

Abridged version
1 Introduction
As far as we know, the influence of volcanic activity on the distribution of ostracod species has only been studied in the lacustrine environment [7]. Kagoshima Bay (Kagoshima Prefecture, Kyushu Island, southern Japan) is located in the southern part of the Japanese archipelago (Fig. 1). The presence of the active Sakurajima volcano authorises to study the influence of volcanism on the distribution of the benthic marine ostracod populations.
Localisation de la baie de Kagoshima (ı̂le de Kyushu, Japon), d'après Oki [20].
Index maps showing the location of the study area: Kagoshima Bay (Kyushu Island, Japan), after [20].
The physiography of the bay may be subdivided into three main parts [20,21]:
- – the mouth of the bay, with a maximum depth of about 100 m; on its west border, hydrothermal vents have been discovered;
- – the central part of the bay, with a maximum depth of 230 m, which is characterised by a topography-like basin;
- – the head of the bay, which is located in its northern part; it is separated from the central part of the Bay by a narrow channel between the western border of the Bay and the volcano.
The Kagoshima Bay is occupied by oceanic water: a branch of the Kuroshio Current enters along the eastern peninsula and flows southward along the western peninsula. The former is called ‘mass a’ [20], the latter ‘mass c’, which is equivalent to the low-salinity embayment water; it receives the discharge load of waste water as a result of the human activity around the bay. On their route, they meet some obstacles and are forced to change their direction. One of the consequences is the occurrence of strong currents and, in particular, in the central part of the bay, a counterclockwise current is the cause of the formation of two stagnant water masses b and d. The bay head area is occupied by an acidic water mass e (Fig. 2).
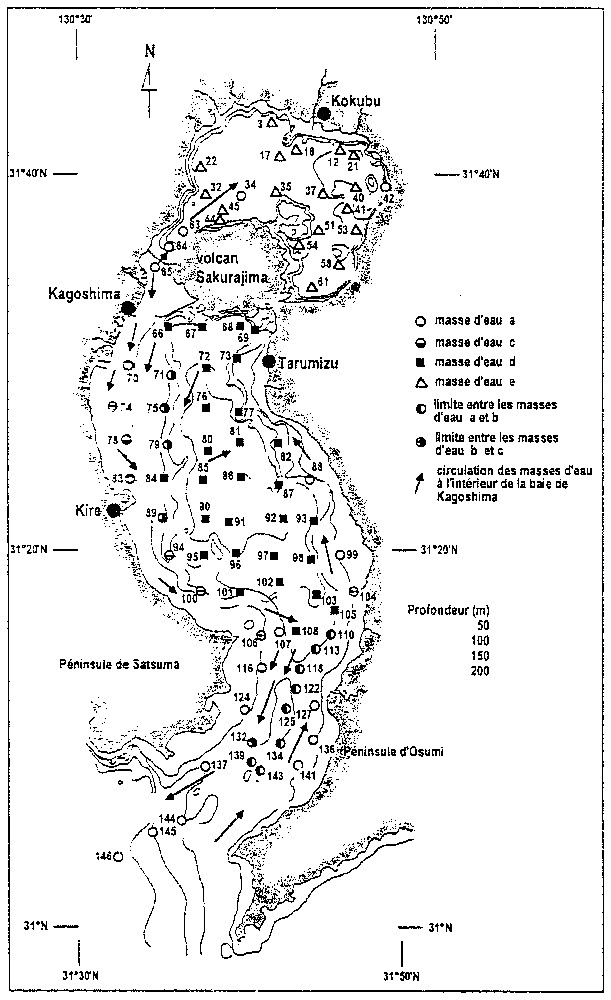
Distribution des stations de prélèvement et circulation des masses d'eau dans la baie de Kagoshima (ı̂le de Kyushu, Japon), d'après Oki [20].
Location of the sampling sites and circulation of the water masses in Kagoshima Bay (Kyushu island, Japan), after Oki [20].
