

Abridged English version
Dwarf pine (Pinus mugo Turra) constitutes a bush colonizing rocky slopes in subalpine ecosystems. In Europe, the present-day repartition of Pinus mugo is highly fragmented, from the Balkans to the occidental Alps. In the Val di Susa (Alps Piedmont, Italy), several small populations are identified, suggesting that this zone was potentially a refuge area for this species during the last glaciation. However, any palaeoecological data supported this assumption. This lack of data is related to the fact that, at this state of research, the Pinus section sylvestris (Pinus sylvestris L, Pinus uncinata Mill. and Pinus mugo Turra) are not discernible either by their needle, pollen grain or wood anatomy. This note presents original data based on fossil cones of Pinus mugo Turra preserved in travertine sequences located in the Val di Susa at 1340 (San Domenico) and 1890 m a.s.l (Selle 1 and 2). Travertine deposits are calcareous formations resulting from saturation of water in bicarbonate and biological processes. They often enclose plant imprints and charcoal fragments. Plant imprints were extracted from travertine blocks and drawn using a stereomicroscope (×6; ×12) coupled with a ‘camera lucida’. Taxonomic identifications were achieved on the observation of morphological features of fossils compared in specimens from reference collection and data from morphological atlases. Charcoal identifications were achieved via comparisons of subfossil materials with a reference collection of charred modern wood species and wood anatomy atlases.
Three U/Th (Uranium/Thorium) dates on calcite were obtained, revealing that the travertine system of San Domenico was built during the Late Dryas (11 506 ± 66 U/Th BP) and the Selle system at the beginning of the Holocene (10 145 ± 225 and 9475 ± 670 U/Th BP). In the travertine of San Domenico, several cone imprints of Pinus sylvestris , Pinus uncinata Pinus mugo and some charcoal fragments of Pinus type sylvestris were identified. In this travertine, leaf imprints of Hippophae rhamnoides and Salix sp. were also collected. These results indicate that, around the travertine system of San Domenico, during the Late Dryas, woodlands of Pinus sylvestris probably dominated the vegetation, with some individuals of Pinus uncinata and Pinus mugo. In the travertine of Selle, the needles of Larix decidua are abundant, with leaf imprints of Salix cf. cinerea , Salix cf. caprea , Acer opalus , Betula cf. pubescens , Sorbus aucuparia , Laburnum sp. and Corylus avellana . In this travertine, three cone imprints of Pinus mugo and charcoal fragments of Larix-Picea (probably Larix, ) were identified. These results indicate that Larix decidua was probably the dominant tree around the travertine at the beginning of the Holocene, with some individuals of Pinus mugo.
The plant assemblages identified in the travertine systems of San Domenico and Selle display open woodlands with Pinus mugo, mostly composed of heliophilous species such as Larix decidua, Hippophae rhamnoides and Betula pubescens. Fire perturbations, testified by the occurrence of charcoal fragments in the two systems seemed to be a major factor structuring the local vegetation during the Lateglacial–Holocene transition.
The different results presented in this note reveal that Pinus mugo was present in the occidental Alps at least since the Late Dryas. These data support the hypothesis that the Val di Susa was potentially a refuge area of this species during the last glaciation. However, our data display that this pine was not abundant in the vegetation during the Late Dryas. Data from central Europe pinpoint that Pinus mugo was a frequent taxon with Pinus sylvestris during the Lateglacial. At the beginning of the Holocene, this pine was replaced by woodlands dominated by Pinus cembra, Picea abies, and Larix decidua. This vegetation dynamics is interpreted as the direct consequence of the warming of the climate, with the establishment of favourable environmental conditions for the development of more competitive coniferous species. In areas where Pinus cembra and the other conifers were present during and after the glaciation, Pinus mugo was driven out, maintaining mostly on inhospitable rocky zones. This ecological dynamics could be retained for explaining the present-day spatial distribution of Pinus mugo in the Val di Susa. Pinus mugo is thus a pioneer species, but not competitive in comparison to the other coniferous species dominating the present-day mountain ecosystems in the Alps. In the past, this pine had probably a more high spatial distribution than today in the occidental Alps.
