

1 Introduction
Pesticides are synthetic molecules aimed at being toxic towards fungi, plants or animals that are detrimental to cultures. Fungicides, herbicides, and insecticides have been developed in order to control as specifically as possible these pests in order to protect cultures. Nevertheless, these pesticides can be toxic to Human and wild fauna. These molecules are intended to be toxic since they are aimed to destroy living organisms, but they are selected to affect precise steps in target organism(s) that are not present in non-target organisms. Also, their intended use is to acutely eradicate unwanted species, and most toxicity tests are also done in acute situations rather than in long-term experiments. And it is generally in long-term situations that xenogeneic molecules can potentially act as endocrine disruptors.
Our aim in the present paper is to define the functional characteristics of endocrine disruptors in order to evaluate if their toxicity toward non-target species is primarily due to endocrine disruption or not.
2 Endocrine disruptors definition(s)
Endocrine disruptor compounds (EDCs) have been defined in 2002 by the WHO: “An endocrine disruptor is an exogenous substance or mixture that alters function(s) of the endocrine system and consequently causes adverse health effects in an intact organism, or its progeny, or (sub)populations”.
The endocrine system is constituted by a large network of hormones allowing the coordinate functions of dozens of different cell types in multicellular organisms. This network possesses numerous loops of stimulation and retroaction in cascade so that the different physiological parameters (such as glycaemia, lipedema, hydro-mineral balances, etc.) and physiological functions (such as development, growth, reproduction, etc.) are set in the proper range for the good health of the whole organism and for the survival of the species.
In this paper, our aim is to introduce a number of central questions concerning EDCs with particular interest in pesticides.
3 Endocrine disruption mechanisms
EDCs are molecules, natural or synthetic, that happen to interfere with the endocrine network of vertebrates, provoking adverse dysregulation of the hormonally controlled physiological parameters or functions [1]. This interference can occur through different mechanisms (Fig. 1) either directly by binding to the hormone receptor, or indirectly by increasing or decreasing the concentration of endogenous hormone(s):
- • the most direct one is an interaction of the EDCs with an hormone receptor leading either to stimulation (Fig. 1A) [2] or inhibition (Fig. 1A) of downstream cellular pathway in target cells;
- • or endogenous active hormone concentrations can be affected by the stimulation or inhibition of either their synthesis (Fig. 1B), or degradation (Fig. 1C) [2,3], or availability (Fig. 1D).
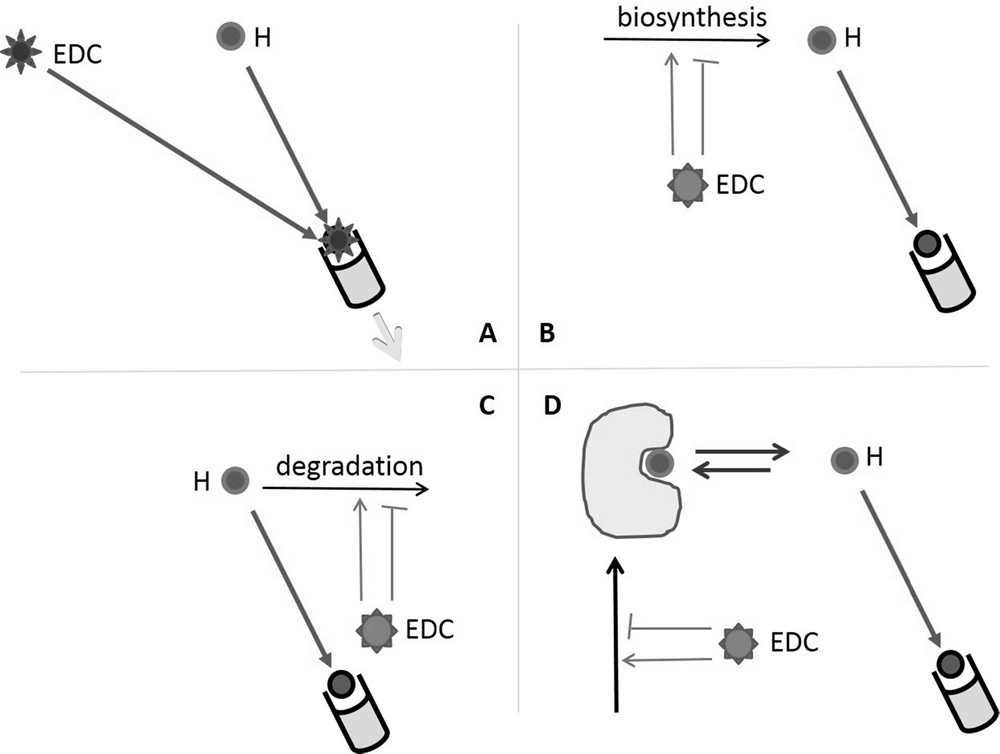
Mechanisms of action of Endocrine Disruptor Compounds (EDCs). A. Direct interaction of EDC with a hormone nuclear receptor leading to stimulation (agonism) or inhibition (antagonism) of its transcriptional activity. B. Stimulation or inhibition of endogeneous hormone biosynthesis. C. Stimulation or inhibition of endogeneous hormone degradation. D. Stimulation or inhibition of endogeneous hormone binding protein leading to decreased or enhanced circulating hormone availability.
