

1 Introduction
The genus Discocyrtanus Roewer, 1929 is currently composed by five valid species, distributed in Central (Cerrado) and Eastern (Atlantic Forest) regions of Brazil, and they are characterized as medium to large gonyleptids, dull and dark colored, with serial armature on femur IV and powerful armature on the ocularium [1]. It has been under synonymy of Discocyrtus Holmberg, 1878 since 1944 [2], when B. Soares considered that the pair of paramedian tubercles present in area I could bring ambiguity to the interpretation of the classification at the genus level according to the Roewerian system. Today, after the revalidation of the genus [1], the taxon began to disassociate from this purely operational paradigm. By means of phylogenetic studies, it was transferred of subfamily leaving the Pachylinae Sørensen, 1884 (highly corroborated as polyphyletic) [3–5] and integrating a subfamily recently proposed by Carvalho and Kury [6]. In this work, we describe a new species of Discocyrtanus (Fig. 1) and provide a key to the species of the genus.

Discocyrtanus canjinjim sp. n., specimens in vivo. A–D. Male, from Vila Bela da Santíssima Trindade, Mato Grosso, Brazil. E–F. Female, same locality.
Photos by A.B. Kury.
2 Material and methods
The descriptions of colors use the standard names followed by the centroid code in parenthesis of the 267 Color Centroids of the NBS/IBCC Color System [7] as described in Kury and Orrico [8]. The formula for the tarsomere count in which the distitarsi of legs I and II are indicated between parentheses was created by Avram [9]. The formula for megaspines of pedipalpus in which I = large spine and i = small spine is shown as used by Kury [10]; here, the same formula is used to discriminate the size of spines in Femur IV. The terminology for the scutum outline follows Kury and Medrano [11]. The terminology for chaetotaxy of the ventral plate of the penis follows Kury and Villarreal [12] for the macrosetae and Kury [13] for the microsetae. The term mesotergum [14] refers to the roughly subrectangular region of dorsal scutum formed by areas I to IV and circumscribed anteriorly by the carapace, laterally by the lateral margins and posteriorly by the area V (posterior margin of scutum). The term dry mark (“mancha seca”) Kury [15] (called “dry-mark” Kury in Da Silva & Gnaspini [16]) refers to the more external serose layer of the cuticle, which forms powdery patterns in the living animal or when it dries up after having been removed from the preserving fluid.
Biogeographical units used here are from the WWF Terrestrial Eco-regions of the World (names starting with “NT” [17]). They are indicated by colored background areas on the map (Fig. 2).

Map of the Mato Grosso state, central-western Brazil, showing the distribution of Discocyrtanus canjinjim sp. n. and Discocyrtanus bugre Kury & Carvalho, 2016. The shaded areas on the background are WWF terrestrial eco-regions: NT 0212 (Chiquitano dry forests, in pink) and NT 0704 (Cerrado, in orange). For the geographic distribution of all four other species of Discocyrtanus, see [1], page 132, fig. 4.
Scanning Electron Microscopy was carried out with a JEOL JSM-6390LV at the Center for Scanning Electron Microscopy of the Museu Nacional/UFRJ. All measurements are in mm.
The diagnoses given here are comparative among the five species of Discocyrtanus: Discocyrtanus bugre Kury & Carvalho, 2016, Discocyrtanus goyazius Roewer, 1929, Discocyrtanus oliverioi (H. Soares, 1945), Discocyrtanus pertenuis (Mello-Leitão, 1935) and Discocyrtanus tocantinensis Kury & Carvalho, 2016.
Abbreviation of the repositories cited are IBSP (Instituto Butantan, São Paulo) and MNRJ (Museu Nacional, Rio de Janeiro). Abbreviation of relevant Brazilian state is MT = Mato Grosso. Other abbreviations used: AS = abdominal scutum, CL = carapace length, CW = carapace width, AL = abdominal scutum length, AW = abdominal scutum width (body); Tr = trochanter, Fe = femur, Pa = patella, Ti = tibia, Mt = metatarsus, Ta = tarsus (appendages); VP = ventral plate, A1–A3 = basal macrosetae of VP, B = ventro-basal macrosetae of VP, C1–C3 = distal macrosetae of VP, D1 = dorso-lateral subdistal small seta of VP (penis).
