

1 Introduction
The family Chaerilidae, with its type genus Chaerilus Simon, 1877, was the subject of numerous publications in recent years, but remains yet poorly-known among extant scorpions; for some historical details, see Lourenço and Lourenço and Pham [1–3] Recently a second genus within the family, Chaerilourencous Rossi, 2018, was proposed for accommodating two peculiar anophthalmus species [4]. Some recent attempts to full revise the taxonomy of the genus were far from being successful [5,6], and brought mainly supplementary confusion to the status of several species. These attempts of revision were followed by an increasing number of publications on the Chaerilidae with the description of numerous new species. Some of these publications were properly constructed and illustrated [1–3,7–10], whereas others were unequally illustrated and worse, constructed without any historical introductions and/or methods [5,11–13]. This absence of introductions and methods apparently indicates a deliberated decision of ignoring previous results.
Although some progress has been made in the study of the Chaerilus species with clarification of the status of some populations [14,15], the precise identity of some old or recently described species remain yet unclear. The classification of the Chaerilus species in natural groups was attempted [1,2], but without a definitive conclusion. This is mainly because several species deposited in private collections could not be examined. Many private collections are in fact not accessible to Academic Experts. Consequently, only a global and detailed revision of this genus will bring answers to a number of existent gaps, but this is not the aim of this publication.
Among the polemic discussions that took place during the last two decades [16], one concerned the precise status and range of distribution of Chaerilus chapmani Vachon & Lourenço, 1985, a species described from caves in Sarawak (Borneo). The original material of C. chapmani was collected by members of the Royal Geographical Society during two expeditions that took place in the caves of the Gunong Mulu National Park, Sarawak in 1977–1978 and 1980–1981 [17]. A number of scorpions were untrusted to Prof. Max Vachon, in France, by Dr Philip Chapman, but others were also sent to Dr Oscar Francke, in the USA. At least one of these specimens was deposited within the American Museum of Natural History, NY, USA. In the already cited revision done by Kovařík [6], this author refers to C. chapmani based only on specimens from Palawan Island collected in St. Paul National Park, in a cave (?) under rocks. The date of February 1997 is proposed, but no collector is indicated. Concerning precisely the provenance of these specimens, recently Rossi [18] noted that, despite they are now kept in the inaccessible Kovařík's private collection in Prague, the specimens were originally part of the collections of the Museo di Storia Naturale dell’Università di Firenze, Sezione di Zoologia “La Specola”, Italy (MZUF), and were collected by Dr Marco Borri and Dr Cecilia Volpi during an official field trip organized by the Museum in the Philippines, during the year 1991. The specimens were precisely collected on 2 February 1991 in the caves of St. Paul National Park, Puerto Princesa Underground River, in the point of Rockpile (Dr Borri, personal communication). Dr Borri has a diary with precise notes of all his field trips. Several years later, Kovařík borrowed the whole scorpions’ collection of MZUF and kept for his private collection a number of specimens. Among them, there are clearly the Chaerilus specimens from Palawan, which obviously are not present anymore in the Museum's collection. According to the loan register of MZUF, Kovařík borrowed in the year 1997 several lots of scorpions and kept these for his private collection, among many, one specimen from India, one specimen from Somalia, and six specimens from the Philippines. Later Kovařík [6,19] cited these six Chaerilus specimens, reporting the following data: “Philippines, Palawan Island, St. Paul National Park, 2 4 juvs., II.1997, FKCP (the author's collection). All specimens were found under rocks in a cave.” Two points need to be better explained. (1) Although MZUF sometimes can accept, after request, to donate specimens from the Museum's collection to private collections, this is possible only if the requested specimen/specimens are not the unique samples of the species in the collections. In this case, as in additional ones to be explored, it is clear that Kovařík kept for himself, and his inaccessible private collection, all specimens of the rare Chaerilus from the Palawan caves. It is obvious that he did not explain the real situation, when he did the request to the curator who was not an expert in arachnology. (2) It is interesting, and also alarming, to note that Kovařík changed the data of the specimens. According to Dr Borri, we know that the specimens were collected by him and Dr C. Volpi on 2 February 1991, but Kovařík reported that they were collected in February 1997, approximately when he got them on loan. Besides he did not indicate the collectors. Thus a serious doubt can be addressed: are all data published by Kovařík real and trustable? The inevitable answer can only be negative. Moreover the responsibility of co-authors and editors involved in such publications could suggest a form of complicity.
