

Abridged version
1 Introduction
At a global scale and compared to Early Bajocian times [22], Bathonian and, at a lesser extent, Late Bajocian, appear as poorly favourable periods for reefal development. Fig. 1, which is based on bibliographical references [1,6,7,9–13,15,16,18–21,23,26,27,33–38], reveals that they are distributed according to two subtropical belts at fairly high latitude, separated by an equatorial belt without coral reefs. In the southern belt, two sets of outcrops have been given a prime importance in the knowledge we have on corals of these times: Kachchh in India and Madagascar.

Les milieux coralliens connus dans l'intervalle Bajocien supérieur–Bathonien. Fond paléogéographique et flèches de courants de Haq et Van Eysinga [17].
Corallian meadows known from the Upper Bajocian–Bathonian interval. Palaeogeographic map and current arrows from Haq and Van Eysinga [17].
The study of Kachchh has been undertaken by Gregory [15] and recently reconsidered for taxonomy [28,30] as well as for ecology [14,29]. The work [14] describes coral beds, and not reefs, where a large diversity of colonial genera were living in a low energy, poorly lit environment, subjected to a moderate sedimentation and controlled by a soft substrate.
This paucity of true reefs seems to be so representative of Bathonian corallian sediments that the descriptions of Madagascan outcrops given by Collignon [9] and Besairie [8] appear as an obvious exception. These authors evoke some thirty coral reefs that represent the remains of a kind of great barrier ageing from Upper Bajocian to Upper Bathonian, extending over two degrees of latitude and exceeding 10 km in width in some places. Atolls are also described. The study of corals from these ‘reefs’ has been undertaken by Alloiteau [2]. Despite his very typological approach, the fauna from Madagascar appears highly diverse and has been used as a basis for the biostratigraphic diagnostic of Bathonian [5], when the exact limit between Bajocian and Bathonian remains unknown in this area.
A field mission in the Betioky area allowed us to see again nearly all sites described by Collignon [9] (Fig. 2). Some outcrops are presented here in order to help the reassessment of these coral formations.

Localisation des affleurements (flèches). Fond géologique adapté de Betioky [39].
Location of outcrops (arrows). Geological background adapted from Betioky [39].
2 Geological framework
The studied area belongs to the southern part of the Morondava Basin. This basin is characterised by a Liassic extensional tectonics (‘Karoo rifting’) and by normal and strike-slip faulting during the drifting of the island from the Dogger to the Early Cretaceous [25,32]. The coral outcrops of the Betioky area occur according to a belt oriented north–south, with a gentle regional dip to the west. In detail, thicknesses are difficult to establish because of faults and discontinuity of outcrops. The Betioky map [39] shows a first eastern belt described as Bajocian-Bathonian coral limestones and a second western belt described as Upper Bathonian sandstones. The Callovian is restricted in the Ranonda area. Because of normal faults, the transition to Upper Jurassic cannot be seen. Within the Middle Jurassic, Collignon [9] distinguished 4 coral beds, the third one being the ‘Anabacia’ bed. Besairie [8, pl. 81] pointed out 3 coral limestone beds under this bed.
From the structural sketch of Besairie [8], it can be seen that this area presents the deepest basement (7000 m) in Southern Madagascar; this leads us to consider a strong subsidence.
3 The section of Ankotoboka
In opposition to the regional dip, this section (Fig. 3) shows a dip of some 15° to northeast. More calcareous beds are extremely thin and separated by large areas where the rock does not crop out but where the ground is covered by post-Jurassic ferruginous alterites. Observation of transitions between coral beds or oolitic beds and the surrounding sediments, as well as the base of the section outcropping over several meters, lead us to think that most of the section is made of clay and sandstone with micaceous debris. Most calcareous beds are either oolitic or oncolitic. Corals occur in some beds and the composition of the coral assemblage is specific to beds. So, a probably monospecific assemblage of massive tridimensional cerioid colonies poorly bioeroded are found in the first bed (BET 2). The largest colony reaches a diameter of 53 cm. Laterally, it passes to quartz and iron ooids facies cemented by dolosparite. Beds BET 5 and 6 are quite different and give only small colonies belonging to 8 genera. The ooids are often built around a quartz nucleus. Coral colonies appear put on the ground, free of sediment, rarely joined together, poorly eroded; their autochtony is quite plausible.