Tarsal formula: numbers of tarsomeres in tarsus I to IV (Table 2), when an individual count is given, order is from left to right side (figures in parentheses denote number of tarsomeres only in the distitarsi I–II).
Right/Left tarsal (disititarsal) counts of Discocyrtanus canjinjim sp. n., male holotype and paratypes.
| ♂ (holotype) | ♂ (n = 7) | ♀ (n = 6) | |
| Leg I | 6 (3)/6 (3) | 6 (3)/6 (3) | 6 (3)–6 (3) |
| Leg II | 9 (3)/9 (3) | 9–10 (3)/9–10 (3) | 9–10 (3)/9–10 (3) |
| Leg III | 7/7 | 7/7 | 7/7 |
| Leg IV | 7/7 | 7/6–7 | 7/7 |
3 Systematic accounts
Discocyrtanus canjinjim sp. n.
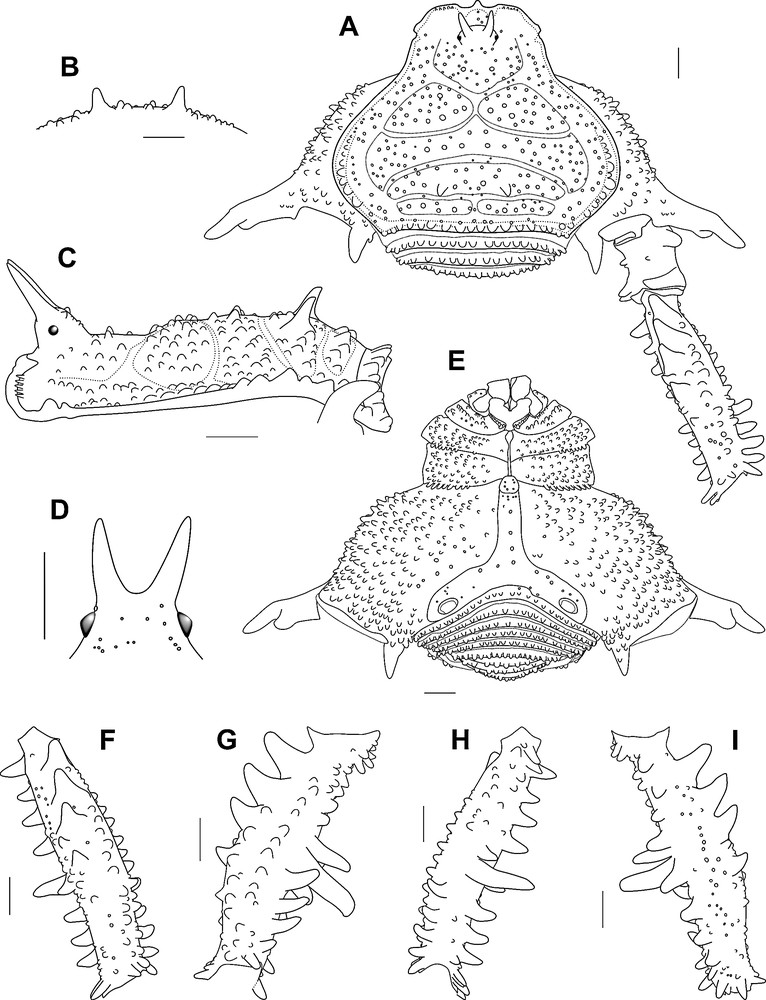
Discocyrtanus canjinjim sp. n., ♂ holotype (MNRJ 2550): A. Habitus, dorsal view. B. Armature of scutal area III, dorso-posterior view. C. Habitus, lateral view. D. Ocularium, frontal view. E. Habitus, ventral view. F. Right femur IV, dorsal view. G. Same, prolateral view. H. Same, ventral view. I. Same, retrolateral view. Scale bars = 1 mm.