1 Introduction
Dans le Val de Suse (Alpes internes italiennes, Piémont), de petites populations de pin mugo (Pinus mugo Turra) suggèrent que cette zone a pu se trouver à proximité d'une aire refuge de cette espèce durant les périodes de glaciations, avec une répartition périglaciaire probablement plus importante au niveau ce massif [1,2]. Cependant, aucune donnée scientifique n'est venue étayer cette hypothèse de zone refuge, qui est fragile au regard des opérations de reboisement datant de l'époque mussolinienne. Le pin mugo est un arbuste possédant des axes prostrés, ayant une grande capacité de marcottage et qui colonise les pentes rocheuses des écosystèmes subalpins [3]. Ce pin présente une très grande résistance à la sécheresse, un besoin crucial de lumière, de très faibles exigences tropiques et craint la concurrence des autres espèces subalpines [4]. Il s'accommode de tous les types de substrat, avec cependant une préférence pour les sols calcaires, et est parfaitement adapté à l'ensevelissement par la neige. Les populations actuelles piémontaises sont fragmentées par rapport à l'aire principale de l'espèce, située en Europe centrale et orientale [5]. Les petites populations du Val de Suse, composées de quelques individus, soulèvent des interrogations d'ordre génétique, mais également biogéographique. En effet, peu de données font état de l'impact de la fragmentation de l'aire de distribution du pin mugo sur la structuration génétique et phénotypique de l'espèce [6]. Par ailleurs, la déprise pastorale qui sévit depuis plus d'un siècle dans les Alpes semble stimuler la dynamique du pin mugo, compte tenu de sa valence écologique et de sa capacité à coloniser rapidement les espaces ouverts abandonnés [5–7].
La paléoécologie peut permettre de répondre aux interrogations au sujet du caractère naturel ou introduit d'une espèce dans une région [8,9]. Le manque de données paléoécologiques sur Pinus mugo est en grande partie dû au fait que les pins de la section Pinus sylvestris, dont fait partie le pin mugo, ne sont discriminables, ni par leur pollen [10], ni par l'anatomie de leur bois [11]. La présente note fait état de la découverte d'empreintes de cônes de Pinus mugo Turra dans deux systèmes travertineux fossiles localisés dans le Val de Suse (San Domenico et Selle). Les travertins sont des dépôts calcaires issus de la saturation des eaux de rivières en bicarbonate soluble [12]. Archives paléoenvironnementales, ces formations naturelles renferment fréquemment des empreintes végétales et des charbons de bois identifiables [13,14]. La mesure du temps sur ces dépôts est réalisée grâce à des datations au 14C pratiquées sur les charbons de bois ou s'appuyant sur le rapport uranium/thorium (U/Th) mesuré sur les faciès travertineux. Les résultats présentés dans cet article permettent d'affirmer que le pin mugo (Pinus mugo Turra) faisait partie de la flore des Alpes piémontaises au moins depuis le Tardiglaciaire, et de discuter son statut écologique durant cette période.
2 Matériels et méthodes
2.1 Présentation des sites
Les systèmes travertineux de San Domenico (45°23N, 7°11E) et de Selle (45°4N, 6°55E) sont localisés dans le Val de Suse (Alpes piémontaises), sur un substrat essentiellement constitué de calschistes et de dolomies. D'après les données météorologiques de la station de Salbertrand-le-Selle (altitude : 1950 m), la température moyenne annuelle est de 4,5 °C, celle du mois le plus froid (décembre) est de et celle du mois le plus chaud (août) 12,6 °C. La précipitation moyenne annuelle est de 869 mm, avec un maximum au cours du mois de mai (86 mm). Le travertin de San Domenico se situe à 1340 m sur le versant ouest du Monte Genevris, qui sépare la vallée de Chiomonte de celle de Suse. La végétation autour du site de San Domenico est dominée par le pin sylvestre (Pinus sylvestris L.) et le mélèze (Larix decidua Mill.), associé à des espèces héliophiles telles que Amelanchier ovalis Medik., Prunus spinosa L. et Berberis vulgaris. Le site de Selle se situe dans le parc naturel Gran Bosco di Salbertrand, sur le même versant et en amont du travertin de San Domenico. Deux dépôts sont localisés à 1890 m d'altitude (Selle 1 et 2). La végétation autour du système travertineux se caractérise par des boisements jeunes dominés par Larix decidua accompagnés de Pinus cembra L., Picea abies (L.) Karst., Abies alba Mill., Populus tremula L. Le sous-bois ligneux est composé majoritairement d'Ericaceae, tels que Rhododendron ferrugineum L., Vaccinium myrtillus L., V. vitis-idaea L., V. uliginosum L. et de Juniperus communis L.
