

1 Introduction
As noted in previous papers, the status and composition of the genus Chaerilus was the subject of a recent taxonomic analysis [1,2] and two distinct groups of species were defined: the ‘variegatus species-group’ and the ‘truncatus species-group’. For further details, readers may refer to [1,2].
Until now, eight species have been described from Cambodia, Laos and Vietnam: Chaerilus petrzelkai Kovařík, 2000, Chaerilus julietteae Lourenço, 2011, Chaerilus phami Lourenço, 2011, Chaerilus anneae Lourenço, 2012, Chaerilus kampuchea Lourenço, 2012 and Chaerilus terueli Kovařík, 2012 from the southern region of the peninsula and Chaerilus laoticus Lourenço and Zhu, 2008 and Chaerilus vietnamicus Lourenço and Zhu, 2008 from the Northern range [2]. These belong to the two distinct groups of species (Fig. 1).

Map of Southeast Asia, showing the distribution of the Chaerilus species known from this region: Chaerilus anneae (black triangle); Chaerilus julietteae (inverted black triangle); Chaerilus kampuchea (black asterisk); Chaerilus laoticus (black flower); Chaerilus pathom sp. n. (black cross) Chaerilus petrzelkai (black star); Chaerilus phami (black square); Chaerilus terueli (open circle); Chaerilus vietnamicus (black circle).
The specimens described here have been found during expeditions led by French zoologist of Russian origin, Prof. Constantine Dawydoff, who in 1929–1934 and 1938–1939 conducted zoological research at Cau Da near Nha-Trang (now Vietnam). However, it is almost certain that the true collector will remain unknown. The specimens were collected in the ‘Pa Thom cave’ in Dien Bien District, west of Dien Bien Phu city in northern Vietnam. The morphological similarities of the new species to other species distributed only in the south range of Cambodia and Vietnam strongly suggests a typical case of vicariance, as synthesized by Bernardi [3], between northern and southern populations in the Southeast Asian peninsula.
2 Pa Thom cave
Geographically located in northern Vietnam in the Dien Bien Province, Dien Bien District (21°18′26″N, 102°55′43″E), Pa Thom cave is situated at an altitude of 1496 m and is part of the Pa Thom Village (Figs. 2 and 3). This location adjoins the border between Vietnam and Laos. The arch-shaped entry is located on the side of a mountain and is 12 m high and 17 m wide. Along the 350 m length of the cave, there are nine arches of different sizes. Conspicuous stones, stalactites and stalagmites are observed. The space inside the cave is quite impressive.

(Color online.) Pa Thom cave, where the scorpion was found. 1. External view. 2. Interior view.
3 Taxonomic remarks
Attempts to clarify the taxonomic status of Chaerilus species have, in most cases, been more or less successful [4]. Kovařík [5], in an approximate revision of the genus, validated 18 species. Following this contribution many other species have been described and an attempt to define ‘species-groups’ was proposed [1,2]. In a very recent publication of a “self-edited and self-financed” “catalogue of scorpions”, Kovařík [6] proposed a severe number of modifications to the composition of the genus Chaerilus, however these changes were unjustified. His decisions varied from classical synonymies, in many cases preceded by an interrogation mark, but also the attribution of the status of nomen dubium to species well described based on well-defined type material deposited in official institutions. The use of nomen dubium is a typical reaction of this (non-academic) author in face of any material he did not examine. Moreover, almost all of the species placed in these ambiguous positions have been described by the first author (WL) and co-authors, demonstrating that the decisions taken by Kovařík [6] are biased by a personal decision based mainly on speculation.
Consequently, all the decisions made by Kovařík [6] are at present systematically rejected, and the validity of the Chaerilus species described by the first author and co-authors is maintained until further detailed studies can be performed. Kovarik's method of work is basically typological (almost philatelic) and no geographic or ecological parameters were taken into consideration. Recent studies suggest that different species of Chaerilus are not well morphologically distinct but rather correspond with micro-endemic populations, and in some cases to vicariant populations. Molecular studies (now in preparation) will certainly bring further evidence to test this hypothesis (Lourenço et al., in preparation).