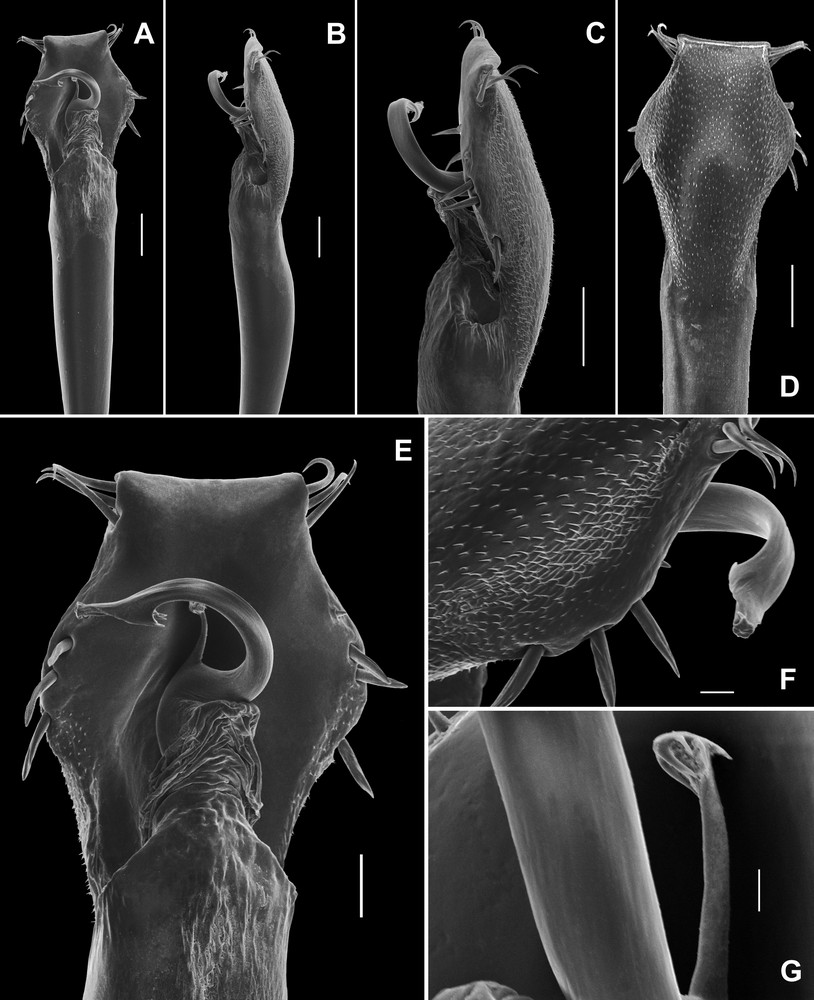
Discocyrtanus canjinjim sp. n. (MNRJ 2547); ♂ genitalia, distal part. A. Dorsal view, panoramic. B. Lateral view, panoramic. C. Lateral view. D. Ventral view. E. Dorsal view. F. Ventro-lateral view, showing the apex of the stylus. G. Ventral process of stylus, dorso-lateral view. Scale bars: 100 μm (A–D), 50 μm (E), 20 μm (F), 10 μm (G).

Discocyrtanus canjinjim sp. n. (MNRJ 2549); ♂, minor form. A. Habitus, dorsal view. B. Habitus, ventral view. C. Right femur IV, prodorsal view. D. Left trochanter, femur and patella IV, dorsal view. E. Right femur, patella and tibia IV, prolateral view. Scale bars = 1 mm.
