Le taux de sédimentation organique est donc important, ce qui témoigne d’une productivité importante de l’écosystème palustre de Bourdim. Le site était probablement :
- • d’abord, un marais à faible tranche d’eau comme l’atteste la courbe d’Isoetes (zones polliniques Bour-a à Bour-c, Fig. 2) ;
- • puis le niveau d’eau a diminué (avec l’apparition de Myriophyllum, zones polliniques Bour-d et Bour-e) ;
- • enfin le marais s’est progressivement comblé, envahi par Alnus, Salix et O. regalis (zones polliniques Bour-g à Bourd-h).
On peut également affirmer que le paysage alentour est totalement déboisé au moins depuis un millénaire ; un matorral dense à E. arborea s’est développé et maintenu à la périphérie du marais. Cependant, en dépit du faible couvert arboréen, les indicateurs herbacés d’anthropisation demeurent relativement discrets ; il n’est pas exclu que la végétation de la zone humide ait joué un rôle filtrant de la pluie pollinique extra-locale. Il faut par ailleurs souligner que le faible pourcentage de pollen d’arbres semble en désaccord avec l’état actuel de la végétation régionale estimée comme majoritairement boisée. Une fois encore, la paléoécologie se heurte à l’épineuse question de la caractérisation d’un couvert forestier et la structuration des paysages en zone méditerranéenne [42].
4 Le site de Garaat El-Ouez : une vieille histoire
4.1 Chronologie
Les âges obtenus pour le site de Garaat El-Ouez posent également problème pour des raisons évidentes de discontinuités stratigraphiques. Les trois plus vieux âges indiquent que l’essentiel de l’enregistrement pollinique (de la base à 70 cm de profondeur) est contemporain de la fin de la dernière glaciation, ce qui représente un élément remarquable pour la région. Mais il semble qu’un imposant hiatus soit présent à la transition entre ces niveaux et l’Holocène. Un tel hiatus nous prive de connaissances précieuses sur la recolonisation postglaciaire par la végétation dans la région : le site de Garaat El-Ouez ne permet donc pas de reconstituer les dynamiques végétales avant l’intensification de la pression anthropique à l’Holocène récent.
4.2 Dynamiques végétales
D’une manière générale, la séquence holocène de Garaat el-Ouez évoque un paysage bien plus boisé qu’à Bourdim et la base de l’enregistrement est bien antérieure à celle de Bourdim. Sur la côte septentrionale de Tunisie la séquence d’Abiare [27], à 110 m d’altitude, témoigne d’un paysage aussi dramatiquement dépourvu d’arbres qu’à Bourdim pendant le dernier millénaire alors que celle de Majen El Orbi, proche mais à 200 m montre l’effondrement des chênes caducifoliés au profit d’un matorral à E. arborea, mais aussi de Q. suber qui demeure abondant jusqu’à nos jours.
Le changement abrupt dans les assemblages polliniques à 65 cm indique la présence d’un hiatus, clairement confirmé par les datations 14C : la base de la séquence correspond à la fin du dernier Pléniglaciaire alors que la partie supérieure correspond à un Holocène certainement très récent (ca. Subatlantique), en accord avec l’image pollinique d’une subéraie dégradée à E. arborea, comme cela est documenté à Bourdim. Il est possible que le réchauffement tardiglaciaire ait durablement asséché la zone humide, stoppant la sédimentation organique, et par conséquent la conservation du pollen. Les deux spectres de 55 et 65 cm, encore riches en pollen de pin pourraient s’expliquer par la percolation de pollen d’âge holocène dans les dépôts plus anciens lors de la remise en eau du marais.
Les deux datations inversées de la partie inférieure de la séquence (16 010 uncal B.P. à 445–455 cm et 16 821 ± 94 uncal B.P. à 265–270 cm) interdisent une interprétation chronologique solide des 4 m inférieurs, bien que la séquence pollinique présente des changements significatifs. Notons néanmoins qu’en dépit de la sur-représentation classique de Pinus pendant le Pléniglaciaire, Cedrus est bien représenté dans l’enregistrement. Il connaît un optimum (zone pollinique Gar-d) qui pourrait correspondre au développement de cédraies à basse altitude, par exemple dans la proche chaîne des Babors. Mais la question de la répartition des cédraies en Algérie à la fin du Pléniglaciaire reste cependant non élucidée.