2 Materials and methods
All samples were taken using PHLEGER's gravity corer. Hundred and forty-six samples of bottom sediments have been collected inside the bay by Oki from 1972 to 1978 [20]. The top centimetre of sediment core, approximately 10 cm3 of wet sediment, was cut off from each core and then was washed through a 0.074 mm opening sieve. Eighty-six specimens have been examined for foraminifers and ostracods (Fig. 2). Table of ostracods taxa and samples has been submitted to a Correspondence Analysis (CA) [1].
3 Results
Ostracod species are mainly represented by warm-water species. 143 species have been identified. The number of individuals and of species decreases from the open sea to the northern part of the bay. Ostracods have been found in 64 stations; they do not occur in most of the stations of the bay head, which is mainly occupied by the acidic water mass e. Nevertheless, in stations 64, 63, 42 and 34, ostracods species occur; only one specimen has been found in station 42.
A valid interpretation of a correspondence analysis is possible only for the first three factors (Fig. 3).
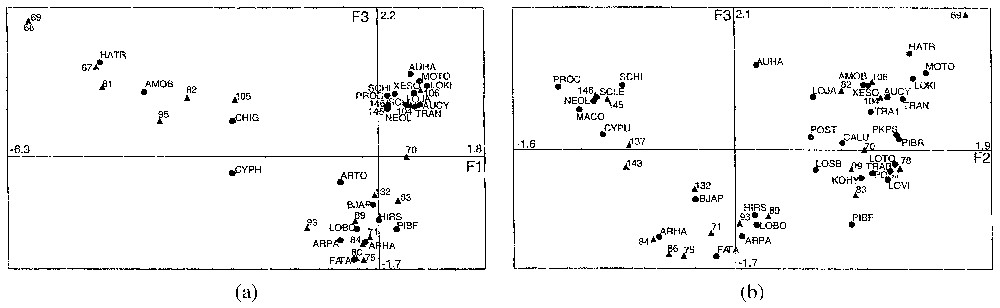
a. Plan factoriel F1–F3 de l'analyse des correspondances des échantillons et des stations. Les cercles noirs représentent les espèces et les triangles, les stations. Abréviations des noms d'espèces : AMOB : Ambtonia obai, ARHA : Argilloecia hanaii, ARPA : A. parallela, ARTO : A. toyamaensis, AUCY : Aurila cymba, AUHA : Aurila hataii, BJAP : Bradleya japonica, FATA : Falsobuntonia taiwanica, HATR : Hanaiborchella triangularis, HIRS : Hirsutocythere ? hanaii, CALU : Callistocythere undulatifacialis, CHIG : Chejudocythere higashikawai, CYPH : Cytheropteron hanaii, CYPU : Cytheropteron uchioi, KOHY : Kobayashina hyalinosa, LOBO : Lobocytheropteron ? donghaisensis, LOJA : Loxoconcha japonica, LOKI : L. kittanipponica, LOTO : L. tosaensis, LOVI : L. viva, LOSB : Loxoconcha sp. B, MACO : Macrocypris okinawae, MOTO : Moosella tomokae, NEOL : Neonesidea oligodentata, PKPS : Parakrithella pseudadonta, PIBF : Pistocythereis bradyformis, PIBR : Pistocythereis bradyi, POJA : Pontocythere japonica, POSJ : Pontocythere subjaponica, POST : P. subtriangularis, PROC : Propontocypris crocata, SCHI : Schizocythere kishinouyei, SCLE : Sclerochilus semirutrens, TRAB : Trachyleberis scabrocuneata, TRAN : Trachyleberis nitsumaii, TRA 1 : Trachyleberis sp. 1, XESO : Xestoleberis opalescenta. b. Plan factoriel F2–F3 de l'analyse des correspondances des échantillons et des stations. Les cercles noirs représentent les espèces et les triangles, les stations. Abréviations des noms d'espèces : voir Fig. 3a.
a. Correspondence analysis (CA) of the table of samples and stations. Representation of samples in the space of the F1–F3 axes (F1, F3). Black circles represent the species; triangles represent the stations. Abbreviations of the name of species: see above. b. Correspondence analysis (CA) of the table of samples and stations. Representation of samples in the space of the F2–F3 axes (F2, F3). Black circles represent the species; triangles represent the stations. Abbreviations of the name of species: see Fig. 3a.