Type data. ♂ Holotype (MNRJ 2550), Brazil, MT, Vila Bela da Santíssima Trindade, –14.93851, –60.00940, 14.xi.2015, Chagas Jr, A. & Kury, A. B. leg.; 1 ♂ 1 ♀ paratypes (IBSP 10423) BRASIL, MT, Vale de São Domingos, Rio Guaporé, UHE Guaporé, 2002, Equipe IBSP leg.; 1 ♂ paratype (IBSP 10432) BRASIL, MT, Vale de São Domingos, Rio Guaporé, UHE Guaporé, 2002, Equipe IBSP leg.; 1 ♂ 1 ♀ paratypes (IBSP 10438) BRASIL, MT, Vale de São Domingos, Rio Guaporé, UHE Guaporé, 2002, Equipe IBSP leg.; 1 ♂ 1 ♀ paratypes (MNRJ 2547), Brazil, MT, Vila Bela da Santíssima Trindade, –14.93851, –60.00940, 14.xi.2015, Chagas Jr, A. & Kury, A. B. leg.; 3 ♂ 2 ♀ paratypes (MNRJ 2548), Brazil, MT, Vila Bela da Santíssima Trindade, –14.93851, –60.00940, 14.xi.2015, Chagas Jr, A. & Kury, A. B. leg.; 3 ♂ 3 ♀ paratypes (MNRJ 2549), Brazil, MT, Vila Bela da Santíssima Trindade, -14.93851, –60.00940, 14.xi.2015, Chagas Jr, A. & Kury, A. B. leg.
Etymology. The specific name is a noun in apposition referring to canjinjim, a traditional spiced-honeyed alcoholic beverage of African origin, typical of Vila Bela da Santíssima Trindade, which originated from a safe haven for escaped slaves.
Diagnosis. Area III with a pair of paramedian rounded acuminate high tubercles (higher than in D. goyazius, D. oliverioi and D. pertenuis and sharper than in D. tocantinensis), slightly curved backwards (Fig. 3B–C). Area IV with sinuous row of six rounded tubercles (transverse row of three rounded tubercles occurring in D. bugre, D. goyazius, D. oliverioi and D. pertenuis) (Fig. 3A, C). Tr IV of ♂ with prodorsal distal apophysis as a stout hook, curved to retrolateral portion (undeveloped in D. tocantinensis, and not bifurcated, with proximal branch reduced as in D. goyazius, D. oliverioi and D. pertenuis) (Fig. 3A). Fe IV shorter and more swollen in comparison to other Discocyrtanus species (Fig. 3A, F–I). Fe IV with dorsal proximal–medial row of five spines (IIIii) (not present in D. oliverioi and D. tocantinensis, and larger and wider in comparison to D. bugre, D. goyazius and D. pertenuis) (Fig. 3A, F, G, I). Fe IV with row of three spines in the proventral distal surface (III) (Fig. 3G, H). Fe IV with a row of nine spines from the retroventral proximal to the distal axis (iiiIIIiii) (Fig. 3H, I). Ventral plate of penis with distal portion forming a trapeze, differently of the rectangular shape of the others Discocyrtanus (Fig. 4A, D, E). Ventral process of stylus half as long as stylus and gently curved, which does not occur in other Discocyrtanus species (Fig. 4A, E, G).
Distribution. BRAZIL: MT, Vale de São Domingos, Vila Bela da Santíssima Trindade.
Description. Male holotype. Measurements. CW 4.8, CL 2.8, AW 9.7, AL 4.8. Leg measurements on Table 1, tarsal counts on Table 2.
Leg measurements of Discocyrtanus canjinjim sp. n., male holotype (MNRJ 2550).