In a note concerning possible biased taxonomic decisions, Lourenço [16] refuted the possible distribution of a true troglobitic species in two islands largely separated by the sea. The presence of C. chapmani out of the Island of Borneo was once again rejected by Lourenço [20] while discussing about the ecology of some cave species of Chaerilus, but based also on the fact that, in his revision, Kovařík [6] had not examined any of the specimens of C. chapmani belonging to the original series. These remarks led immediately to a reply [19], in which this author insisted about the presence of C. chapmani in Palawan, basing his conclusions on two comparative photos of the female specimen of C. chapmani deposited in the American Museum of Natural History, NY, with a female deposited in his private collection. Although this ‘comparative taxonomic analysis’ sounded mostly irrelevant in face of solid ecological arguments associated with cave adaptation, a new re-analysis was not possible in the absence of available material from Palawan. The present study of a relevant number of freshly collected specimens in a cave in Palawan (Fig. 1) leads us to confirm the taxonomic differences between the two populations in Borneo and Palawan, and a new species is described here.
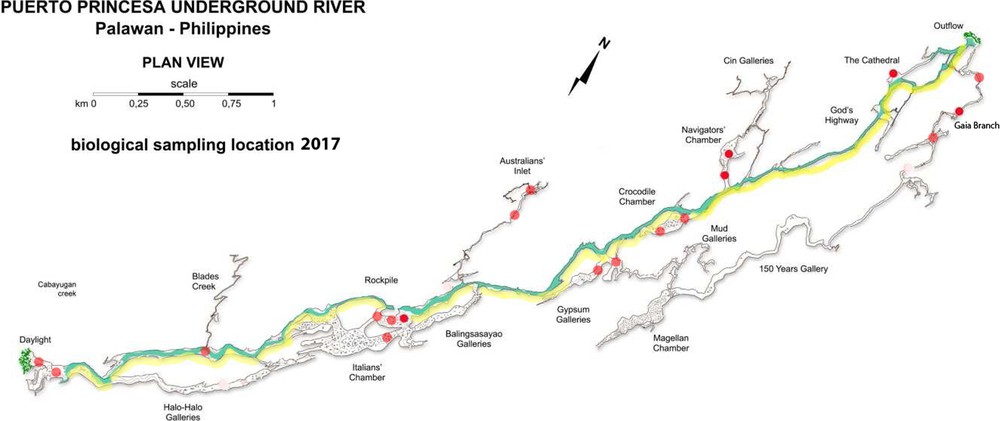
Schematic map of Puerto Princesa Underground River Cave, also known as the caves of ‘Natuturingam’.
2 Methods
Illustrations and measurements were made with the aid of a Wild M5 stereomicroscope that has a drawing tube (camera lucida) and an ocular micrometer. Measurements follow Stahnke [21] and are given in millimeters. Trichobothrial notations follow Vachon [22], and morphological terminology mostly follows Hjelle [23].
3 Taxonomic treatment
Chaerilidae Pocock, 1893
Chaerilus Simon, 1877
Chaerilus chapmani Vachon & Lourenço 1985 (Figs. 2 and 3)
Chaerilus chapmani, male holotype. Habitus, dorsal and ventral aspects.

Chaerilus chapmani, female paratype. Habitus, dorsal and ventral aspects.
This species was described based on 1 male holotype, 1 female allotype, 2 males and 1 female paratypes collected in caves located in the Gunong Mulu National Park in Sarawak (Island of Borneo). The species was properly diagnosed and described with full illustrations. Consequently, a redescription does not appear as necessary. For details about dates and collectors, refer to Vachon & Lourenço [17] (Figs. 2 and 3.