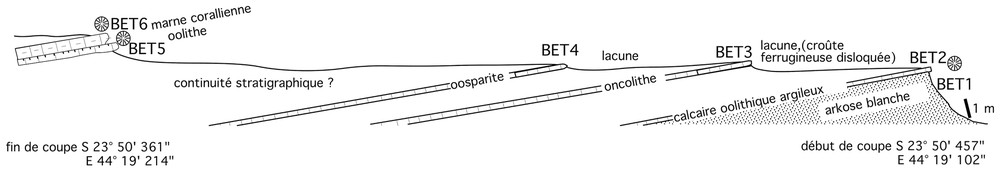
La coupe d'Ankotoboka.
Section of Ankotoboka.
4 The section of the first cuesta of the Dogger between Betioky and Ranonda
Coming from Betioky, the first cuesta oriented north–south is incised by an east-west river that allows us to observe partially the succession (Fig. 4). First, the road goes across white sandstones and then arrives to the base of the section (Fig. 4, GPS coordinates: S23° 42 278–E 44° 20 488). Here, the river allows us to observe the first calcareous facies of the Dogger.

La coupe de la première cuesta du Jurassique moyen entre Besavoa et Betioky.
Section of the first Middle Jurassic cuesta between Besavoa and Betioky.
The front and the back slope of the cuesta are mapped as coral limestone. Collignon [9] points out them as his first coral ‘bed’ (“premier niveau à coraux”) and attributed them to Upper Bajocian or Bathonian. Brachiopods have been collected and determined as Burmirhynchia termierae Rousselle and Baeorhynchia transversa Cooper. They indicate the top of Lower Bajocian (Humphriesianum Zone) and early Upper Bajocian (Subfurcatum Zone); see [3] (p. 320), [4] (pl. 1, Figs. 2 and 3). A preliminary comparison of coral faunas with recently revised faunas of the Humphriesianum zone [22] gives more credibility to an Upper Bajocian assignation. The very close outcrop S23° 42 424–E44° 20 714, which is stratigraphically situated below, has provided an intraformational pebble, with a palynological content. Six Nannoceratopsis sp. iculata Stover and three N. gracilis Alberti indicate a Toarcian–Upper Bathonian (Discus Zone excluded) global age, with a more probable Bajocian age.
The section crops out very discontinuously (Fig. 3). The most obvious facies are the most calcareous, but they represent a very small thickness compared to siliciclastic and clayey sediments. Basal limestones include mudstones with bird-eyes and oolitic facies. Clay-sandstone facies show some beds that furnished some bivalves and gastropods as internal molds. An internal mold of Nerineid has been found and indicates the marine depositional environment. The small topographic scarp of the lower half of the front is characterised by the occurrence of sparse, dome shaped middle size (ca 10 cm) colonies; they are well shaped, poorly bioeroded and loosely embedded in an oolitic sediment. The top of the cuesta is made of a true limestone rich in poorly preserved brachiopods (Burmirhynchia sp. and Monsardithyris sp.), corals and furnishes the quite rare occasion to observe coral facies in vertical section. Corals occur in rudstone/floatstone with an oolitic matrix. At the spot 23° 42 134–44° 20 599, tri-dimensional colonies, reaching a 29 cm diameter, can be observed. Diversity and bioerosion are low; the preservation is rather poor.