4 Methods
Illustrations and measurements were made with the aid of a Wild M5 stereo-microscope with a drawing tube (camera lucida) and an ocular micrometer. Measurements follow Stahnke [7] and are given in mm. Trichobothrial notations follow Vachon [8] and morphological terminology mostly follows Hjelle [9].
5 Taxonomic treatment
Chaerilidae Pocock, 1893
Chaerilus Simon, 1877
Chaerilus pathom sp. n. (Figs. 4–6)

Chaerilus pathom sp. n. Male holotype. Habitus (previously identified as Chaerilus truncatus [Vachon in litt.]).
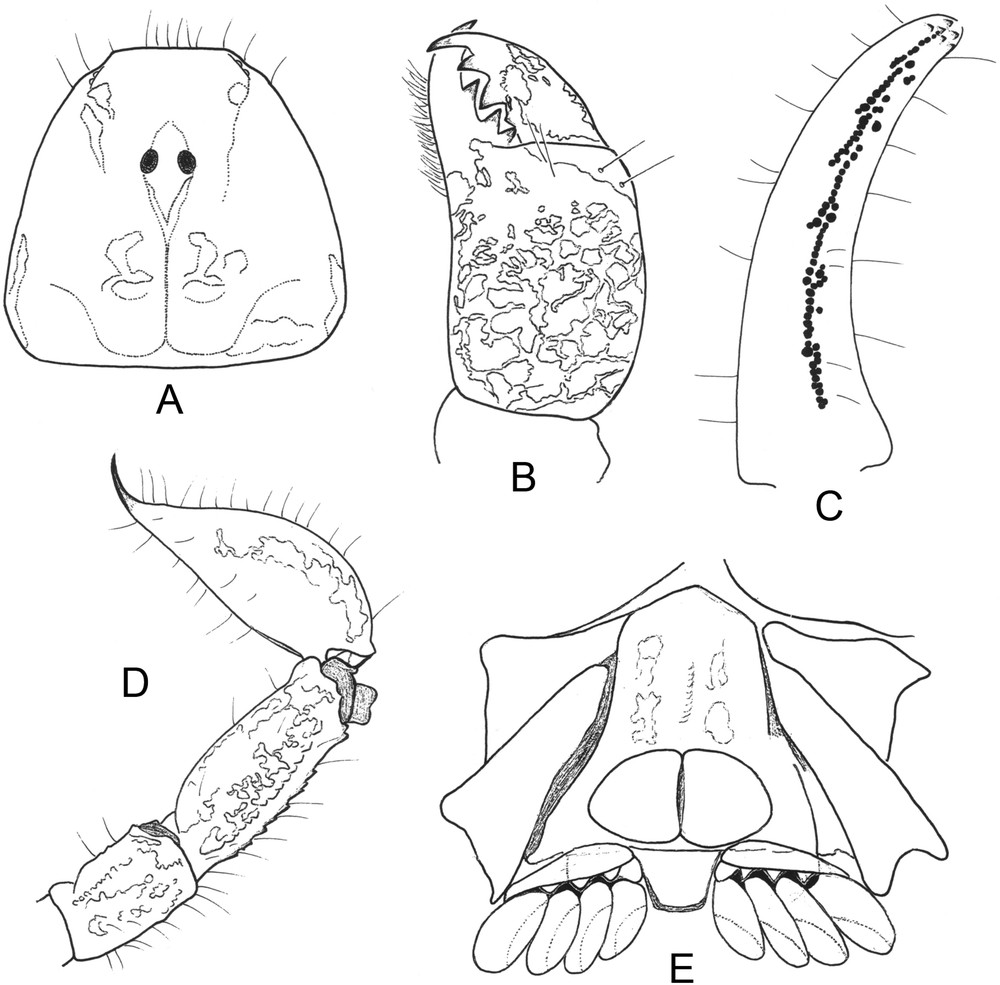
Chaerilus pathom sp. n. Male holotype. A. Carapace, dorsal aspect. B. Chelicera, dorsal aspect. C. Cutting edge of movable finger with rows of granules. D. Metasomal segments IV–V and telson, lateral aspect. E. Ventral aspect, showing coxapophysis, sternum, genital operculum and pectines.