2.2 Échantillonnage et identification des macrorestes
Les blocs de travertins extraits des séquences contiennent des empreintes qui sont dégagées et dessinées à l'aide d'une loupe binoculaire (×3, ×6) équipée d'une chambre claire. La détermination taxinomique des échantillons fossiles s'effectue par comparaison des structures morphologiques conservées avec des spécimens actuels de collections de références (Centre de bio-archéologie et d'écologie, herbier de l'institut de botanique de Montpellier).
Les critères d'identification utilisés dans cette étude paléobotanique pour discriminer les pins montagnards et subalpins de la section sylvestris (Pinus sylvestris, Pinus uncinata et Pinus mugo) s'appuient sur la morphologie des écussons des écailles et la position du mucron sur les écussons. Ces critères sont utilisés dans certaines flores pour distinguer les différentes espèces de pins de l'Europe occidentale [3–15]. Les observations qui nous ont permis d'établir les éléments de diagnose s'appuient sur des cônes actuels provenant des Tatras (Slovaquie), de la haute vallée de la Maurienne (Savoie, France), de l'Aigoual (Gard, France) et du Queyras (Hautes-Alpes, France).
Les charbons de bois contenus dans les travertins ont été identifiés au microscope épiscopique (métallographique) à grossissement () par comparaison avec des charbons de collection de référence, mais aussi par la consultation d'ouvrages xylologiques [16,17] et d'articles spécialisés d'anatomie de charbon de bois [17,18].
2.3 Datation 238U/230Th
Les systèmes travertineux ont été datés par U/Th selon une nouvelle méthode de comptage des ions radioactifs (MIC–ICP–MS : Multi-collector, Inductively Coupled with Plasma Mass Spectrometry) développée par Fietzke et al. [19]. Pour qu'une date U/Th soit acceptable, il faut s'assurer que l'échantillon n'ait pas été contaminé par du thorium détritique [20]. Le rapport 230Th/232Th permet d'évaluer le taux de contamination. Expérimentalement ce rapport doit être selon les auteurs, supérieur à 17 [21] ou à 20 [20]. Les datations U/Th permettent d'obtenir des âges calendaires comparables aux datations 14C calibrées.
3 Résultats
3.1 Datations et chronologie
Les dates obtenues au Selle présentent des rapports 230Th/232Th relativement faibles (Tableau 1). Il faut noter que ces dates ont été corrigées en prenant en compte le taux de contamination effectif. Par ailleurs, dans le cas de rapport 230Th/232Th faibles, les dates ne sont pas automatiquement rejetées. En effet, des chronoséquences établies à partir de dates U/Th a priori erronées se révèlent parfaitement correctes après confrontation avec des datations 14C [22,23]. De ce fait, les dates U/Th, mêmes associées à de faibles valeurs du rapport 230Th/232Th, doivent être considérées favorablement, sauf dans le cas de totale incohérence chronostratigraphique (Yves Quinif, communication orale). Les dates montrent que le système travertineux de San Domenico s'est construit durant le Dryas récent, aux alentours de 11 506 ± 66 U/Th BP. Le système travertineux du Selle s'est probablement mis en place au début de l'Holocène, entre 10 145 ± 225 et 9475 ± 670 U/Th BP.
Dates U/Th
| Site | Date BP | 230Th/232Th | Laboratoire |
| San Domenico | 11 506 ± 66 | 67,99 | IFM-GEOMAR, Allemagne |
| Selle 1 | 10145 ± 225 | 11,25 | IFM-GEOMAR, Allemagne |
| Selle 2 | 9475 ± 670 | 3,83 | IFM-GEOMAR, Allemagne |
3.2 La paléoflore de San Domenico (1340 m)
La base du système travertineux a livré de nombreuses empreintes de feuilles d'Hippophae rhamnoides , de Salix sp. , des aiguilles de pins de la section sylvestris, des aiguilles de Larix decidua, des cônes de Pinus uncinata (, Fig. 1), de Pinus sylvestris (, Fig. 2A) et de Pinus mugo (, Fig. 2B). Quelques charbons de bois de Pinus type sylvestris ont été également identifiés. Parmi ces espèces héliophiles strictes au stade plantule, Pinus uncinata est l'espèce la plus tolérante à l'ombre. Ces données font référence à une végétation probablement ouverte et soumise à des incendies.