Chaerilus agnellivanniorum sp. n. (Figs. 4–7)

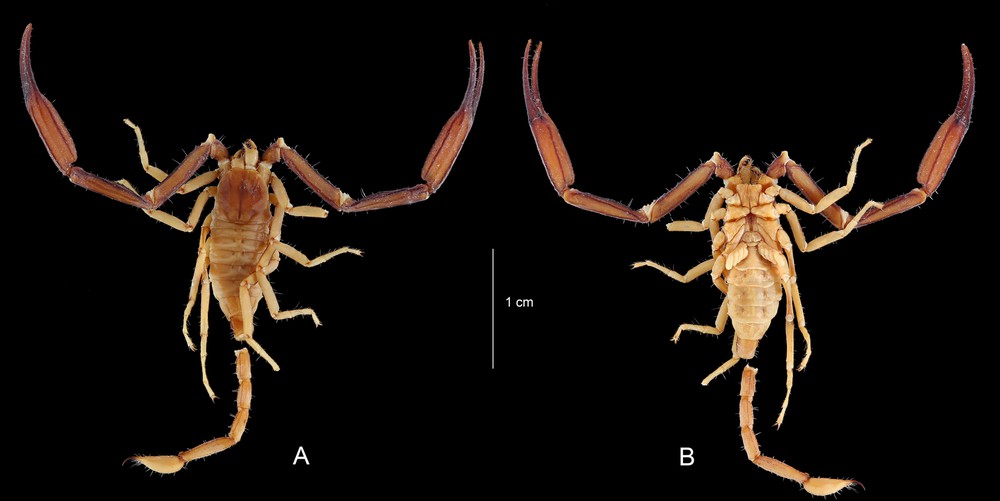
Chaerilus agnellivanniorum sp. n., male holotype. Habitus, dorsal and ventral aspects.

Chaerilus agnellivanniorum sp. n., female paratype. Habitus, dorsal and ventral aspects.
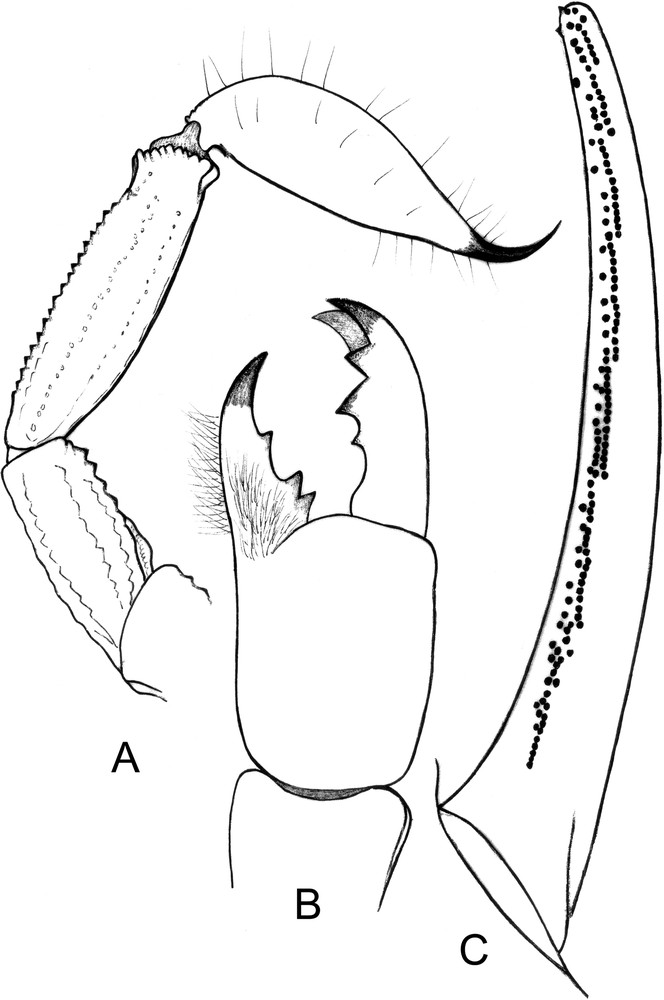
Chaerilus agnellivanniorum sp. n., male holotype. A. Metasomal segments IV-V and telson, lateral aspect. B. Chelicera, dorsal aspect. C. Cutting edge of movable finger with granulations.