A number of in vitro and in vivo tests have been set up by diverse national and international agencies to identify EDCs [4–7].
4 Structures and intended toxic activity of pesticides
A large majority of pesticides are small organic molecules with molecular weights around 300 to 2000 Da.
The insecticides control insects by interfering with their nervous system or by inhibiting their molt. For example, acetylcholinesterase inhibitors (organophosphates, carbamates) and sodium channel agonists (pyrethroids) act on the insect's nervous system [8,9] as well as neonicotinoid insecticides (imidacloprid, acetamiprid, thiacloprid, clothianidin, thiamethoxam, and dinotefuran) that act through their preferential affinity for nicotinic receptor (nAChR) subtypes [10–15]. The latter have favourable safety profiles, due to their poor penetration of the mammalian blood–brain barrier and low application rates, and they effectively control pest species that have evolved resistance to other insecticide classes. However, due to their high intrinsic toxicity to honey bees [16,17], nitro-substituted neonicotinoid insecticides have been intensively examined worldwide by regulatory agencies and temporarily suspended in the European Union for seed treatment, soil application, and foliar treatment in crops attractive to bees. This illustrates the difficulty to develop and use insecticides without affecting non-target insects [18,19].
The herbicides control development of unwanted plants by inhibiting synthesis of some of their amino-acids or their photosynthesis or by specifically antagonizing the action of natural regulators of their development [20–22]. For example, triketone herbicides alter the formation of carotenoids and therefore disrupt the photosynthetic electron transport in plants. Since the aim is to have physiological impact on specific weeds and not on crops, the rapid metabolism of triketones by maize, in particular by cytochrome P450 enzymes, makes this plant insensitive to the herbicide treatment [23]. A precise knowledge of the weed species is important to obtain maximum efficiency by choosing the most specific and efficient herbicide [24]. The fungicides [25] control fungi by inhibiting the synthesis of some of their amino-acids or their cell division.
5 Structures and endocrine-disrupting activity of pesticides
Many in vitro tests have been set up to detect direct potential stimulating or inhibiting effects (Fig. 1A) of pesticides on the transcriptional activity of nuclear receptors, that are the main direct targets of EDCs [26]. In 2010, a large survey of 200 pesticides in in vitro transcriptional tests for six different receptors (hERα, hERβ, hAR, hPXR, mPPARα, mAhR) was published [27], and pointed out that, among them, 47 showed hERα-mediated action and 33 had hERβ-mediated action. Only three showed mPPARα-mediated transcription, whereas 11 had mAhR-mediated activity. Interestingly, as much as 106 out of 200 exhibited agonist activity in hPXR-dependent systems. It is interesting to point out that the PXR is the nuclear receptor with the most voluminous binding pocket (≥ 1200 Å3), thus susceptible to accommodate a larger number of molecules, and larger molecules.
It is much more difficult to set up in vitro tests corresponding to indirect mechanisms (Fig. 1B, C and D) as they can be very diverse.