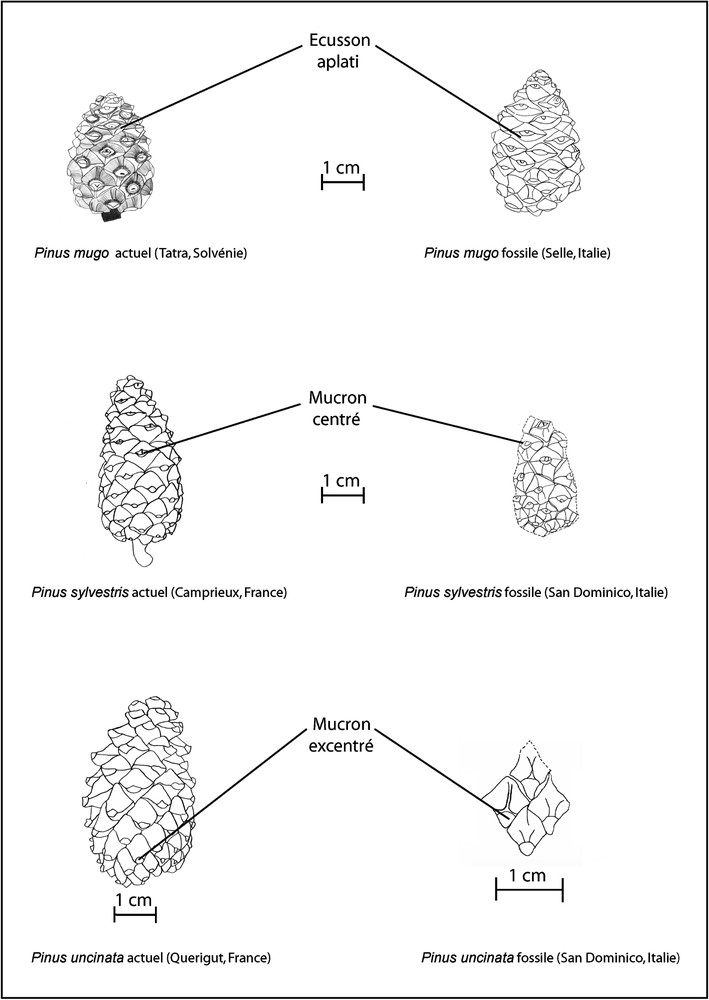
Dessins de cônes actuels et fossiles de Pinus mugo Turra, Pinus sylvestris L. et Pinus uncinata Mill.
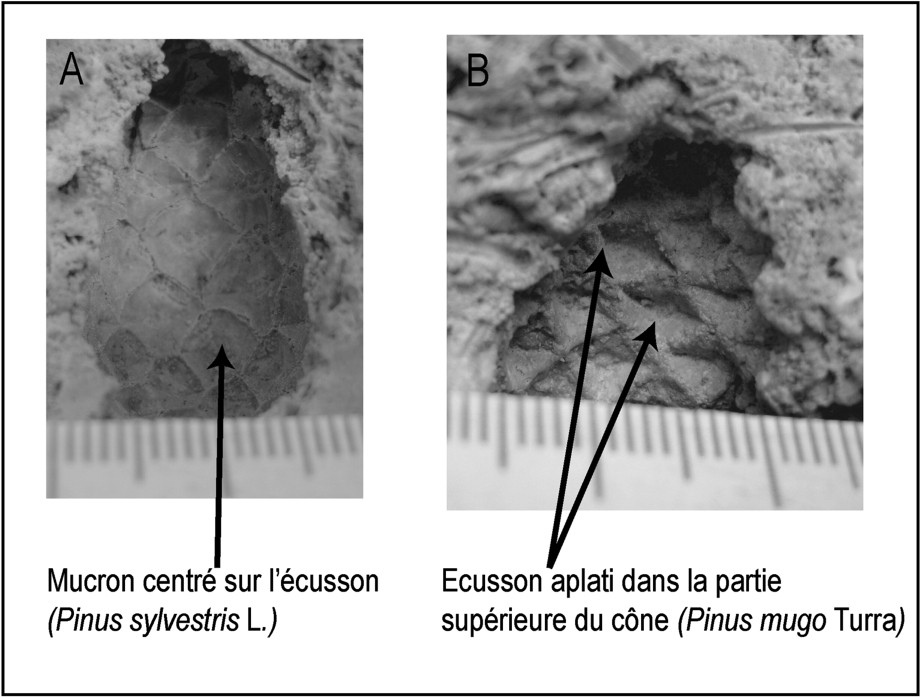
Empreintes fossiles de cônes identifiées dans le système travertineux de San Domenico (Val de Suse, Italie). (A) Pinus sylvestris L. (B) Pinus mugo Turra.
