

Abridged version
1 Introduction
The geological site at Tercis comprises a continuous Campanian–Maastrichtian succession where several thousand samples including 2500 macrofossils have been collected and studied during years 1993–2000 under the aegis of the Subcommission on Cretaceous Stratigraphy of the Commission on Stratigraphy. Here is a summary of the results obtained by the 70 members of the relevant working group. These results have recently been gathered in a monograph [16].
2 Lithostratigraphy and physicochemical stratigraphy
The section in the quarry has been subdivided into two units. The d'Avezac Unit, from level 0 to level 100, is silex-free; the Les Vignes Unit, from level 100 upwards, bears silex (Fig. 1). Five subunits can be differentiated in the lower unit according to their alternatively high and low glaucony contents; their limits occur at levels 11.3, 30.0, 68.5 and 80.0; two subunits (limit at level 131.3 or so) can be distinguished in the Les Vignes Unit, with successively pale and dark flint from bottom to top.

Lithostratigraphie, physico-chimie et géochronologie de la série campano-maastrichtienne de Tercis. Les teneurs en glauconie supérieures à 1% sont notées par une flèche (maximum inférieur à 3%). Les cinq séquences proposées d'après la glauconie sont cohérentes avec les indices suggérés par les teneurs en manganèse : croissantes pour les cortèges transgressifs (CT) et décroissantes pour les cortèges de haut niveau (CHN) [17, p. 132]. La magnétostratigraphie, incomplète sur place, est suggérée par corrélation avec les dépôts des Apennins. Dans la colonne intervalles, les pointillés notent des condensations sédimentaires, de durées estimées d'après l'évolution de la glauconie. L'âge de la limite d'étage est estimé entre 71,0 et 72,5 Ma [16, p. 777].
Lithostratigraphy, physicochemistry and geochronology of the Campanian–Maastrichtian Tercis series. The five suggested sequences are consistent with the information suggested by Mn contents: increasing for transgressive (CT) and decreasing for highstand systems tracts (CHN) [17, p. 132]. Magnetostratigraphy is deduced by correlation with the Apennines [11]. In the column ‘intervalles’, dots indicate slightly condensed deposits. The stage boundary is located between 71.0 and 72.5 Ma [16, p. 777].
Sedimentological data suggest a continuous deposition with four decreases in rate around levels 6, 46, 67, and 83 as documented by larger amounts of glaucony and more common bivalves; traces of phosphate are also present around levels 6 and 67. In all cases, glaucony is poorly evolved and phosphatisation incomplete, which indicates that condensation must have been light [[14, p. 320], [17, p. 123]]. Five depositional sequences lacking low stand deposits have been suspected according to the variation in glaucony contents between levels 0 and about 130 [17].
Depositional mean rates have been estimated using acknowledged stage durations, acknowledged biozone durations, as well as rhythmic changes climatically constrained and interpreted as related to obliquity. These three different procedures led to consistent estimates of 1 m per 40 ka (25 m Ma−1), which, combined to the sedimentological data summarised above, allowed to establish a documented continuous time scale for the whole series [17].
Trace-element data documented the sequence interpretation; data on strontium-isotope ratios pointed to a diagenetic disturbance of the original composition [20]; palaeomagnetism has partly been documented in the Campanian portion of the Tercis section; in the rest of the section, it can be deduced using a precise biostratigraphic correlation between Tercis and Gubbio, one of the few land sections where palaeomagnetism is well recorded [1,11].
3 Biostratigraphy
Following long-term collection, several hundred ammonites, echinoids, and inoceramids have been obtained. Many other macrofossil groups are less abundant or less significant (sponges, corals, bryozoans, gastropods, other bivalves, nautiloids, crustaceans, asteroids, crinoids, fishes). The three main fossil groups document well-established biohorizons (Fig. 2), several of which can be recognised in the field. They allow correlation to other Tethyan or boreal European successions. Correlation to North America is possible using a number of common ammonite forms; inoceramids also appear very promising for the same purpose, according to studies in progress (I. Walaszczyk, in preparation). Two first occurrences and one last occurrence of ammonites, one first occurrence of an easy recognised inoceramid (genus Trochoceramus) have been selected for documentation of the Campanian–Maastrichtian stage boundary.