6 How endocrine disruptors differ from hormones?
Hormones and their receptors have co-evolved as to ensure proper specificity of hormone–receptor pairs in all generations of evolving and diversifying species. The natural ligands for nuclear receptors are not polypeptide or protein hormones and, therefore, they are not directly encoded by genes. It can also be observed that the hormones binding to nuclear receptors are very well conserved during evolution (estrogens, progestagens, thyroid hormones, etc.). It can thus be concluded that the evolution of nuclear receptors has been restricted by the need for recognizing its cognate hormone and also, by the need not to bind another circulating molecule. It can be hypothesized that an animal with a receptor allowing its stimulation by a wrong circulating ligand would not survive or would not be able to reproduce (which is the same in terms of Evolution).
The problem with the arrival of thousands of small industrial organic molecules is that the nuclear receptors have not been selected during Evolution to avoid their low-affinity binding. Therefore, a certain percentage of these molecules can, more or less, accommodate the receptors binding site and promote receptor activation or inactivation; i.e. these molecules act as endocrine disruptors.
Moreover, conjugating and degrading systems have also been set up during Evolution to limit the half-life of hormones, whereas synthetic molecules can remain unchanged and potentially accumulate in the body.
7 Hazard versus risk of endocrine-disrupting activity of pesticides
As for all other forms of toxicity, it is important to consider both the potential hazard of all suspected molecules as thoroughly as possible in order to evaluate the risk for human and/or wildlife populations. In this prospect, it is important to consider (1) the exposure of populations to the chemical under study, (2) the dose responses of its effects, and (3) the cocktail effect.
In the case of pesticides, it is clear that professional exposure [28] is considerably higher than the general population's exposure, but professionals can take appropriate measures, if correctly informed, while the public should not be exposed to detrimental levels of pesticide(s). Moreover, the prenatal and postnatal periods of development until puberty are particularly sensitive, because of their persistent effects due to possible detrimental effects during developmental steps [29]. The exposure of the general population to persistent organic pollutants, including pesticides, can be followed by the detection of their presence in body fluids or tissues. Due to the extreme sensitivity of most modern assays, it is necessary to consider, not solely the presence of these molecules, but their actual concentration.
As for all other forms of toxicity, the potency of potential endocrine disruptors is important information to consider, and the risk is generally dependent on both exposure and potency.
Non-monotonous dose responses are sometimes claimed as a hallmark of the effects of endocrine disruptors. The observation, in some experiments, of U-shape dose-response curves where low doses of chemical exhibit higher effect than medium doses, and comparable effect with high doses cannot be attributed to intrinsic properties of endocrine disruptors. Indeed, this is rather due to the complexity of endocrine regulations in animals and depends on the chosen end-point for the study of the effect(s). The EDCs exhibit low affinity and low specificity towards their target proteins. Indeed, they generally bind to one or several proteins (receptors, binding proteins, conjugating enzymes, etc.) susceptible to affect the animals’ endocrine system. Their positive and negative effects at different sites can lead to non-monotonous responses, possibly different depending on the endpoint chosen. Moreover, the opinion that it is a property of EDCs to exhibit higher activity at lower doses than at higher doses could lead to the erroneous simplistic thought that it would better to always have a certain level of EDCs in the environment to avoid low-dose effects!
It is clear that the number of potentially toxic molecules in the environment leads to take into consideration the possibly additive, or worse, synergic effects of EDCs. Owing to the complexity of endocrine regulations, such effects would require to study EDCs two by two, or more. It is of course an impossible task, and rather representative molecules with given mechanisms should be studied for additive, synergic or counteracting effects with representative molecules of other groups to detect the most likely cocktail effects.
8 Conclusion and perspectives
Endocrine disruption is a mild, but insidious, form of toxicity so that it is often difficult to prove or reject its involvement. The complexity of endocrine regulations in vertebrates together with the high number of new synthetic molecules, including pesticides, render the task of determining the endocrine-disrupting risk for each molecule, enormous, and often contradictory.
For these reasons, a classification of molecules under study per mechanism(s) of action on endocrine regulations would be helpful to reduce the number of assays needed to validate endocrine disruption and also to take into account potential additive [30,31], synergistic or counteracting cocktail effects [31,32].