3.3 La paléoflore du Selle (1890 m)
Les niveaux datés sont riches en empreintes d'aiguilles de Larix decidua, associées à des aiguilles de Pinus cembra et de Pinus type sylvestris (Tableau 2). Trois cônes de Pinus mugo ont été identifiés (Fig. 1). Des empreintes de Salix cf. cinerea , Salix cf. caprea , Acer opalus , Betula cf. pubescens , Sorbus aucuparia , Laburnum sp. , Corylus avellana ont été également recensées. Les identifications de charbons de bois de trois niveaux associés à la coupe de Selle 2 montre la prédominance des restes de type Larix–Picea . La transition abrupte entre le bois initial et le bois final laisse penser que les fragments appartiendraient au mélèze, cf. Larix [17,18]. L'ensemble des résultats montre qu'au début de l'Holocène, la végétation du Selle était dominée par Larix decidua, avec du Pinus cembra et du Pinus mugo. Dans cette végétation, des feuillus strictement héliophiles au stade plantule (Salix sp.) et d'autres plus tolérants à l'ombre tels que Acer opalus, Laburnum sp. et Sorbus aucuparia étaient également présents. Les charbons de bois de cf. Larix attestent l'existence d'incendies dans la zone d'étude.
Liste des taxons et des types de reste identifiés dans les systèmes travertineux de San Domenico et du Selle (Val de Suse, Italie)
| Taxons | San Domenico (1340 m, 11 506 ± 66 BP) | Selle 1 (1890 m, 10 145 ± 225 BP) | Selle 2 (1890 m, 9475 ± 670 BP) |
| Larix decidua | aiguille | aiguille | aiguille, charbon |
| Pinus mugo | cône | cône | cône |
| Pinus sylvestris | cône | ||
| Pinus uncinata | cône | ||
| Pinus type sylvestris | aguille, charbon | aiguille | aiguille |
| Pinus cembra | aiguille | aiguille, charbon | |
| Acer opalus | feuille | ||
| Hippophae rhamnoides | feuille | ||
| Salix sp. | feuille | ||
| Salix cf. caprea | feuille | feuille | |
| Salix cf. cinerea | feuille | feuille | |
| Populus tremula | feuille | ||
| Corylus avellana | feuille | ||
| Betula cf. pubescens | feuille | ||
| Sorbus aucuparia | feuille | ||
| Rubus sp. | feuille | ||
| Laburnum sp. | feuille |
4 Discussion
4.1 Communautés à Pinus mugo Turra
Le pin mugo est une espèce des écosystèmes d'altitudes peu compétitrice. Les assemblages enregistrés au Selle et à San Domenico traduisent des communautés ouvertes sous l'influence de fortes contraintes environnementales (par exemple, limite supérieure des arbres) ou de perturbations fréquentes (par exemple, incendies, avalanches).
Les assemblages matérialisés par Pinus mugo se caractérisent par une composition à tendance héliophiles (i.e. Larix decidua, Pinus sylvestris, Salix sp., Hippophae rhamnoides, Betula pubescens, etc.) bien que des espèces de fin de succession aient été également observées (Pinus cembra, Pinus uncinata). Il est possible que l'abondance des espèces hélophiles dans les assemblages puisse être mis en relation avec des ouvertures du milieu, provoquées par des incendies, attestés par la présence de charbons de bois dans les deux systèmes travertineux.
Dans les zones d'avalanche, les espèces observées n'abondent pas ; bien au contraire, elles ne résistent pas au caractère répétitif des coulées de neige. Dans les couloirs d'avalanches, l'espèce la plus abondante reste l'aulne vert – Alnus viridis (Chaix) DC –, qui n'a été observé, ni à San Domenico, ni au Selle. Il est peu probable que l'hypothèse de l'avalanche puisse être retenue pour expliquer l'abondance des espèces héliophiles. De même, l'hypothèse de la limite supérieure des arbres semble peu cohérente au regard de la composition floristique des assemblages, contenant Acer opalus et Corylus avellana, qui sont des espèces qui se développent préférentiellement dans l'étage montagnard. Le feu est donc probablement l'un des facteurs ayant contribué au développement des espèces héliophiles dans la zone d'étude.