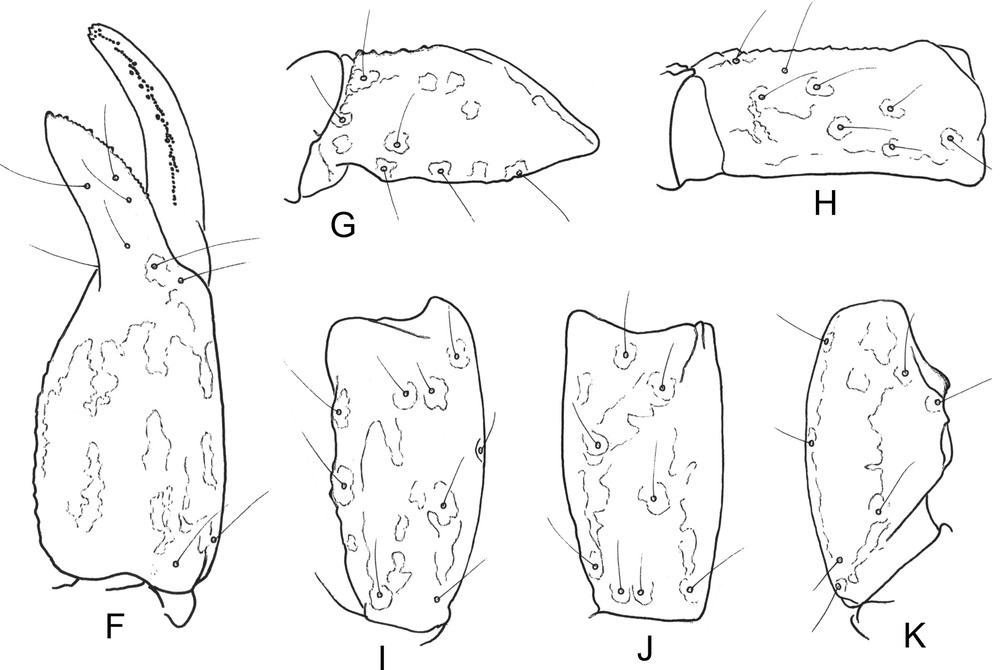
Chaerilus pathom sp. n. Male holotype. Trichobothrial pattern. F. Chela dorso-external aspect. G–H. Femur, dorsal and external aspects. I–K. Patella, dorsal, external and internal aspects.
Material. Vietnam, Dien Bien District, Pa Thom cave, 30 km W of Dien Bien Phu city. Approximately 60 m from entry. Male holotype and one male paratype (C. Dawidoff leg.) 1938–1939 (donation to the Muséum, Paris, 1948). Type material deposited in the Muséum national d’histoire naturelle, Paris (MNHN).
Etymology: the specific name is placed in apposition to the generic name and refers to Pa Thom cave, the location in which the new species was collected.
Diagnosis: the species is relatively small in size compared to that of the other species in the genus, 17.1 mm in total length for male. General coloration yellowish-brown, marked with variegated brownish spots. Carapace strongly narrowed toward the anterior edge; acarinate and weakly granulated to smooth; anterior margin not emarginated; furrows shallow. Metasomal carinae moderately marked; ventral carinae absent from segments I and II, weakly marked on segments III and IV; latero-ventral and ventral carinae on segment V with minute spinoid granules. Telson with pear-like shape; aculeus weakly curved. Dentate margins of fixed and movable fingers with 6–7 rows of granules. Pectinal tooth count 4–4 in male holotype and male paratype. Genital operculum plates have a sub-oval shape. Trichobothriotaxy of type B, orthobothriotaxic.