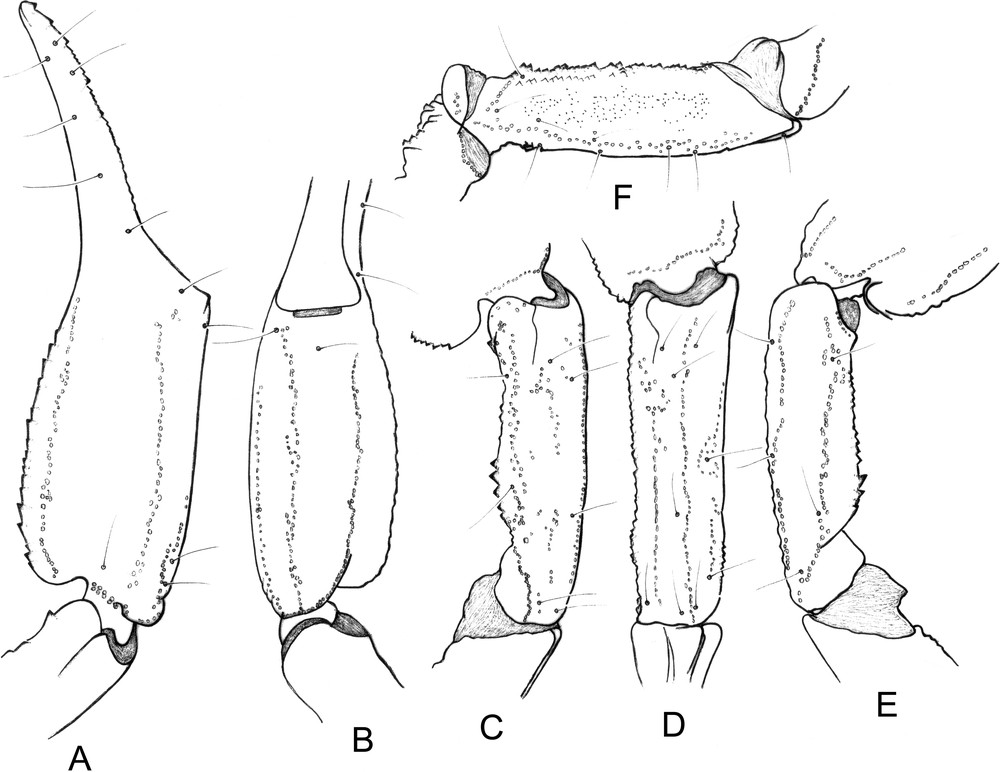
Chaerilus agnellivanniorum sp. n., female paratype. Trichobothrial pattern. A-B. Chela, dorso-external and ventral aspects. C-E. Patella, dorsal, external and ventral aspects. F. Femur, dorsal aspect.
Diagnosis. Species of moderate to small size in relation to the other species of the genus, reaching 36.8 and 40.9 mm in total length, respectively for male and female. Carapace moderately narrowed toward the anterior edge in both sexes; anterior margin weakly emarginated; carinae and granulations moderate to strong, better marked on females; furrows moderately deep in both sexes. Metasomal carinae strong; better marked on females; ventral carinae reduced on segment I, moderately marked on segment II; latero-ventral and ventral carinae on segments IV–V composed of stronger granules that can be moderately spinoid. Dentate margins of fixed and movable fingers of pedipalp chela with 6–7 rows of granules in both sexes, but better defined in males. Pectinal teeth count 4–4 in males and 3–3 in females, without exceptions. Genital operculum plates have an oval to sub-oval shape. Trichobothriotaxy of type B, orthobothriotaxic.
Relationships: Chaerilus agnellivanniorum sp. n. shows several morphological similarities with Chaerilus chapmani described from Sarawak. Both species probably derived from a common ancestral proto-element. The new species can, however, be readily distinguished from C. chapmani by a number of features: (i) different morphometric values, in particular for pedipalp segments that are bulkier, (ii) carination and granulations on carapace, tergites, metasomal segments, and pedipalps clearly better marked, in particular on females (iii) smaller number of pectinal teeth; exclusively four on males and three on females, (iv) median eyes and two pairs of lateral eyes of normal size and never absent, (v) more conspicuous chetotaxy on pedipalps.