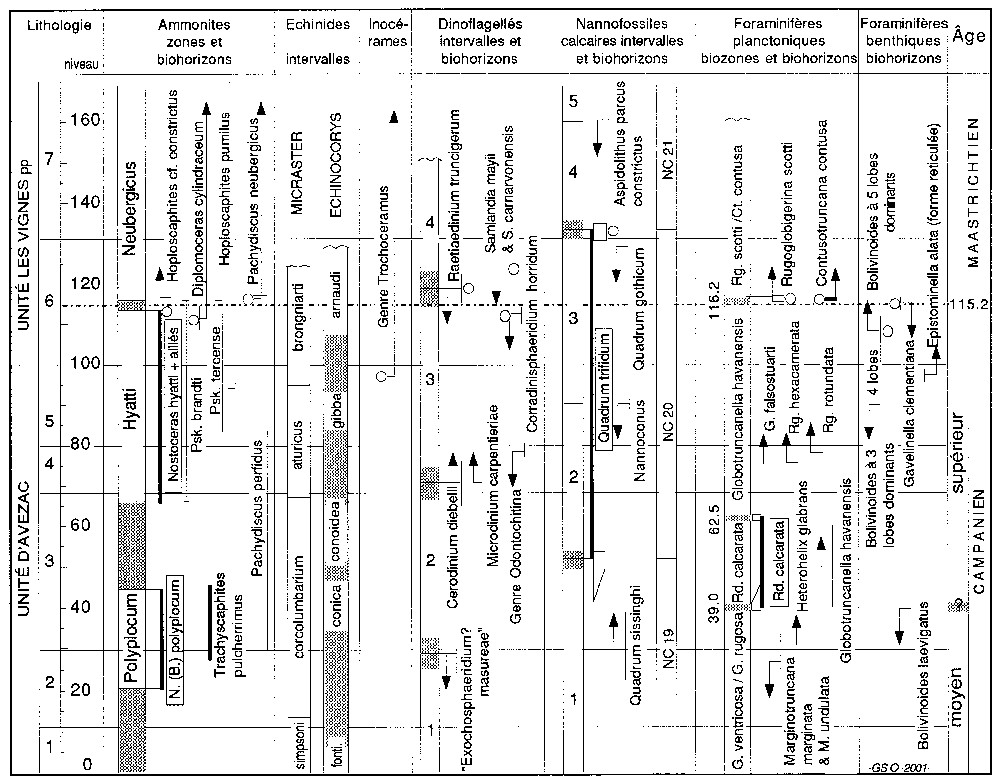
Biostratigraphie de la série de Tercis-les-Bains. Biozones NC 19 à NC 21 (Roth, 1978). Les intervalles locaux (dinoflagellés : 1 à 4, nannofossiles calcaires : 1 à 5) sont bornés par les biohorizons portés en regard avec leur incertitude. Abréviations : fonti, fonticola ; G., Globotruncana ; Psk., Pseudokossmaticeras ; Rd., Radotruncana. Cercles : 12 biohorizons retenus pour localiser la limite d'étage [16, p. 827].
Biostratigraphy of the Tercis-les-Bains series. Roth's 1978 biozones NC 19 to NC 21 are shown. Biohorizons shown with their uncertainty mark the limits between the local intervals (dinoflagellates: 1 to 4, calcareous nannofossils: 1 to 5). Abbreviations: fonti., fonticola; G., Globotruncana; Psk., Pseudokossmaticeras; Rd., Radotruncana. Circles: 12 biohorizons used for location of the stage boundary level [16, p. 827].
Microfossils, including pollen, dinoflagellate cysts, calcareous nannofossils, benthic foraminifera, planktonic foraminifera, ostracods, holothurians, ophiuroids are present at Tercis. Pollen will be used to correlate continental deposits with the marine chronostratigraphic calendar. Three fossil groups (dinoflagellate cysts, calcareous nannofossils, planktonic foraminifera) have been carefully and independently studied by three to four experts or by groups of experts. From comparison of the different results, an ‘experimental uncertainty’ could be defined and calculated; it will allow to establish correlations with a realistic practical uncertainty, a problem that used to be underestimated.