Relationships: Chaerilus pathom sp. n. shows morphological similarities with some Chaerilus species distributed in the Southern range of Vietnam and Cambodia, namely, C. anneae Lourenço, 2012 and C. kampuchea Lourenço, 2012. The new species can, however, be readily distinguished by a combination of characters:
- • a paler overall coloration, in particular on the ventral aspect, without spots over the genital operculum and pectines;
- • pedipalp chela fingers with 6–7 rows of granulations;
- • carapace and tergites weakly granulated to smooth;
- • metasomal carinae with only weakly marked spinoid granules;
- • distinct morphometric values.
Moreover, the new species was collected in a cave and has a distinct range of distribution.
Description
Coloration: basically yellowish–brown, marked with brownish variegated spots. Carapace yellowish-brown, with dark spots on the anterior and central areas. Tergites yellowish marked with dark confluent spots, more intensely on tergites IV to VII. Metasomal segments yellowish with dark variegated spots. Telson yellowish with dark variegated spots; aculeus pale red. Chelicerae yellowish marked with variegated spots; fingers marked with blackish spots; teeth slightly red. Pedipalps yellowish-brown; femur more intensely spotted than patella and chela; dentate margins of fingers reddish. Legs yellowish with brownish variegated spots. Venter and sternites yellowish marbled with brown; sternite VII more intensely marked; genital operculum and pectines without variegated spots.
Morphology: carapace strongly narrowed anteriorly; anterior margin not emarginated, acarinated and weakly granulated to smooth; furrows shallow. Two pairs of lateral eyes, and one pair of moderate median eyes, more than twice the size of lateral eyes; median eyes anterior to the centre of the carapace. Tergites weakly granulated to smooth; carinae obsolete. Sternum pentagonal, longer than wide; genital operculum plates with sub-oval shape. Pectinal tooth count 4–4 in male holotype. Sternites smooth; spiracles small and round; carinae absent from VII. Metasomal segments I and II wider than long; segment III as long as wide; IV and V longer than wide. All the carinae moderately granular; ventral carinae absent from segments I and II; weakly marked on segments III and IV; dorsal and latero-dorsal carinae with inconspicuous spinoid granules on segments I–IV; latero-ventral and ventral carinae on segment V composed of minute spinoid granules. Vesicle moderately elongated with a pear-like shape, smooth; aculeus short and weakly curved. Pedipalps not elongated; femur with five carinae; internal with minute spinoid granules. Patella with inconspicuous carinae. Chela weakly enlarged and with 7–8 inconspicuous carinae. Tegument weakly granular to smooth. Fixed and movable fingers shorter than manus, with 6–7 rows of granulations on the dentate margins. Chelicerae characteristic of the family Chaerilidae [10]. Trichobothriotaxy of type B; orthobothriotaxic [8]; femur with 9 trichobothria, patella with 14, and chela with 14. Legs with pedal spurs moderately developed. Tarsi with two rows of setae. Hemispermatophore unknown.
Morphometric values (in mm) of the male holotype. Total length (including telson), 17.1. Carapace: length, 2.4; anterior width, 1.5; posterior width, 2.7. Mesosoma length, 5.4. Metasomal segments. I: length, 0.7; width, 1.5; II: length, 1.0; width, 1.2; III: length, 1.1; width, 1.1; IV: length, 1.4; width, 1.1; V: length, 2.3; width, 1.0; depth, 0.9. Telson length, 2.8. Vesicle: width, 1.1; depth, 0.9. Pedipalp: femur length, 1.8, width, 1.0; patella length, 2.0, width, 0.9; chela length, 4.1, width, 1.3, depth, 1.4; movable finger length, 2.1.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgements
We are most grateful to Michael M. Webber, University of Nevada, Las Vegas for her review of an earlier version of the manuscript and to Elise-Anne Leguin (MNHN, Paris) for her assistance with the preparation of the photos and plates.