Stations that contribute to F1 (67, 68, 69, 81, 82, 95 and 105) are under the influence of stagnant water masses b and d. In most stations (except 81), either volcanic ashes, or hot springs occur. Associated species are Hanaiborchella triangularis, Ambtonia obai, Cytheropteron hanaii and Chejudocythere higashikawai. In the Kagoshima Bay, the two former have been collected in the central basin, in stations under the influence of stagnant water mass b or d; the two latter can occur in stations under the influence of the water masses a or c.
Stations 104, 70, 106, 78, 83, and 99 contribute to F2 (Fig. 3b); all of them are located along the seashore of the bay. Most of them are under the influence of the low salinity water mass c; station 99, under the influence of the water mass a, is located in front of the outlet of a small river. Associated species are Callistocythere undulatifacialis, Xestoleberis opalescenta, Loxoconcha tosaensis, Aurila cymba, Loxoconcha japonica, L. kitanipponica, L. viva, Moosella tomokoae, Loxoconcha sp. B, Pistocythereis bradyi, Parakrithella pseudadonta, Pontocythere japonica, P. subtriangularis, Trachyleberis scabrocuneata, Kobayashina hyalinosa; they can occur in stations under the influence of water masses a, b, c, or d but all of them have been collected in stations under the influence of the low salinity water mass c. P. pseudadonta is never found under the influence of the open-sea water mass.
On the opposite side, open-sea stations (146, 145) and mouth stations (137 and 143) contribute to F2; the first three ones are under the influence of water mass a, the fourth under that of water masses a and b. Associated species are Schizocythere kishinouyei, Argilloecia toyamaensis, Neonesidea oligodentata, Sclerochilus semirutrens, Macrocypris okinawae, Cytheropteron uchioi, Propontocypris crocata.
Station 106, which is under the influence of water mass c, contributes to F3; associated species are Aurila hataii and Loxoconcha japonica, which are innerbay species. Stations 71, 75, 84, 86 89, 93 and 132 contribute also to F3; they are located either at the boundary of water masses b and c, or a and b, or under the influence of water mass d. Associated species are: Falsobuntonia taı̈wanica, Hirsutocythere? hanaii, Argilloecia parallela, Argilloecia hanaii, Bradleya japonica, Lobosocytheropteron donghaisensis; in Kagoshima Bay, they have been collected between 50 and 150 m deep.
4 Interpretation
Occurrence of several warm-water species among ostracod faunas indicates the influence of the Kuroshio Current.
Some of the species associated to stations 67, 68, 69, 81, 82, 95 and 105 can live in unfavourable environments for ostracods [11] as under stagnant water masses b and d; other ones are known on fine bottom sediments rich in organic matter [12].
All species associated to stations 70, 78, 83, 99, 104 and 106 are innerbay species and they can survive in low salinity environmental water and most of them occur on substrate where pellets are abundant as a consequence of the human pollution [14,15,22,23]. Benthonic foraminifers are abundant in stations 78, 83, 84 and 99: this is interpreted by the input of nutriments, as a result of the domestic activities. Buliminella elegantissima occurs in stations 104, 78, 70 and 83: it is usually mentioned in areas near the outfall of sewage [20].
Species associated to stations 145, 146, 143 and 137 occur usually in the open-sea environment, at the mouth of the bays and can be found in the bathyal environments [13,16,17]. Among these species, it is worthy of note that M. okinawae and N. oligodentata occur in the bay head area (station 34). In this part of the bay, ostracods occur only in four stations (34, 42, 63 and 64); the frequency of agglutinated foraminifers is low in stations 34, 63 and 64 and high in the eastern and northern parts [20]. This could attest the influence of the open-sea water entering through the Sakurajima Passage into the head environment, which is mainly occupied by acidic water mass e.
Species associated to stations 71, 75, 84, 86, 89, 93 and 132 are reported in the Japanese innerbays and also in the platforms; they can colonise the inner parts of the Bay from the open-sea environment, and particularly in sites under the influence of more or less stagnant water mass [2,3,8,10,18].