Three dinoflagellate cysts [2,3,21,22], one calcareous nannofossil [5,7,8,12,25], two benthic foraminiferas [24] and two planktonic foraminiferas [4,6,10,18] show significant biohorizons near the Campanian–Maastrichtian stage boundary (Fig. 2); they have been used to calculate the level where the stage boundary should be located.
4 Conclusions
Twelve biohorizons documented in the Tercis section (circles in Fig. 2) have been combined in order to establish a multi-criteria definition of the level where the Campanian–Maastrichtian boundary stratotype should be located. These biohorizons were selected in the vicinity of an a priori chosen ‘guide-event’; being the result of a combination, the newly proposed stage boundary (level 115.2 in the succession) is formally independent of any particular event.
In addition to this new defining procedure, the study at Tercis considered definition of uncertainties on the location of biostratigraphical changes.
It is believed that these improvements will allow more precise and more correct correlations of conventional stage boundaries. Its multiple basis will make the new definition recognisable in a wide palaeogeographic domain, with a precision that can be estimated smaller than 0.1 Ma, while previous uses quoted biohorizons that are known today to be included in a 2.5 Ma interval [16, p. 802].
The Campanian–Maastrichtian stage boundary, newly defined at Tercis, was agreed in 2001 and the Geological Site became the international reference and stratotype for the relevant interval of time.
1 Introduction
D'intéressantes formations campano-maastrichtiennes affleurent dans le site géologique de Tercis, une ancienne carrière située dans ce qui fut le Bassin aturien, au nord des Pyrénées [19]. Ce site, candidat à la définition de la limite Campanien–Maastrichtien [15], a été étudié, tandis que des démarches ont été entreprises afin d'assurer sa sauvegarde, avec la coopération des dirigeants de la société Ciments Calcia. Des milliers de prélèvements, dont plus de 2500 macrofossiles, ont été réalisés.
Cette présentation fait le point des résultats du Groupe de travail maastrichtien, qui a rassemblé des résultats sur ce site de 1992 à 2000. L'essentiel des expertises est rassemblé dans une monographie [16].
2 Lithostratigraphie et stratigraphie physicochimique
La lithologie [19] a conduit à séparer, entre les cotes 0 et 100 (les cotes sont approximativement métriques et inscrites sur le site), l'unité d'Avezac (calcaires sans silex, entre les cotes 100 et 170) et l'unité Les Vignes (calcaires à silex souvent en lits). Cinq sous-unités, distinguées par leur teneur en glauconie, ont été proposées pour l'unité inférieure (limites autour des cotes 11,3, 30,0, 68,5 et 80,0) et deux (limite à la cote 131,3), distinguées par la couleur dominante des silex, pour l'unité supérieure. Les épaisseurs reconnues dans notre description diffèrent de celles des pionniers de l'étude du site [9,23,26].
L'étude sédimentologique, et singulièrement celle de la glauconie, a permis d'interpréter la série comme résultant d'un dépôt de plate-forme externe continu, dont le taux s'est réduit à quatre reprises. Ces condensations sont faibles, d'après l'évolution modérée des grains verts et marquent des échanges eau de mer–sédiment pendant 0,1 à 0,4 Ma au plus [[14, p. 320], [17, p. 123]]. Vers les cotes 6 et 67, la présence d'éléments phosphatisés renforce l'impression de condensation, mais souligne sa modération par la rareté des nodules incomplètement phosphatisés. La glauconie suggère trois séquences (type Goldschmidt) ou cinq séquences génétiques, sans cortège de bas niveau marin [10], pour les niveaux 0 à 130 (Fig. 1).
Le taux moyen de dépôt a été calculé par diverses méthodes : (1) le quotient durée approchée des étages/épaisseur des dépôts correspondants, (2) le quotient durée estimée en divers lieux de biozones bien identifiées/épaisseur mesurée sur le lieu correspondant, (3) les rythmes de dépôt, de période métrique, dont l'origine climatique (obliquité de l'écliptique) est probable [28]. Ces approches signent un taux de 1 m pour 40 ka (25 m Ma−1) [10]. Ce résultat, combiné aux irrégularités notées plus haut, a permis d'établir l'échelle temporelle des phénomènes observés.