| Tr | Fe | Pa | Ti | Mt | Ta | Claw | Total | |
| Pp | 0.89 | 1.45 | 1.05 | 1.18 | − | 1.00 | 0.45 | 6.02 |
| Leg I | 0.77 | 2.82 | 1.27 | 2.63 | 3.82 | 1.82 | − | 13.13 |
| Leg II | 1.05 | 6.82 | 1.73 | 6.00 | 6.91 | 4.82 | − | 27.33 |
| Leg III | 1.23 | 5.82 | 1.91 | 3.81 | 5.09 | 2.54 | − | 20.40 |
| Leg IV | 2.27 | 6.36 | 2.36 | 6.13 | 9.27 | 3.00 | − | 29.39 |
Dorsum. Dorsal scutum, almost as long as wide, abdominal scutum with lateral margins strongly convex, widest at area II and highest at area III (Fig. 3A, C). Carapace with a posterior pair of paramedian higher tubercles, with several ordinary tubercles around them (Fig. 3A, C). Cheliceral sockets shallow, with a small apophysis in the center (Fig. 3A). Ocularium elliptical, high, inclined frontwards, placed in the anterior portion of the carapace, armed with a pair of divergent high spines fused at baseline and inclined frontwards (Fig. 3A, C, D). Mesotergum divided into four clearly defined areas (Fig. 3A, C). Areas I and IV divided into left and right halves by median groove. Area II anterior lateral border invading space of area I and posterior lateral border invading the space of area III. AS lateral borders with ordinary tubercles from area II backwards. All areas with many tubercles, area I with a pair of paramedian tubercles a little higher than the others (Fig. 3A). Area III with a pair of paramedian rounded sharped higher tubercles slightly curved backwards (Fig. 3A–C). Area IV with sinuous row of six rounded tubercles. Posterior border of dorsal scutum and free tergites with a horizontal row of larger rounded and acuminated tubercles (Fig. 3A).
Venter. Cx I–III parallel to each other; each with several ventral transverse rows of 9–11 setiferous tubercles (Cx I main row with higher and sharper tubercles). Cx II retroventral distal with a row of seven acuminated tubercles. Cx III retroventral distal with a row of eight acuminated tubercles (Fig. 3E). Cx IV much larger than the others, directed obliquely. Stigmatic area Y-shaped, clearly sunken relative to distal part of coxa IV. Intercoxal bridges well-marked. Stigmata clearly visible. Free sternites and anal operculum each with one transverse row of tubercles (Fig. 3E).
Chelicera. Basichelicerite elongate, bulla well-marked, with marginal setiferous tubercles – three ectal, three posterior, three mesal; hand not swollen (Fig. 5A).
Pedipalpus. Tr with two geminated ventral setiferous tubercles. Fe with a prolateral apical setiferous tubercle and one ventral basal setiferous tubercle. Pa unarmed. Ti with two rows of setiferous tubercles; four (IiIi) ventro-mesal and three (IiI) ventro-ectal. Ta with two rows of setiferous tubercles; three (III) ventro-mesal and four (IiIi) ventro-ectal.
Legs. Cx I–III each with ventral transverse rows of 9–11 setiferous tubercles (Fig. 3E). Tr I–III each with several ventral tubercles. Fe I–II straight. Fe and Ti I–II with two rows (proventral and retroventral) of small tubercles. Fe II with a little retrodorsal distal spur. Leg III sub-straight. Fe III and Ti III with two rows (proventral and retroventral) of acuminated tubercles. Fe III with a developed proventral (geminated) and retrodorsal distal spore. Cx IV ending distally at area IV of dorsal scutum (Fig. 3A). Cx IV with (1) a thick falciform apical prolateral apophysis, moderately elongate, with small accessory blunt branch and (2) a retroventral spiniform apophysis with secondary branch (Fig. 3A, E). Cx IV prodorsal, prolateral, proventral and ventral with rows of acuminated tubercles (Fig. 3A, E). Tr IV retrolateral with row of three spines (iiI). Tr IV retrodorsal distal with a spine (I). Tr IV apophysis prolateral proximal convex. Tr IV with prodorsal distal apophysis, forming a stout hook (Fig. 3A). Tr IV ventrally covered by tubercles along its entire length. Fe IV curved from the proximal-distal region toward dorsal, swollen in the proximal–medial part (Fig. 3G, I). Fe IV with row of nine spines from the retroventral proximal to distal axis (iiiIIIiii), ending with a well-developed spur (Fig. 3H, I). Fe IV ventral medial with two spines (iI), the distal curved to the retrolateral portion (Fig. 3H). Fe IV with row of three spines in the proventral distal surface (III) (Fig. 3G, H). Fe IV prodorsal, prolateral and retrodorsal with row of rounded tubercles. Fe IV dorsal proximal–medial with row of five spines (IIIii) (Fig. 3F, G, I). Pa IV proventral and retroventral with row of three spines. Pa IV retrolateral distal with a spine. Ti IV prodorsal, proventral, retrodorsal and retroventral with row of acuminate tubercles. Mt IV proventral and retroventral distal with spur.