Stations and species are distributed in different sets (Fig. 3); in each of them, stations are under the influence of a mass of water or located at the limit of two water masses. In the Kagoshima Bay, the distribution of ostracods species is controlled by the circulation of the water masses; it allows the colonisation of some stations in the head bay area, mainly occupied by an acidic water mass. Dynamic, quality and structure of the water masses can be more important than the effects of volcanism. This latter can be drastic to the ostracods species by producing acidic water.
1 Introduction
En milieu lacustre, les modifications de la composition chimique de l'eau en relation avec l'activité volcanique déterminent la structure des paléobiocénoses d'ostracodes [7]. Les travaux concernant l'influence du volcanisme sur les microfaunes benthiques en milieu marin ont surtout intéressé les foraminifères [4,5,9,20].
Dans la baie de Kagoshima, située à l'extrémité méridionale de l'ı̂le de Kyushu (Japon), les ostracodes sont associés aux foraminifères. Cette baie s'étend entre la péninsule de Satsuma à l'ouest et d'Osumi à l'est (Fig. 1) sur une longueur de 75 km et une largeur de 25 km. La profondeur maximale est de 230 m, au centre de la baie [20]. Son originalité réside dans l'activité quotidienne du volcan Sakurajima.
Différentes unités géologiques ont été reconnues dans la partie méridionale de l'ı̂le de Kyushu. Le substratum est constitué de roches sédimentaires d'âges Mésozoı̈que et Paléogène, dans lesquelles un massif granitique, daté de 12 à 16 Ma, est intrusif. Ce substratum est recouvert, sur environ 2500 m, par d'importants dépôts d'ignimbrites (datées ±2,9 Ma) et par des pyroclastites [6]. Cette activité volcanique a débuté pendant le Néogène.
La baie de Kagoshima [19] trouve son origine dans la formation de deux caldeiras, celle de Aira dans sa partie la plus septentrionale et celle de Ata, dans sa partie méridionale. Le volcan Sakurajima appartient à l'arc insulaire du Japon du Sud-Ouest et se situe à la convergence des plaques Philippines et Eurasie ; il se dresse sur la bordure méridionale de la caldeira de Aira et, depuis l'éruption de 1914, il divise la baie en deux parties : lors de cet événement, une importante coulée de lave a obstrué le chenal compris entre l'ı̂le de Sakurajima et la péninsule d'Otsumi (Fig. 1).
En s'appuyant sur les caractéristiques physiographiques de la baie [21], il est possible de la subdiviser en trois parties principales (Fig. 1) :
- – l'embouchure de la baie, qui correspond à un chenal dont la largeur est quasiment constante et dont la profondeur est au maximum de 100 m ; sur sa bordure occidentale, des sources hydrothermales ont été découvertes ;
- – la partie centrale de la baie, qui occupe un bassin d'une profondeur d'environ 230 m et s'approfondit légèrement du sud vers le nord ;
- – le fond de la baie, au nord de l'ı̂le de Sakurajima, est séparé du bassin central par un étroit chenal, compris entre la base du volcan et la péninsule de Satsuma.
Dans la baie de Kagoshima, la température de l'eau diminue avec la profondeur ; au-delà de 150 m, elle est relativement stable au cours de l'année et est comprise entre 14 et 18 °C dans l'ensemble de la baie. Dans la partie septentrionale et centrale, elle est d'environ 20 °C à 100 m de profondeur et les différences sont minimes entre l'hiver et l'été. En revanche, de 0 à 75 m de profondeur, les fluctuations saisonnières sont beaucoup plus importantes (14,2 °C à 30 °C). À l'entrée de la baie, les valeurs sont beaucoup plus stables.
Au-dessous de 150 m, la salinité de l'eau est stable au cours de l'année ; elle est comprise entre 33,6 et 34,8‰ dans l'ensemble de la baie. En revanche, au-dessus de 25 m de profondeur, des fluctuations saisonnières sont enregistrées au cours de l'année : en hiver, de 34 à 34,9‰ à l'entrée de la baie et 33 à 34,3‰ dans la partie septentrionale et en été de 31,2 à 34‰ pour la première et de 23,3 à 32,7‰ pour la seconde. La salinité de l'eau tend donc à diminuer depuis l'entrée de la baie vers sa partie la plus septentrionale.
