

Abridged English version
1 Introduction
In the Moroccan Western Anti-Atlas (Fig. 1), the Lower-Cambrian deposits, exposed in the Issafène syncline (Fig. 2), show an alternation of carbonated and siliciclastic facies. Within carbonated facies, the vertical distribution of cyanobacterial microbialites structures and faunal archaeocyathes allow us to subdivide the succession in three units (Fig. 3A): the stromatolitic series, the thrombolitic series and the archaeocyathan series.
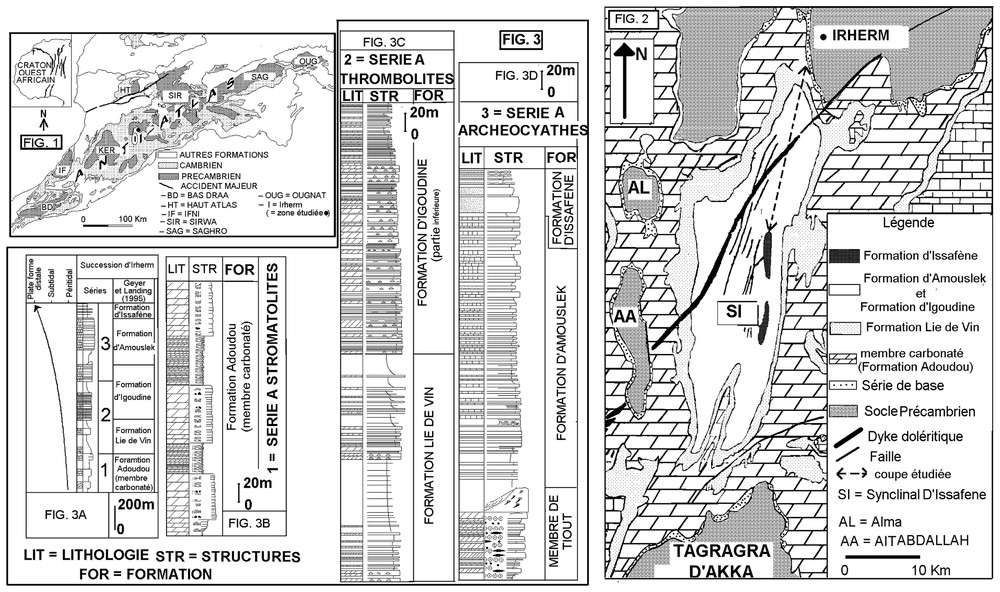
Schéma structural de l'Anti-Atlas et localisation de la région d'Irherm.
Anti-Atlasic sketch map showing the location of Irherm area.
Coupe effectuée dans le synclinal d'Issafène.
Sketch map of the Issafène syncline, showing the location of the studied section.
(A) Succession du Cambrien inférieur de la région d'Irherm. (B) Série à stromatolites. (C) Série à thrombolites. (D) Série à archéocyathes.
(A) Lower Cambrian Irherm succession. (B) Stromatolitic series. (C) Thrombolitic series. (D) Archaeocyathic series (D).
2 Stromatolitic series (Fig. 3B)
This series, which corresponds to the lower member of the Adoudou Formation [7], represents a long period in which a large area of the Anti-Atlasic platform was occupied by peritidal cyanobacterial mats. The stromatolites, locally interlayered with silicified and brechified evaporitic layers, are mainly domal and planar-shaped.
Examination of stromatolitic internal structures shows alternating layers of dark dolomicrosparite and light dolosparite. The dolomicrosparite organic-rich layers resulted from a diagenetic degradation of cyanobacterial organic laminae. The genesis of these stromatolitic organic laminae is related to non-calcified cyanobacteries in a peritidal zone, as indicated by the associated evaporitic and silicified facies. The associated light dolosparite layers are probably of sedimentary origin. As in the Lagoa Vermelha in Brazil [19] and in the Australian Coroong Lagoon [4], we suggest that dolomite is a primary feature in these layers. Precipitation of micritic dolomite is realised by the decomposition of an organic microbial substrate under anoxic and hypersaline conditions.