Depositional environments at the bottom are inner marine calm meadows marked by emersion (bird-eyes) or more agitated but always very shallow (limestone ooids). The environments corresponding to clay sandstones facies are marine. Finally, the deposit of limestone at the top is probably related to storms capable to re-sediment large colonies. These three-dimensional colonies have grown in a well-lit environment; the fairly regular shape of these colonies indicates more a loose pavement of coral living on a sandy substrate rather than a reefal origin.
5 Discussion
These observations lead to re-consider the general organisation of these deposits. The section of Ankotoboka had been previously thought as an atoll [8,9]. We think that this interpretation depends on the isochrony that has been admitted between BET 2 and BET 5–6. This isochrony is not geometrically admissible because the monoclinal structure implies a superposition of these beds. Nevertheless, a tiny doubt could remain due to observation gaps. Facies analysis removes the doubt, biofacies and lithofacies being different among beds. The setting is neither an atoll, nor a boundstone. Another supposed atoll had been evoked by Collignon [9] near Mikaikarivo. It has been observed and comes under the same re-assessments.
The view of a great barrier extending over two degrees of latitude and wider than 10 km is no longer acceptable. It is only a belt of outcrops including some coral facies that are neither bioherms nor biostromes, but thin coral beds. The section of the path from Betioky to Ranonda shows a fairly high density of corals that does not correspond to the framework of a true boundstone, but mainly to rudstone deposits controlled by storm wave action. We could consider that corals come from the destruction of reefs; however, the corals generally display the regular shape known for corals living in loose pavements on a sandy substrate.
6 Conclusions
Early corallian sediments of the Dogger in the Betioky area are related to early Upper Bajocian. They do correspond neither to a great barrier nor to atolls, but to sparse and thin limestone beds in a basically siliciclastic context. Mainly colonial corals grew up to a moderate size, often in a sparse fashion on a sandy substrate. This carbonate sedimentation originated in brief episodes of high relative sea level or of particular climates in an extensive, strongly subsiding, structural context, and a palaeogeographic frame dominated by the vicinity of a continent capable of supplying the high terrigenous input.
1 Introduction
À l'échelle de notre planète (Fig. 1), le Bathonien et, à un moindre degré, le Bajocien supérieur apparaissent comme des époques assez peu favorables au développement des récifs coralliens. Le développement des récifs du Bajocien inférieur récemment étudiés [22] apparaı̂t ensuite fortement interrompu, ou au moins ralenti. C'est sans doute pour cette raison que les coraux de ces périodes sont encore très mal connus. Les milieux coralliens apparaissent répartis en deux ceintures d'assez hautes latitudes, séparées par une absence de ces environnements autour de l'équateur. La littérature qui décrit les coraux susceptibles de construire des récifs fait apparaı̂tre quelques gisements en Europe de l'Ouest, où ils ont sans doute été davantage prospectés qu'ailleurs : en Angleterre [26] avec, en particulier, le riche gisement de Fairford [27], en France à Saint-Gaultier [6,20], en Normandie, en Provence [21] et en Suisse [19]. En Europe orientale, on connaı̂t le gisement oolithique polonais de Balin [7] et des affleurements, probablement bathoniens, en Roumanie [10]. Les véritables récifs du Maroc [1,35], considérés comme bajociens ou bathoniens, représentent plutôt l'équivalent stratigraphique des récifs algériens de la base du Bajocien supérieur [12,34]. Des coraux assez mal datés, mais probablement bathoniens ou calloviens, sont cités par Flügel [13], en Iran. Les coraux décrits récemment par Pandey et al. [31] en Jordanie sont surtout calloviens (deux échantillons bathoniens, dont un solitaire). Un âge Bathonien est proposé pour les faunes du Kenya [16,37,38], de Tanzanie [33] et du Tibet [23,24] ; He [18] propose un âge Bajocien–Bathonien pour d'autres faunes du Tibet. Une petite partie des faunes mésozoı̈ques décrites au Japon [11] pourrait se rapporter à cet intervalle [36].