La répartition des températures de surface, de la salinité, de la transparence de l'eau, des sédiments sur le fond de la baie, la topographie sous-marine ont permis de mettre en évidence l'existence de courants de surface, à l'entrée de la baie et dans sa partie centrale, ainsi que de masses d'eaux stagnantes dans les parties centrale et septentrionale.
L'ensemble de ces données permet de différencier plusieurs masses d'eau [20] à l'intérieur de la baie (Fig. 2) :
- – une masse d'eau a : elle correspond à une branche du courant Kuroshio et pénètre à l'intérieur de la baie le long de la péninsule d'Otsuma, la traverse dans la partie septentrionale du bassin central et va heurter la péninsule de Satsuma ; il en résulte la formation d'un fort courant circulaire au nord du bassin central ; un diverticule de cette masse d'eau pénètre dans la partie la plus septentrionale de la baie, à la faveur du chenal existant entre le volcan et la péninsule de Satsuma ;
- – une masse d'eau c : elle correspond à l'eau de la baie qui a subi une légère dessalure, consécutivement à l'apport des rivières se déversant le long du littoral ; elle est donc enrichie en nutriments, résultat des rejets anthropiques consécutifs aux activités agricoles, urbaines et industrielles développées autour de la baie ; cette masse d'eau circule le long de la péninsule de Satsuma ; elle est obligée de changer de direction lorsqu'elle se heurte aux installations de Kire-Cho, gagnées sur la mer ; une branche de cette masse d'eau se dirige alors vers la péninsule d'Otsuma, en traversant le bassin central dans sa partie méridionale ;
- – deux masses d'eau stagnantes, b et d, occupant les parties les plus profondes du bassin central ;
- – une masse d'eau stagnante et acide, e, s'étendant dans la partie la plus septentrionale de la baie.
L'importance de ces différentes masses d'eau fluctue selon les saisons. La limite entre deux masses d'eau n'est pas stable au cours de l'année.
2 Matériel et méthodes d'étude
Des échantillons de sédiment ont été récoltés à l'aide d'un carottier de type Phleger, par Oki, de 1972 à 1978, au niveau de 146 stations situées à l'intérieur de la baie et de trois autres localisées en pleine mer (Fig. 2). Le centimètre supérieur de chaque sondage a été recueilli, soit environ 10 cm3 de sédiment. Ce volume a été lavé sur un tamis de 74 μm, puis séché et trié. Quatre-vingt-six échantillons ont été examinés pour les foraminifères et les ostracodes. En ce qui concerne l'ostracofaune, pour chaque station, le nombre d'espèces et le nombre d'individus recueillis ont été comptés. Les résultats ont été traités à l'aide d'une analyse des correspondances [1].
3 Résultats
Les ostracodes ont été trouvés dans 64 stations. De façon générale, les espèces recueillies sont caractéristiques de masses d'eau chaude. L'ostracofaune est peu abondante et peu diversifiée. Au total, 143 espèces ont été dénombrées. L'abondance et la richesse spécifique sont très variables d'une station à l'autre. La station 146 a livré 304 individus et 43 espèces et la station 42, seulement un individu. Trente-cinq espèces sont représentées par un seul individu et 59 par moins de cinq. Les stations de mer ouverte sont les plus riches en nombre d'individus et en nombre d'espèces. Ces derniers diminuent vers le bassin central et la partie septentrionale de la baie. Dans cette dernière, quatre stations seulement (64, 63, 42 et 34) ont livré des ostracodes ; les espèces rencontrées dénotent des influences océaniques.
Le traitement du tableau de dénombrement des espèces et des individus dans chaque station par l'analyse des correspondances nous a conduits à retenir les trois premiers facteurs.