Penis. VP divided into two regions: distal part trapezoidal, proximal part elliptical (Fig. 4A, D, E). Ventral surface of VP entirely covered with microsetae of the type 1 (Fig. 4D). All macrosetae inserted on lateral of VP: A1–A3, cylindrical, thick, acuminate, forming a triangle on basal third of VP (Fig. 4A–E); B inserted ventrally, proximal to A3 (Fig. 4C, F); C1–C3 slender, only moderately elongate, forming a tight row on the distal part of VP (Fig. 4A–E); D1 small, midway between C3 and A1 (Fig. 4C, E, F); E1–E2 inserted on distal lateral border of VP, E1 between C1 and C2, E2 proximal to C3. Glans sac short, arising from middle bulge on podium, not extended as a dorsal process (Fig. 4A–C, E). Stylus stout, cylindrical and S-shaped curved to dorsal (Fig. 4A, E). Apex of stylus flattened dorsoventrally and spiny (Fig. 4C, E, F). Ventral process of stylus gently curved with half the longitudinal length of the stylus (Fig. 4A, E, G). Apex of ventral process of stylus forming a small flabellum with flaps deeply serrated (Fig. 4G).
Color (in vivo) (Fig. 1). Dorsal scutum background Deep Reddish Brown (41) with darker shading especially in lateral areas and carapace. Smaller granules of scutum with dry marks Grayish Yellowish Pink (32), main paramedian tubercles of area III are dark as the scutum. Scutal grooves lighter, Grayish Reddish Orange (39). Articular membranes White (263). Chelicerae and pedipalps Deep Yellowish Brown (75). Legs I to III Grayish Yellowish Brown (80) with darker reticle. Leg IV Dark Grayish Reddish Brown (47), except for tips of larger apophyses of Cx, Tr and Fe, which are Deep Reddish Orange (36) and the rest of Cx–Tr which are like the dorsal scutum.
Color (in alcohol) (Fig. 5). Dorsal scutum background deep orange yellow (69) with darker shading especially in lateral areas and carapace. Smaller granules of scutum, tips of main paramedian tubercles of area III and larger rounded tubercles of lateral area Strong Yellowish Brown (74). Articular membranes White (263). Chelicerae and pedipalps background Vivid Greenish Yellow (97) with dense dark Yellowish Brown (78) honeycombed reticle. Legs I to III Moderate Greenish Yellow (102) with darker reticle, especially so in leg III. Leg IV Moderate Olive Brown (95), except for tips of larger apophyses of Cx, Tr and Fe, which are Strong Yellowish Brown (74) and the tips of proventral and retroventral rows of spines, which are Moderate Greenish Yellow (102).
Female paratype (MNRJ 2549): CW 4.8, CL 3.2; AW 9.2, AL 5.0. Side of the dorsal scutum edges with lower level of concavity compared to male (Fig. 1E, F). Cx IV with acuminate prodorsal apophysis. Fe IV thinner and less curve when compared to male. No spines on Fe IV, only the spore retrolateral distal (Fig. 1E, F). No acuminate tubercles Ti IV.
Minor forms of males (MNRJ 2549): CW 4.5, CL 2.8; AW 7.7, AL 4.2. Side of the dorsal scutum edges with concavity level between those found in major forms and females (proportionally to the dorsal scutum) (Fig. 5A, B). Cx IV with prolateral apophysis apical without developed second branch (Fig. 5B). Fe IV thinner and less armed when compared to major form (Fig. 5C–E).