Patronym: the name honours Dr Paolo Agnelli (MZUF) and Dr Stefano Vanni (MZUF), who organised the scientific expedition in Palawan, The Philippines.
Type-material: The Philippines, Palawan Island, Puerto Princesa Underground River Cave, Gaia Branch, 6/V/2017 (P. Agnelli & M. Ciaramella), 1 male holotype, 1 male, 1 female juveniles paratypes (MZUF); 28/IV/2017 (Jo De Waele), 1 male, 2 female juveniles paratypes (MZUF); Crocodile Chamber, 7/V/2017 (P. Agnelli, S. Vanni & M. Ciaramella), 1 female juvenile paratype (MZUF); Gypsum Mud Galleries, 26/IV/2017 (P. Agnelli, S. Vanni, M. Ciaramella & C. Paniccia), 1 female paratype (MZUF); 26/IV/2017 (P. Agnelli, S. Vanni & M. Ciaramella), 1 male paratype (ARPC); Italian Chamber, 30/IV/2017 (P. Agnelli), 1 male, 1 female juveniles paratypes (MZUF); 13-14/V/2017 (M. Ciaramella), 1 female, 1 male juvenile paratypes (MNHN); Rockpile, 28/IV/2017 (P. Agnelli, S. Vanni & M. Ciaramella), 1 female paratype (ARPC).
Repositories:
MNHN = Muséum national d’histoire naturelle, Paris, France.
MZUF = Museo di Storia Naturale dell’Università di Firenze, Sezione di Zoologia “La Specola”, Florence, Italy.
ARPC = Andrea Rossi collection, Massa, Italy.
Description based on holotype and paratypes (measurements in Table 1)
Measurements (in mm) of the male holotype and female allotype of Chaerilus chapmani (MNHN) and male holotype and female paratype of Chaerilus agnellivanniorum sp. n. (MZUF–MNHN).
| Chaerilus chapmani | Chaerilus agnellivanniorum | |||
| Total length (including telson) | 30.9 | 39.2 | 36.8 | 40.9 |
| Carapace | ||||
| Length | 4.5 | 6.5 | 5.6 | 6.6 |
| Anterior width | 2.4 | 3.3 | 3.4 | 3.9 |
| Posterior width | 4.8 | 6.3 | 5.6 | 6.7 |
| Mesosoma length | 7.5 | 9.6 | 10.3 | 10.8 |
| Metasomal segment I | ||||
| Length | 2.0 | 2.1 | 2.1 | 2.3 |
| Width | 1.8 | 2.4 | 2.2 | 2.5 |
| Metasomal segment II | ||||
| Length | 2.3 | 2.6 | 2.4 | 2.6 |
| Width | 1.6 | 2.2 | 1.9 | 2.2 |
| Metasomal segment III | ||||
| Length | 2.4 | 2.9 | 2.5 | 2.7 |
| Width | 1.5 | 2.1 | 1.8 | 2.2 |
| Metasomal segment IV | ||||
| Length | 2.8 | 3.3 | 2.9 | 3.3 |
| Width | 1.4 | 2.0 | 1.8 | 1.9 |
| Metasoma, segment V | ||||
| Length | 4.1 | 5.5 | 5.2 | 5.8 |
| Width | 1.4 | 1.8 | 1.7 | 1.8 |
| Depth | 1.1 | 1.5 | 1.3 | 1.5 |
| Telson length | 5.3 | 6.7 | 5.8 | 6.8 |
| Vesicle | ||||
| Width | 1.6 | 2.0 | 2.1 | 2.1 |
| Depth | 1.5 | 1.8 | 1.8 | 1.8 |
| Femur | ||||
| Length | 7.4 | 8.1 | 8.5 | 8.1 |
| Width | 1.4 | 2.1 | 1.8 | 2.4 |
| Patella | ||||
| Length | 7.5 | 8.0 | 8.6 | 8.4 |
| Width | 1.4 | 1.9 | 1.8 | 2.4 |
| Chela | ||||
| Length | 13.5 | 16.0 | 15.6 | 15.5 |
| Width | 2.1 | 3.2 | 2.6 | 3.8 |
| Depth | 2.1 | 3.2 | 2.6 | 3.6 |
| Movable finger | ||||
| Length | 6.6 | 8.8 | 7.2 | 7.6 |
Coloration: Basically yellow to reddish-yellow. Carapace yellow on males and reddish-yellow on females. Tergites with the same colour as the carapaces and some diffused confluent yellow pigmentation. Metasomal segments yellow, paler on males; carinae darker on females. Telson yellow with the aculeus reddish-yellow to reddish. Chelicerae yellow without any variegated spots; fingers darker with reddish teeth. Pedipalps reddish-yellow to dark reddish; chela fingers darker, almost blackish. Legs yellow to slightly reddish-yellow. Venter and sternites yellow with some infuscations; pectines pale yellow.