L'analyse des éléments en trace, des isotopes du strontium et du magnétisme a été abordée. Les éléments en trace ont aidé à caractériser des séquences. Les isotopes du strontium ont révélé une diagenèse des phosphates [20]. Le paléomagnétisme des couches campaniennes est cohérent avec les signaux fournis par les microfossiles entre les sites de Tercis et de Gubbio [1,11]. La comparaison niveau à niveau permet une localisation des intervalles magnétiques, avec une incertitude de l'ordre de 100 ka (Fig. 1).
3 Macrofossiles
Les macrofossiles ont fait la réputation du site de Tercis. Lors de notre étude, plus de 500 ammonites ont été récoltées [16, p. 551] ; c'est encore peu pour des déterminations spécifiques rendues délicates par une préservation souvent insuffisante. Trois taxons ont leur répartition totale dans le Campanien : Nostoceras hyatti Stephenson, 1941, Nostoceras (Bostrychoceras) polyplocum (Roemer, 1841) et Trachyscaphites pulcherrimus Roemer. Le Maastrichtien est caractérisé par la coexistence de Pachydiscus neubergicus (von Hauer, 1858) et Diplomoceras cylindraceum (Defrance, 1816). La répartition du premier est confortée par celles de son ascendant, Pachydiscus perfidus De Grossouvre, 1894 et de formes intermédiaires [16, p. 566]. Remarquable, enfin, est le petit Hoploscaphites pumilus Stephenson, 1941, forme américaine trouvée en nombre, pour la première fois en Europe, avec une durée de vie de près de 3 Ma sur le site. La disparition de N. hyatti, les apparitions de P. neubergicus et de D. cylindraceum ont été retenues pour caractériser la limite d'étage (Fig. 2).
Les échinides étaient les fossiles les plus prisés de Tercis. La détermination des espèces du genre Echinocorys ne fait pas l'objet d'un consensus, car leur variabilité morphologique intraspécifique est mal connue, tandis que des convergences de formes existent entre espèces différentes [9,13]. Au contraire, le genre Micraster livre des fossiles distincts, qui se relaient dans la partie campanienne de la succession (Fig. 2).
Les inocérames sont les fossiles les plus communs du site. Jusqu'en 2000, le genre Trochoceramus, identifiable sur le terrain, était considéré comme maastrichtien [27] ; à Tercis, il apparaı̂t environ 0,7 Ma avant sa limite inférieure. Ce signal a été retenu pour caractériser un moment antérieur à la limite. L'intérêt des inocérames sera renforcé par l'étude comparée des faunes des États-Unis et de Tercis (I. Walaszczyk, travaux en cours), qui laisse espérer la première corrélation fine Europe–Amérique pour le Crétacé final en faciès de plate-forme.
Brachiopodes, autres bivalves, astérides et dents de requins ont fourni des possibilités de corrélation meilleures que celles qu'offrent gastéropodes et crinoı̈des ; éponges (communes), coraux (rares, individuels) et bryozoaires restent à étudier, et reptiles à découvrir en place.
4 Microfossiles
Les groupes prometteurs ont été étudiés indépendamment par plusieurs experts ou groupes d'experts et les résultats ont été comparés.
Les grains de pollen et kystes de dinoflagellés sont excellemment préservés. Les premiers fournissent des repères stratigraphiques utilisables dans le domaine continental, souvent négligé dans les études de stratotypes. Les seconds ont livré des marqueurs de grand intérêt [3]. Quatre intervalles, longs de 1 à 2 Ma ont été proposés (1, 2, 3, 4, Fig. 2). L'incertitude sur leurs limites (apparition ou disparition d'un ou plusieurs taxons) et sur la position des biohorizons a été déduite de la « reproductibilité expérimentale », c'est-à-dire des différences de résultats entre études indépendantes réalisées par différents auteurs [2,21,22]. Cette reproductibilité a permis de sélectionner les meilleurs marqueurs. Pour repérer la limite Campanian–Maastrichtien, les disparitions des taxons Corradinisphaeridium horridum (Deflandre, 1937), Raetiaedinium truncigerum (Deflandre, 1937) et du couple Samlandia mayii (McMinn, 1988)/Samlandia carnarvonensis (McMinn, 1988) ont été retenues, suggérant des corrélations vers l'Europe boréale et vers la Téthys, jusqu'aux séries néo-zélandaises.