4 Discussion
The sympatric distribution of species of Discocyrtanus is shown in [1], where one can see that there is overlap in areas of occurrence of Discocyrtanus goyazius Roewer, 1929, and Discocyrtanus pertenuis Mello-Leitão, 1935, on the border region between Mato Grosso do Sul and São Paulo, where a transition zone occurs between NT 0704 (Cerrado) and NT 0150 (Alto Paraná Atlantic Forests). In the case of the allopatry of Discocyrtanus canjinjim sp. n. and Discocyrtanus bugre Kury & Carvalho, 2016 (Fig. 2), the second has its distribution on Cerrado and the borders of NT 0212 (Chiquitano Dry Forests), where the first is endemic from NT 0212. Due to the lack of fieldwork in the region west to Vila Bela da Santíssima Trindade area, it is not known whether this species could extend to central-eastern Bolivia, where NT 0212 is present. If such occurrence is confirmed in the future, it will be the first case of species of Discocyrtanus outside Brazil.
The new species described here, D. canjinjim, has a new form of genitalia to the genus, showing a ventral process of stylus and a distal part of VP trapezoidal. This composition differs from the uniform shape presented by all the five other species of Discocyrtanus, as noted in [1]. However, the genitalia, along with the pattern of dorsal scutum and the Fe IV armature, indicate the presence of notable synapomorphies of this genus, as pointed out by Carvalho and Kury [6].
5 Key to males of Discocyrtanus species
| 1. Free tergites with a row of acuminate tubercles, Fe IV prolaterally convex, Tr IV rectangular, Glans sac without ventral process | 2 |
| 1′. Free tergites with a row of rounded tubercles; Fe IV straight, with swelling in the medial region; Tr IV quadrate; Glans sac with ventral process (Figs. 3A, C; 5A, E, G) | Discocyrtanus canjinjim sp. n. |
| 2. Area III of mesotergum without median groove, with paramedian acuminated tubercles; Tr IV prodorsal distal apophysis forming a hook | 3 |
| 2′. Area III of mesotergum with median groove, with paramedian rounded tubercles; Tr IV prodorsal distal apophysis not forming a hook ([1] fig. 10A, B) | Discocyrtanus tocantinensis Kury & Carvalho, 2016 |
| 3. Divergent armature of ocularium, Tr IV prodorsal distal apophysis without developed second branch, Fe IV with spines on proximal dorsal portion | 4 |
| 3′. Parallel armature of ocularium, Tr IV prodorsal distal apophysis without developed second branch (one-third the size of the main branch), Fe IV without spines on proximal dorsal portion ([18] Fig. 3A) | Discocyrtanus oliverioi (H. Soares, 1945) |
| 4. Area IV with ordinary tubercles only; Fe IV without armature of spatulate form; Fe IV proventral medial–distal with row of 10 or more spines | 5 |
| 4′. Area IV with transversal row, on both sides, of three conspicuous tubercles in relation to the others; Fe IV retrolateral proximal with two spatulated spines; Fe IV proventral medial–distal with row of seven spines ([1] Fig. 5A, D–G) | Discocyrtanus bugre Kury & Carvalho, 2016 |
| 5. Abdominal scutum flattened dorsoventrally; Fe IV with same diameter of the Fe III distal portion; Fe IV dorsal proximal–medial with five tiny spines ([19] fig. 31) | Discocyrtanus goyazius Roewer, 1929 |
| 5′. Abdominal scutum dorsally convex; Fe IV with a diameter greater than that found on Fe III; Fe IV dorsal proximal–medial with four conspicuous spines ([20] Fig. 1) | Discocyrtanus pertenuis (Mello-Leitão, 1935) |
Disclosure of interest
The authors declare that they have no competing interest.
Acknowledgements
This study has been supported by grant #306411/2015-6 from the “Conselho Nacional de Desenvolvimento Científico e Tecnológico” (CNPq) to ABK and scholarship # 134421/2016-7 (CNPq) to RNC. The SEM micrographs were taken in the Center for Scanning Electron Microscopy of the Museu Nacional/UFRJ (financed by PETROBRAS), with the kind assistance of Beatriz Cordeiro. We thank two anonymous referees for welcome suggestions to the draft.