4.2 Élément de biogéographie historique de Pinus mugo Turra
Les assemblages de macroflores de San Domenico et de Selle mettent en évidence, pour la première fois, la présence du pin mugo (Pinus mugo Turra) dans la végétation des Alpes occidentales durant la transition Tardiglaciaire–Holocène. Ce résultat renforce l'hypothèse selon laquelle cette zone se situe à proximité d'une aire refuge de ce pin durant la dernière glaciation (Würm), et que les petites populations recensées dans le Val de Suse peuvent être naturelles.
Dans certaines zones de l'Europe centrale, comme la Roumanie et la Pologne, où l'espèce présente actuellement une répartition plus homogène et moins fragmentée, des études fondées sur l'analyse de restes macrofossiles préservés dans des lacs ou des tourbières montrent que, durant le Tardiglaciaire, le pin mugo associé à Pinus sylvestris était une espèce abondante dans la végétation [24]. Par la suite les boisements à Pinus mugo et Pinus sylvestris ont été remplacés par des formations dominées par Pinus cembra, Larix decidua et Picea abies [24]. Cette dynamique de la végétation est interprétée comme une réponse écologique faisant suite à l'augmentation graduelle de la température et à la mise en place de conditions favorables au développement d'autres espèces de conifères plus compétitrices [24,25].
La biogéographie postglaciaire du pin mugo, du moins dans la partie occidentale de son aire de distribution, a certainement été en partie façonnée par la compétition interspécifique. Dans les zones où des boisements à Pinus cembra, Pinus uncinata, Larix decidua et Picea abies étaient présents durant et après la glaciation, le pin mugo ne s'est maintenu que dans les zones rocailleuses moins favorables à l'établissement de forêts denses. Dans le Val de Suse, ce scénario écologique peut être retenu pour expliquer la distribution actuelle du pin mugo, hypothèse également évoquée en ce qui concerne les populations de pin mugo des Alpes maritimes et ligures [26]. L'importance actuelle de cette espèce dans les Alpes orientales, les Carpates et les Balkans serait associée à un ou plusieurs refuges de cette espèce localisés dans ces régions [26,27], mais également au fait que l'Europe de l'Est était peu soumise aux glaciations du Pléistocène, agent de fragmentation de l'aire de Pinus mugo dans les Alpes occidentales. Il est important de souligner que des études biométriques, visant à quantifier l'impact du processus de fragmentation spatiale sur la variabilité morphologique et génétique, soulignent peu de différences entre des populations de pin mugo, caractérisées par un grand nombre d'individus, de l'Est et de l'Ouest de l'Europe [26,28]. Ces études suggèrent une origine commune et une fragmentation Pléistocène récente avec un flux génétique important entre les populations.
5 Conclusion
La présence d'empreintes de cône de Pinus mugo dans des gisements travertineux tardiglaciaires et holocènes du Val de Suse (Alpes piémontaises, Italie) montrent que cette espèce était présente dans ou à proximité des Alpes occidentales durant la dernière glaciation. Les petites populations fragmentées actuelles résultent probablement d'une aire postglaciaire plus étendue. Dans les Alpes occidentales, la dynamique postglaciaire de ce pin prostré fut probablement inhibée par les autres espèces de conifères plus compétitifs qui dominent actuellement la végétation de l'étage subalpin. Les reconstitutions paléoécologiques, fondées essentiellement sur les analyses palynologiques, ont négligé le rôle de certains pins de montagne (Pinus uncinata et Pinus mugo) dans les dynamiques postglaciaires de la végétation [29]. Les communautés héliophiles à Pinus mugo semblent associées aux incendies. Les changements climatiques (augmentation des températures et diminution des précipitations) et la déprise agricole en cours dans les montagnes du Sud de l'Europe pourraient permettre au pin mugo de coloniser les espaces asylvatiques abandonnés par les pasteurs, mais aussi les zones se trouvant au voisinage de la limite supérieure des arbres [7]. Il pourrait donc résulter une expansion de Pinus mugo dans les massifs où il est actuellement présent sporadiquement. Le rôle des feux dans la conservation des habitats à pin mugo reste à élucider.
Remerciements
Cette étude a bénéficié d'un financement du Centre national pour la recherche scientifique (CNRS). Adam A. Ali a bénéficié d'un contrat de post-doctorat (ATER) de l'École pratique des hautes études (EPHE). Cette étude participe au programme « Populations fractionnées et insulaires »de l'EPHE.