Morphology: Carapace moderately narrowed anteriorly in both sexes; anterior margin weakly emarginated; carinae and granulations moderate to strong, better marked on females; furrows moderately deep in both sexes. Two pairs of lateral eyes, and a pair of median eyes; all eyes of normal size without any reduction or regression; median eyes about twice the size of the lateral eyes; median eyes anterior to the centre of the carapace. Tergites with moderate to strong granulations better marked in the female; carinae weak. Sternum pentagonal, longer than wide; genital plates of the operculum oval to sub-oval in shape. Pectinal teeth count 4–4 in the male holotype, 3–3 in the female paratype (see diagnosis for variation). Sternites almost smooth, with some granulations in the female; spiracles small and round-shaped; carinae absent from VII. Metasomal segment I wider than long; segments II to V longer than wide. All the carinae strongly granular; ventral carinae reduced on I, moderately marked on II; segments IV and V with carinae composed of stronger moderately spinoid granules. Vesicle very elongated with a pear-like shape, smooth, with a short aculeus. Pedipalps: femur with five carinae; internal having spinoid granules. Patella with seven carinae; dorsal and dorso-external moderately granular; dorso and basal-internal with strong spinoid granules. Chela with eight carinae, moderately to strongly granular; internal carinae with strong spinoid granules. Tegument with few granulations. Finger shorter than manus, with 6–7 rows of granulations on the dentate margins. Chelicerae characteristic of the family Chaerilidae [24]. Trichobothriotaxy type B; orthobothriotaxic [22]; femur with 9 trichobothria, patella with 14, and chela with 14. Legs with pedal spurs moderately developed. Tarsi with two rows of spiniform setae.
4 Ecological remarks about Chaerilidae species living in caves
A few species of Chaerilidae are known from caves [25] but only two species have their status of true troglobitic confirmed: Chaerilourencous sabinae and Chaerilus chapmani [17,26]. Other species are suspected of having a subterranean way of life, mainly because found inside caves, but more formal evidence may yet be necessary to demonstrate this aspect. These are the cases of Chaerilus cavernicola Pocock, 1893 found in the Ngalau cave in Sumatra and Chaerilus agilis Pocock, 1899 collected in the Selangor cave from Malaya. These species remain, however, extremely rare; only recently, the male of C. agilis was finally described [20,27].
The condition of Chaerilus chapmani as a true troglobitic element is unequivocal. This species was found exclusively in the deepest sections of the caves in Gunong Mulu National Park and no individual of this species was found outside the caves. Besides both adult and juveniles specimens have been found inside the caves, suggesting therefore that the entire cycle takes place in parallel with a subterranean way of life [17].
Concerning the new species described at present, both adult and juvenile forms were equally being found in very deep sections of the cave. This could suggest a permanent subterranean way of life, although many morphological traits do not show major regressions.