Les nannofossiles calcaires ont livré des informations dont la reproductibilité expérimentale est variable, parfois à mieux que 100 ka près [8]. Ils permettent une corrélation avec les dépôts téthysiens, mais aussi, pour les formes cosmopolites, avec les régions tempérées. Quadrum trifidum (Stradner) est un taxon reconnu dans les quatre études réalisées indépendamment [5,7,12,25] ; sa zone d'existence totale entre les cotes 52,5±2,4 et 134,2±2,7 entoure la limite d'étages, qui se situe à la base du tiers supérieur de cet intervalle, représentant 4 Ma de dépôt à Tercis. La disparition du taxon a été retenue pour caractériser un moment maastrichtien situé environ 0,8 Ma après la limite Campanien–Maastrichtien.
Les foraminifères planctoniques ont livré des informations plus significatives [4,6,10] que lors des études antérieures [23]. Le marqueur Radotruncana calcarata (Cushman, 1927) a été découvert puis illustré [18] entre les cotes 47 et 62 ; il est plus rare en dessous ; sa zone d'existence est réduite à 1 Ma environ. Sa disparition, longtemps utilisée dans la Téthys pour marquer la fin du Campanien, se trouve rejetée de 2,5 Ma dans le Campanien, tandis que son apparition pourrait servir à marquer la limite entre un Campanien moyen et un Campanien supérieur avec le concours d'autres marqueurs. L'apparition de Contusotruncana contusa (Cushman, 1926) et celle de Rugoglobigerina scotti Brönnimann ont été retenues [18] pour caractériser la limite d'étages.
Les foraminifères benthiques, intéressants pour des corrélations européennes, n'ont fait l'objet que d'études restreintes [24]. Deux marqueurs ont été retenus, l'un dans l'évolution du genre Bolivinoides (prédominance d'une forme à cinq lobes sur la dernière loge à un niveau situé sous la limite), l'autre à la disparition de Gavelinella clementiana (d'Orbigny), localisée à la limite d'étage.
Les ostracodes, les ossicules d'ophiures et d'holothuries ont fait l'objet d'examens qui mériteraient d'être complétés.
5 Conclusions
Grâce aux informations recueillies, le site géologique de Tercis a été accepté comme stratotype de la limite Campanien–Maastrichtien. Ces informations ont conduit à se départir de l'habitude de fixer une limite avec un seul groupe fossile et un seul événement, habitude tendant à mythifier la signification chronologique de cet événement et à limiter l'utilisation des unités définies au seul domaine où il est observé. L'utilisation d'une combinaison de biohorizons (12 cercles, Fig. 2) autour d'un événement guide ouvre de larges horizons pour une stratigraphie plus précise et plus exacte [15].
L'exigence de déterminer une incertitude et d'estimer les durées complète cette proposition de renouveau. La limite définie à Tercis à la cote 115,2 est exportable dans de vastes domaines, avec une incertitude de l'ordre de 100 ka, alors que les différentes limites utilisées auparavant s'échelonnaient sur 2,5 Ma [16, p. 802].
La carrière des Ciments Calcia est devenue le site géologique de Tercis. Son intérêt comme référence pour l'histoire géologique de la fin du Crétacé est reconnu. Il devrait être renforcé par des études précises sur inocérames, foraminifères benthiques, holothuries, microbrachiopodes et pollen, par une étude des géochronomètres présents, mais délicats et coûteux à mettre en œuvre, par un aménagement permettant de mener des investigations dans les couches déposées durant l'intervalle de temps 70–65 Ma, enfin, et très efficacement, par son aménagement comme lieu éducatif privilégié en géologie.
Remerciements
Les études ont été réalisées sur le terrain avec la permission de la société Calcia, dans le cadre d'un groupe de travail de la Sous-commission de stratigraphie du Crétacé. M.A. Lamaurelle a bien voulu apporter son aide constante à la réalisation de ces études et à la rédaction de leur compte rendu.