La zone pollinique Gar-e couvre la totalité de la période holocène (postérieure à ca. 12 300 cal. B.P.) mais l’histoire de la végétation est malheureusement condensée sur 50 cm ; il est par ailleurs difficile d’évaluer la possibilité de hiatus dans l’enregistrement, voire de détailler les dynamiques forestières retracées. Néanmoins, il semble qu’E. arborea ait constitué un taxon majeur au cours de la première moitié de l’Holocène, Q. suber semblant plus présent en fin de zone pollinique.
4.3 Vers une meilleure connaissance de l’histoire des forêts pléistocènes et holocènes en Afrique du Nord
La séquence de Garaat El-Ouez est exceptionnelle car c’est la toute première fois qu’est décrit en Algérie l’état de la végétation à la fin du Pléniglaciaire. Le site tunisien de Dar Fatma à 780 m [27] avait révélé une succession forestière extraordinaire marquée par la prédominance du chêne zen pendant le Pléniglaciaire moyen, suivie par une période marquée par un optimum pour les boisements mixtes de conifères (Cedrus, Pinus et Abies) dont l’abondance passée au Maghreb est attestée vers 24 000 B.P. (si l’on considère que les datations 14C disponibles sont valides). Cette découverte atteste que le climat de l’Afrique du nord a été suffisamment clément et surtout humide pour assurer la prospérité d’arbres dont certains ont quasiment disparu, comme Abies numidica, pendant que d’autres, (comme Cedrus atlantica) n’ont subsisté jusqu’à aujourd’hui que dans les zones d’altitude. Entre cette date et la fin du Tardiglaciaire – enregistrée dans le site montagnard de la Chataigneraie, vers 1000 m d’altitude [30] où une cédraie dense est remplacée par un boisement en chênes zéens à la fin du Dryas récent – aucune information n’était disponible jusqu’alors : le site de Garaat El-Ouez comble partiellement cette lacune et délivre un témoignage d’un court intervalle autour de 18 000 B.P., caractérisé par une pinède ouverte (certainement à pins d’Alep), comportant certainement des peuplements de cèdres, comme cela est suggéré par la courbe continue de Cedrus de notre enregistrement. Il faut remarquer également que le pin est extrêmement rare dans les séquences holocènes littorales en Algérie mais également en Tunisie (voir Magen Ben M’Hida in [26]) ; celui-ci s’efface généralement au profit de Quercus canariensis au début de l’Holocène. Seul le diagramme de Oum El-Khaled, à environ 500 m d’altitude, près de Tebessa [28] relate une prédominance du pin d’Alep associé au chêne vert jusqu’à environ 4000 B.P., puis une recrudescence de la steppe à alfa correspondant à une aridification pendant l’Holocène supérieur. La « forêt steppe pléniglaciaire » dont témoigne le site Garaat El-Ouez peut évoquer certains paysages actuels encore boisés en pins d’Alep des Aurès. Cela confirme que pendant la dernière période glaciaire la région d’El Kala était soumise à des températures moyennes annuelles bien inférieures aux actuelles et un abaissement de la pluviométrie de 300 à 400 mm/an.
Le site de Bourdim est également riche d’enseignements, compte tenu de la rareté des archives paléoécologiques en Afrique du Nord. La forte représentation pollinique de Myrtus communis observée après 1977 ± 34 B.P. est originale car cette espèce est généralement présente dans les diagrammes péri-méditerranéens sous la forme de quelques occurrences. Seuls, quelques sites corses [41], espagnols [43,44] et tunisiens [27] ont enregistré des taux importants de Myrtus. Cet arbuste, caractéristique de matorrals méditerranéens, se développe préférentiellement sous climat méditerranéen humide à per-humide (zones à ripisylves) et prospère au sein de mosaïques d’arbustes et d’herbacées méditerranéens thermophiles.