Dans la ceinture sud, les deux gisements de Kachchh, en Inde, et de Madagascar apparaissent fondamentaux pour la connaissance que nous avons des coraux de cette époque.
L'étude du gisement de Kachchh, entreprise par Gregory en 1900 [15], a été reprise récemment, sur le plan tant taxinomique [28,30] qu'écologique [14,29]. Ces travaux nous livrent l'image de bancs coralliens et non de récifs où vivait une large diversité de genres coloniaux, dans un environnement de faible énergie, peu éclairé, soumis à un taux de sédimentation modérée et largement contraint par les caractéristiques du substrat mou.
À en juger par la littérature, cette image des milieux coralliens indiens dépourvus de vrais récifs semble assez représentative et les cas rarissimes de récifs bathoniens cités sont toujours petits. En cela, les descriptions faites par Collignon [9] et appuyées par Besairie [8] apparaissent comme une exception flagrante, dans la mesure où ces auteurs évoquent une trentaine de récifs, qui représentent le reste d'une sorte de « grande barrière » corallienne, d'âge Bajocien supérieur à Bathonien supérieur, s'étendant sur 2 degrés de latitude et dépassant par endroits 10 km de large. Ces auteurs évoquent également la présence d'atolls. Sur le plan taxinomique, les coraux issus de ces formations ont été étudiés par Alloiteau [2], qui décrit 73 espèces, dont 50 sont formellement reconnues comme nouvelles. Parmi ces espèces nouvelles, 72 % sont définies sur un ou deux spécimens, ce qui caractérise une démarche typologique dont les excès ont certainement conduit à une pulvérisation outrancière des taxons. Néanmoins, ce travail ainsi que les formes récoltées non décrites par cet auteur laissent envisager une diversité réelle non négligeable et tout à fait compatible avec la paléolatitude proche de 30°S, envisagée pour ces gisements (Fig. 1). Ces premières descriptions ont servi de base à la caractérisation du Bathonien [5] à partir des coraux, alors même que l'attribution stratigraphique des différents niveaux à coraux reste incertaine, en particulier au niveau de la limite Bajocien–Bathonien, encore mal située.
Une mission de terrain dans la région de Betioky (Fig. 2) a permis de revoir la quasi-totalité des sites décrits par Collignon ([9] Fig. 2). Quelques affleurements sont présentés ici pour appuyer la réinterprétation de ces formations à coraux.
2 Cadre géologique
La région étudiée appartient à la partie sud du bassin de Morondava. Ce bassin est marqué par une structuration liasique en extension (Karoo rifting) et par le rejeu normal et décrochant des accidents durant la dérive de l'ı̂le, depuis le Dogger jusqu'au Crétacé inférieur [25,32]. Les affleurements corallifères de la région de Betioky se présentent cartographiquement suivant une bande nord–sud de structure monoclinale à léger pendage ouest. Dans le détail, l'ensemble est faillé et les affleurements sont assez discontinus, de telle sorte qu'il n'est pas possible d'évaluer précisément les épaisseurs. La carte de Betioky au [39] distingue une première bande orientale, rapportée à des calcaires à polypiers bajociens à bathoniens, et une deuxième bande gréseuse, attribuée au Bathonien supérieur. Le Callovien y est limité au secteur de Ranonda. La transition avec le Jurassique supérieur est invisible, un ensemble de failles mettant en contact le Dogger et le Crétacé. À l'intérieur de cet ensemble du Jurassique moyen, Collignon [9] distingue quatre niveaux à polypiers, le troisième étant un banc à « Anabacia ». Besairie [8, (pl. 81)] évoque trois niveaux de calcaires à polypiers sous ce banc.
Le schéma structural de Besairie [8] indique que le secteur étudié est, de tout le Sud malgache, celui qui présente le socle le plus profond (7000 m), ce qui laisse envisager une forte subsidence.
3 La coupe d'Ankotoboka
La coupe (Fig. 3) débute au point S23° 50 457′–E44° 19 102′ et se termine au point S23° 50 361′–E44° 19 214′ (coordonnées prises au GPS).