Les stations 67, 68, 69, 81, 82, 95 et 105 contribuent à l'axe 1 (Fig. 3a). Elles sont situées dans la partie centrale de la baie et sont sous l'influence de la masse d'eau d [20]. Des sources hydrothermales sont connues dans les stations 67, 68, 69 et 105 et des cendres volcaniques sont signalées dans les sédiments des sites 82 et 95. Les espèces associées, pour l'ensemble de la baie, ont été rencontrées entre 20 et 220 m ; il s'agit de Hanaiborchella triangularis, Ambtonia obai, Cytheropteron hanaii, Chejudocythere higashikawai. Dans la baie de Kagoshima, les deux premières espèces ont été récoltées dans le bassin central, dans des stations sous l'influence des masses d'eau b et/ou d ; les deux autres espèces peuvent aussi se rencontrer dans des stations pouvant être sous l'influence des masses d'eau a ou c.
Les stations 104, 70, 106, 78, 99 et 83 contribuent à l'axe 2 (Fig. 3b). Elles sont toutes localisées le long du littoral de la baie et sont sous l'influence de la masse d'eau c, hormis la station 99 ; cette dernière, sous l'influence de la masse d'eau a, se situe au droit du débouché d'une petite rivière. Dans la baie de Kagoshima, les espèces associées peuvent occasionnellement se rencontrer dans des stations plus profondes, en pleine mer, à l'entrée de la baie et dans le bassin central et sous l'influence des masses d'eau a, b ou d. Il s'agit de : Callistocythere undulatifacialis, Xestoleberis opalescenta, Loxoconcha tosaensis, Aurila cymba, Loxoconcha japonica, L. kitanipponica, L. viva, Moosella tomokoae, Loxoconcha sp. B, Pistocythereis bradyi, Parakrithella pseudadonta, Pontocythere japonica, P. subtriangularis, Trachyleberis scabrocuneata, Kobayashina hyalinosa. La plupart d'entre elles ont été collectées uniquement dans des stations sous l'influence de la masse d'eau c.
En position opposée sur le même diagramme, les stations situées en pleine mer, 146 et 145 et celles de l'entrée de la baie, 137 et 143 contribuent également à l'axe 2. Les trois premières sont sous l'influence de la masse d'eau a, la quatrième se situe à la limite des masses d'eau a et b. Les espèces associées sont Schizocythere kishinouyei, Argilloecia toyamaensis, Neonesidea oligodentata, Sclerochilus semirutrens, Macrocypris okinawae, Cytheropteron uchioi, Propontocypris crocata ; il faut souligner que M. okinawae et N. oligodentata sont présentes dans la partie la plus septentrionale de la baie (station 34), occupée quasi totalement par la masse d'eau acide e.
Deux ensembles s'opposent donc sur l'axe 2 : l'un regroupe les stations et les espèces de pleine mer, le second, celles de l'intérieur de la baie.
La station 106 contribue au seul axe 3 (Fig. 3b). Les espèces associées sont Aurila hataii et Loxoconcha japonica ; l'une et l'autre sont connues à l'intérieur des baies japonaises. À l'opposé, les stations 71, 75, 89, situées à la limite des masses d'eau b et c, 132, à la limite des masses d'eau a et b, 84, 86 et 93, sous l'influence de la masse d'eau d, contribuent aussi à F3. Les espèces associées sont Falsobuntonia taı̈wanica, Hirsutocythere ? hanaii, Argilloecia parallela, Argilloecia hanaii, Bradleya japonica, Lobosocytheropteron donghaisensis. Dans la baie de Kagoshima, elles sont présentes dans des stations situées entre 50 et 150 m.
4 Interprétation
L'ostracofaune de la baie de Kagoshima comporte essentiellement des espèces d'eau chaude ; ceci dénote l'influence du courant Kuroshio, dont une branche pénètre à l'intérieur de la baie.
Dans les stations situées à l'entrée de la baie, les populations sont abondantes et diversifiées ; le nombre d'espèces et le nombre d'individus diminuent depuis l'entrée de la baie jusque vers le bassin central et la partie la plus septentrionale, où la plupart des stations sont stériles. Cet environnement est essentiellement occupé par la masse d'eau e, stagnante, acide, conséquence de l'activité volcanique du Sakurajima ; de nombreuses sources hydrothermales y ont été répertoriées : la dissolution dans l'eau du dioxyde de carbone émis par les fumerolles a pour effet de faire baisser le pH du milieu de vie. Celui-ci peut devenir hostile au développement des organismes à test carbonaté. L'effet nocif de l'acidité de l'eau est particulièrement bien marqué, puisque, dans cette partie de la baie, quatre stations seulement ont livré des ostracodes. Les populations de Foraminifères, quant à elles, sont dominées par les agglutinants dans les stations orientales et nord-occidentales ; en revanche, les stations 34, 63 et 64 sont caractérisées par la faible fréquence de ces derniers [20].