All specimens of Chaerilus agnellivanniorum sp. n. were collected deeply in the caves of ‘Natuturingam’ (the local Filipino name), better known as Puerto Princesa Underground River Cave (Fig. 1), more than 1 km from entry and more or less near the subterranean river (Fig. 8). In the map, the two entrances are indicated as ‘daylight’ and ‘outflow’. The scorpions were found at the ground level, on rocks and on guano. They were present in both areas with much guano and in areas with small amounts of guano. The scorpions were never under rocks, but always on them. Besides, they were not present in vertical rock walls. Also a great amount of other arthropods were found and collected in the caves, such as Arachnida, Myriapoda, Insects, Crustacea.

Interior view of Puerto Princesa Underground River Cave, showing the typical habitat.
It is interesting to underline that the specimens of Chaerilus agnellivanniorum sp. n. did not fluoresce under UV lamp while researchers were collecting them in the caves (Fig. 9). The experiment was repeated also with the preserved specimens. This confirms the most interesting discovery by Lourenço [28], which is supported even by an additional test on a recently described species, Chaerilus merendinoi Rossi, 2018, from Bali, Indonesia [18].

Chaerilus agnellivanniorum sp. n., male paratype, alive in natural habitat.
5 Biogeographic considerations
Palawan Island is stretched between Mindoro Island in the northeast and Borneo Island in the southwest. Palawan is about 450 km long and 50 km wide. With an area of 12,188.6 km2, it is the fifth largest island of the country and the 64th in the world. For its peculiar position, between Filipino islands and Indonesian islands, it was, and still is, the subject of important studies in biogeography and zoogeography. Since it is in a particular geographic position, it may have served as a springboard to the oceanic Philippines for some organisms, while simultaneously representing a dead-end Sundaic peninsula for others [29]. According to the most recent discoveries, using multiple approaches, it seems evident that Palawan and Borneo were not connected by a land bridge at 17,000 year BP. Voris and Reis & Garong [30,31] noted that if Palawan was really connected to the Greater Sundas and mainland Southeast Asia via land-bridges during the last two major glacial periods (ca. 18,000 and 160,000 year BP), then it should be assessed through the fossil record because species indicative of main-land fauna, such as large carnivores, may have occurred on Palawan during the Middle and Late Pleistocene. In reality, there are neither fossil nor modern tiger (Panthera tigris), clouded leopard (Neofelis nebulosa), or sun bear (Helarctos malayanus) documented from Palawan, but there are from Borneo, Java, Sumatra, and Bali, thus it was suggested [31] that these large carnivores may have not dispersed from the Sunda Shelf in the Pleistocene. Already Heaney [32] showed that Palawan and Borneo are now separated by a water depth of about 145 m and taking into account the fact that the sea level was reduced in the past by only 120 m, a land bridge really did not exist between these two islands. In conclusion, as already suggested by Lourenço [16] for the new species here described, and also as outlined by Rossi [4,18] for other Chaerilids species, to deal with the taxonomy of scorpions from this area, which has a very complex geological past, it is fundamental to take in consideration all aspects of biogeography and not only those based on morphology (typology). The idea that a single species of small land arthropods such as Chaerilus celebensis can be present in largely separated islands and territories such as Luzon, Sulawesi and continental Thailand, never connected by landbridges, can hardly be accepted [4]. Moreover, the presence of the same Chaerilus species in two different cave systems from two islands not connected in the late Pleistocene can only be rejected [16].
Acknowledgements
We are most grateful to Dr Paolo Agnelli (MZUF) and Dr Stefano Vanni (MZUF) for information on the 2017 field expedition to Palawan and documentation on the caves, to Dr Alessio Romeo for photos of habitat and alive specimens, to Dr Marco Borri (MZUF) for information on the scorpions collected in the 1991 expedition, to Prof. Jo De Waele (University of Bologna, Italy) for help in collecting scorpions. Field investigations were organized by “La Venta Esplorazioni Geografiche” of Treviso, Italy, in collaboration with the “TagBalay Foundation” of Puerto Princesa (Palawan) and the “Puerto Princesa Underground River–Protected Areas Management Bureau (PPUR-PAMB)”, as part of a project promoted by the Philippines Government named “Support for Sustainable Eco-Tourism in the Puerto Princesa Underground River Area”. Our thanks go also to Miss Élise-Anne Leguin, (MNHN), Paris, for assistance in the preparation of photos and plates.