Cette coupe présente un pendage d'environ 15° vers le nord-est, ce qui va à l'encontre du pendage régional. Les niveaux calcaires sont extrêmement minces et séparés par des étendues où la roche n'est pas affleurante, mais où le sol est recouvert d'altérites ferrugineuses post-jurassiques. L'examen des transitions entre les bancs à coraux ou oolithiques et leur encaissant ainsi que la base de la série, visible sur plusieurs mètres, laissent penser que la roche qui constitue l'essentiel de la série est argilo-gréseuse, avec de nombreux débris micacés. Les niveaux plus calcaires présentent des faciès oolithiques ou oncolithiques. Les coraux apparaissent dans certains bancs et la composition des assemblages coralliens est différente suivant les bancs. Ainsi, le premier banc (BET 2) permet d'observer un peuplement probablement monospécifique de colonies cérioı̈des massives, tridimensionnelles, peu bioérodées, atteignant la taille maximale de 53 cm de diamètre. Latéralement, il passe à des faciès à grains de quartz et oolithes ferrugineuses, cimentés par de la dolosparite. Les niveaux BET 5 et 6 sont très différents, ne présentant que de petites colonies, appartenant à huit genres. Ils présentent des ooı̈des dont le nucléus est très souvent un grain de quartz.
Les colonies de coraux posées sur le sol apparaissent bien dégagées d'une gangue qui était nécessairement meuble, un grès à ciment dolomitique. Elles sont rarement jointives, n'apparaissent pas particulièrement usées ; leur autochtonie est donc tout à fait vraisemblable.
4 La coupe de la première cuesta du Dogger entre Betioky et Ranonda
En venant de Betioky, la première cuesta, de direction nord–sud, est entamée par une rivière orientée est–ouest, qui permet d'observer partiellement la succession (Fig. 4). La route traverse d'abord les grès blancs avant d'arriver au point de coordonnées GPS : S23° 42 278–E44° 20 488. Là, la rivière permet d'observer les premiers faciès calcaires du Dogger.
Le front et le revers de la cuesta sont cartographiés comme calcaire à polypiers (carte au ). Collignon [9] les rapporte à son premier niveau à coraux et au Bajocien supérieur ou Bathonien. Les Brachiopodes récemment collectés Burmirhynchia termierae Rousselle et Baeorhynchia transversa Cooper donnent un âge incluant le Bajocien inférieur élevé (Zone à Humphriesianum) et le Bajocien supérieur basal (Zone à Subfurcatum) [3] (p. 320), [4] (pl. 2, Figs. 2 et 3). La comparaison des faunes coralliennes avec celles récemment révisées de la Zone à Humphriesianum [22] accrédite un âge Bajocien supérieur plutôt que Bajocien inférieur.
L'affleurement très proche (S23° 42 424–E 44° 20 714), qui est situé stratigraphiquement au-dessous, a fourni un galet argileux intraformationnel, avec contenu palynologique. Six Nannoceratopsis spiculata Stover et trois N. gracilis Alberti indiquent un intervalle Toarcien–Bathonien supérieur (Zone à Discus exclue), avec un âge Bajocien plus probable.
La coupe apparaı̂t de façon très discontinue (Fig. 4). Les faciès les mieux affleurants sont les calcaires, qui ne représentent qu'une épaisseur très faible. L'essentiel de la série est argilo-gréseux. Les calcaires de la base montrent des faciès mudstone à fenestrae et des faciès oolithiques. Les faciès argilo-gréseux constituent des niveaux modérément fossilifères à bivalves et gastropodes sous forme de moules internes. Un moule interne de nérinée atteste le caractère marin de ces faciès. Le petit ressaut topographique de la moitié inférieure du front est marqué par la présence de colonies éparses de coraux en dômes, bien formées, de taille moyenne (de l'ordre de 10 cm), peu bioérodées dans un sédiment oolithique. Le sommet de cuesta est marqué par un vrai calcaire riche en brachiopodes assez mal conservés (Burmirhynchia sp., Monsardithyris sp.) et en coraux ; il fournit une des très rares occasions de voir ces calcaires à coraux en coupe. Les coraux se présentent en place dans un rudstone–floatstone à matrice oolithique. Au point 23° 42 134–44° 20 599, on observe des colonies tridimensionnelles, qui peuvent atteindre 29 cm de diamètre. La diversité est faible, les perforations peu nombreuses, la préservation assez mauvaise.