À Bourdim, la recrudescence de M. communis pourrait être attribuée au développement de la zone humide, favorisée par une période climatique plus humide. D’après Leveau [45], l’Afrique du Nord à l’époque romaine était soumise à un climat comparable à l’actuel, mais caractérisé par des précipitations plus importantes : l’optimum du myrte (zone pollinique Bour-e, Fig. 2), postérieur à ca. 2000 B.P. témoignerait alors de ce climat plus humide. L’augmentation des taux de Salix, Alnus glutinosa, ou encore O. regalis indique également le développement local d’une ripisylve, dont la composition était probablement proche des phytocénoses à Osmundo-Alnion observées actuellement en Toscane, en Espagne, en Corse, Sardaigne et Sicile [46].
Une dégradation de la strate arbustive liée à des activités agropastorales, la compétitition interspécifique, et tout changement paléohydrologique a pu également conditionner la dynamique de Myrtus à proximité du site, et dans la région. En effet, l’enregistrement pollinique de Bourdim témoigne de phases récurrentes d’ouverture du milieu : à plusieurs reprises (zones polliniques Bour-b, Bour-c, Bourd-d et Bour-f, Fig. 2) des taux significatifs de Plantago sp., associés à une recrudescence des Poaceae, des Asteroideae et des Apiaceae montrent que la dynamique végétale locale a certainement subi des perturbations d’origine anthropique, comme en témoignent les phases à Olea et à Cerealia observées dans le diagramme (Fig. 2, zones polliniques Bour-b et Bour-d).
Ces épisodes coïncident avec la diminution d’Erica (au profit de Myrtus) dont le déclin semble se précipiter vers 1977 ± 34 uncal. B.P. (1865 cal B.P.–1997 cal B.P.). Cette dégradation du paysage végétal correspond-t-elle à l’implantation romaine dans la région ? A-t-elle débuté antérieurement sous la domination phénicienne ? Ces questions restent ouvertes et nécessitent de nouvelles investigations incluant l’archéologie.
5 Conclusions
Ces nouvelles données confirment le rôle de refuge du nord-ouest du Maghreb pour les taxons mésophiles pendant la dernière glaciation. Une première hypothèse était qu’à partir de ces zones d’Algérie, le cèdre aurait migré vers le Maroc où il n’était pas connu avec certitude antérieurement à 4000 B.P. [19], mais de nouvelles données paléoécologiques et phylogéographiques tendent à démontrer la multiplicité des zones refuges au Maghreb [47]. Les récents travaux sur le lac d’Ifrah au Maroc [48] ont notamment montré qu’à la fin du Pléistocène, le climat du Moyen Atlas était caractérisé par des phases froides mais relativement humides, par conséquent favorables au maintien du cèdre et de la chênaie entre 24 000 et 12 000 B.P., probablement dans des stations isolées.
Compte tenu des incohérences des séries de datations 14C obtenues pour cette étude, il n’est pas envisageable d’aller plus loin dans l’interprétation chronologique des changements observés dans nos deux séquences. Ces dernières ont cependant montré les potentialités des zones humides de l’est algérien et incitent à une stratégie de prospections systématiques. En effet, alors que le rôle de refuges glaciaires joué par les péninsules nord-méditerranéennes a été abondamment discuté, celui de l’Afrique du nord, île méditerranéenne coincée entre le désert et la mer demeure presque inconnu, faute d’un jeu suffisant d’informations paléobiogéographiques. Localement, les grandes différences d’âge entre les remplissages de deux zones humides pourtant voisines imposent également des études géomorphologiques précisant l’origine de la mise en place de ces marais et des formations quaternaires qui leur sont associées. Il faudra (comme Stambouli en Tunisie) explorer des zones humides dans les différents étages de végétation de la région, du littoral perhumide jusqu’aux zones arides. Enfin, une connaissance plus approfondie de l’occupation humaine à l’Holocène en Afrique du Nord est nécessaire afin d’évaluer le rôle structurant des activités anthropiques sur le couvert végétal algérien : seule la reprise d’investigations archéologiques permettrait d’améliorer ce point essentiel.