Les espèces H. triangularis, A. obai, C. hanaii, C. higashikawai sont associées aux stations 67, 68, 69, 82, 95, 105, où des indices de volcanisme sont connus, mais aussi à la station 81, où ni cendres volcaniques, ni activité hydrothermale n'ont été signalées. Toutes ces stations, dont la bathymétrie est comprise entre 97 et 220 m, sont sous l'influence de la masse d'eau d. A. obai est citée dans des milieux défavorables aux ostracodes [11]. C. hanaii et C. higashikawai sont rencontrés sur des substrats riches en matière organique [12]. Ces espèces s'adapteraient à des environnements profonds, déficitaires en oxygène ; dans la baie de Kagoshima, elles caractérisent les stations sous l'influence de la masse d'eau stagnante b ou d.
Dans la station 104, des sources hydrothermales ont été signalées. Avec les stations 70, 106, 78, 83 et 99, où aucun indice d'activité volcanique n'est connu, elle est associée à des espèces connues dans l'environnement littoral ; toutes sont rencontrées à l'intérieur des baies japonaises et, la plupart du temps, à moins de 150 m de profondeur ; quelques-unes sont épiphytes. Elles peuvent tolérer une légère dessalure de l'eau du milieu de vie se rencontrant souvent sur des substrats riches en matière organique où abondent les pellets, indice de pollution d'origine anthropique [14,15,22,23]. Les milieux de vie, hormis la station 99, dans la baie de Kagoshima, sont tous sous l'influence de la masse d'eau, c, dessalée, polluée ; la station 99, se situe au débouché d'un petit cours d'eau, vecteur de rejets d'origine anthropique. Dans les stations 78, 83, 84 et 99, l'apport de nutriments d'origine anthropique expliquerait l'abondance élevée des foraminifères benthiques [20] ; Buliminella elegantissima souvent citée au droit des émissaires anthropiques est présente dans les stations 104, 70, 78 et 83.
Ni sources hydrothermales, ni cendres volcaniques ne sont connues dans les stations sous l'influence de la masse d'eau a. Aux stations de pleine mer 146, 145 comme à celles 137 et 143 de l'entrée de la baie sont associées des espèces habituellement citées dans les environnements de pleine mer, à l'entrée des baies japonaises, voire dans l'environnement bathyal [2,3,8,10,18] ; certaines pourraient s'adapter aux conditions de milieu de vie des baies japonaises. La présence de M. okinawae et N. oligodentata dans la partie la plus septentrionale de la baie implique que l'eau d'origine océanique y pénètre à la faveur du passage compris entre le volcan et la péninsule de Satsuma.
Aucun indice de volcanisme n'a été décelé au niveau des stations 71, 75, 84, 86, 89, 93 et 132, qui lui sont associées. Ces sept stations se situent dans un intervalle bathymétrique compris entre 88 et 222 m. Elles sont localisées à la limite, soit des masses d'eau a et b, soit b et c, ou sont sous l'influence de la masse d'eau d, donc de masses d'eau plus ou moins stagnantes, se déplaçant latéralement au cours de l'année. Les espèces associées sont connues dans les environnements de plate-forme, mais aussi à l'intérieur des baies japonaises [13,16,17] ; dans la baie de Kagoshima, elles s'adapteraient à des environnements plus ou moins déficitaires en oxygène, voire instables.
Dans la baie de Kagoshima, la dissémination des espèces d'ostracodes et donc leur distribution est assurée par la structure, la qualité et la dynamique des masses d'eau. Leur influence peut gommer celle du volcanisme. Ce dernier se révèle néfaste au développement des ostracodes, lorsqu'il se traduit par l'émission d'eaux acides, le milieu de vie devenant hostile aux organismes benthiques à test carbonaté.