Les paléoenvironnements de la base sont des milieux marins internes, marqués par l'émersion (fenestrae) ou un peu plus agités mais toujours très peu profonds (oolithe calcaire). Les environnements correspondant à la partie argilo-gréseuse sont marins. Enfin, les environnements de dépôt des calcaires du sommet sont probablement liés à des tempêtes susceptibles de remanier des colonies d'assez grande taille. Par ailleurs, ces colonies tridimensionnelles ont vécu dans un environnement bien éclairé. La forme assez régulière de ces colonies indique plutôt une biocénose initiale non récifale, c'est-à-dire davantage un peuplement épars sur un fond granuleux bien éclairé.
5 Discussion
Ces observations permettent de repenser différemment l'organisation générale de ces dépôts. La coupe d'Ankotoboka avait été interprétée comme un atoll [8,9]. On peut penser que cette interprétation résulte de l'isochronie admise par ces auteurs entre les niveaux BET 2 et BET 5–6. Or, cette isochronie n'est pas admissible, parce que géométriquement la structure monoclinale oblige à considérer ces niveaux comme superposés. Un léger doute peut néanmoins subsister, du fait des lacunes d'observation. L'analyse faciologique suffit à lever ce doute, les assemblages coralliens et les lithofaciès étant différents. La structure n'a rien d'un atoll, ni même d'un calcaire construit. Un autre atoll supposé avait été évoqué par Collignon [9] dans le secteur de Mikaikarivo. Il a été observé et relève des mêmes ré-interprétations.
L'image d'une grande barrière s'étendant sur 2 degrés de latitude et dépassant par endroits 10 km de large n'est pas davantage acceptable. Il s'agit seulement d'une bande d'affleurements comprenant des faciès à coraux. Ces faciès à coraux ne sont généralement, ni biohermaux, ni même biostromaux, mais constituent des bancs à coraux peu épais, sans structure construite. La coupe de la route de Betioky à Ranonda montre en sommet de cuesta une densité de coraux assez forte. Il s'agit néanmoins de rudstone, indiquant plutôt une mise en place par dépôt de tempête. On pourrait envisager que ces coraux aient été produits par des vrais récifs. On peut aussi imaginer, avec une meilleure probabilité, que ces coraux proviennent de peuplements épars de milieux meubles, la forme régulière des coraux massifs évoquant plutôt la croissance sans contrainte observée habituellement dans ces environnements.
6 Conclusions
Les premiers sédiments coralliens du Dogger de la région de Betioky sont attribués au début du Bajocien supérieur. Ils ne correspondent, ni à une grande barrière récifale, ni à des atolls, mais à de rares bancs calcaires peu épais, dans un contexte fondamentalement silicoclastique. Les coraux y sont présents sous la forme de colonies, de taille souvent modeste, rarement agglomérées, avec une forme évoquant souvent une vie sur substrat meuble. Cette sédimentation carbonatée correspond à des épisodes brefs, pouvant être liés à de hauts niveaux marins relatifs ou à des périodes climatiques particulières dans un contexte structural extensif, à forte subsidence, dans lequel la proximité continentale explique le fort détritisme.
Remerciements
Ce travail a été réalisé grâce au concours financier que l'AUPELF UREF et le FICU ont apporté à la réalisation de la campagne de terrain.