Lateral branching of cyanobacterial filaments leads to the formation of extensive laminae and the vertical stacking of these forms allows us to built the stromatolitic facies. These features were probably appropriate growth forms of the soft and fragile microbialites under the littoral hydrodynamic conditions (Fig. 4).
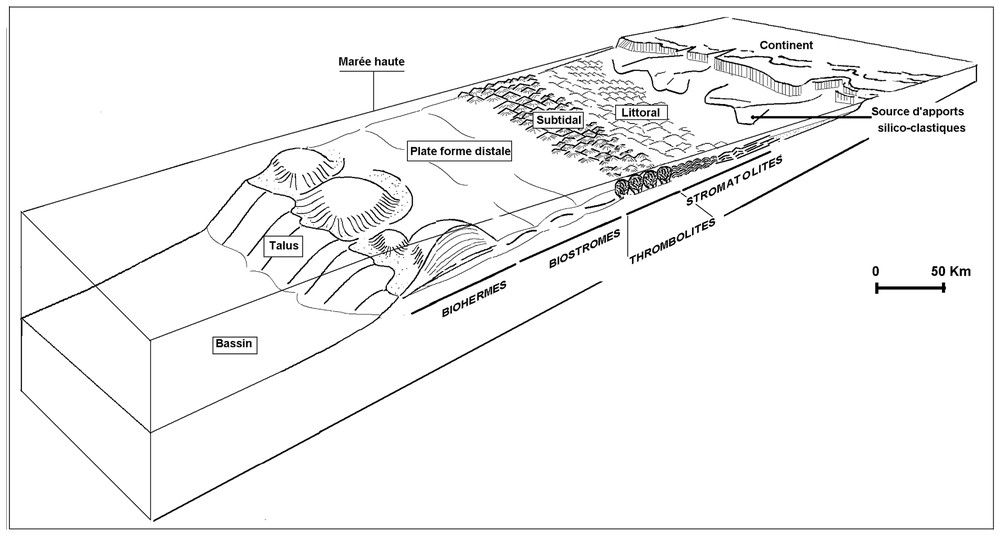
Zonation bathymétrique des microbialites et des faciès associés de la succession cambrienne d'Irherm.
Palaeo-bathymetric zonation of the Lower-Cambrian western Anti-Atlasic microbialites.
3 Thrombolitic series (Fig. 3C)
This series corresponds to the ‘Lie-de-Vin’ Formation and the lower part of the Igoudine Formation [7]. Its base is marked by the first development of thrombolitic facies.
Thrombolitic facies resulted from lateral juxtaposition of individual thrombolitic bioherm to form metric scale beds. The internal structure of each thrombolite shows a dark clotted micritic fabric separated by small and irregular cavities, occasionally filled with clastic sediments and sparitic cements.
The organic-rich micritic part of this structure was probably edified by coccoidal shaped calcimicrobes, secreting an acidic mucilaginous substance, as in modern lacustrine thrombolites [1]. Diagenetic alteration of this non-calcified mucus, by the activity of hetertrophic bacteria [1], determines the void-filling clastic sediment or dolomitic cement.
The vertical development of calcimicrobes within thrombolitic biohermes is assured by increasing water depth from stromatolitic and evaporitic supratidal zone to subtidal thrombolitic environment (Fig. 4) and by the first appearance of calcified and mucilaginous cyanobacteria in the Lower-Cambrian Anti-Atlasic platform, as suggested by Latham and Riding [11].
4 Archaeocyatic series (Fig. 3D)
This series correspond to the Tiout Member, Amouslek Formation and Issafène Formation [7]. Dendritic calcimicrobes constitute the typical microbialite forms in this fossiliferous part of the Lower-Cambrian succession (archaeocyathan series). Carbonated facies are dominated by archaeocyathic and calcified microbialite offshore reefal build-up. Archaeocyathes, which constitute an abundant faunal constituent, are of bafflestone type. Their branching framework seems to be the necessary substrate for dendritic microbialite development. These epibionte calcimicrobes are attached to the sidewalls of cavities formed by this archaeocyathan branching framework. Reworked calcimicrobe and other skeletal debris contribute significantly to interbiohermal and adjacent shoal grainstones formation. Occurrence of this offshore reefal build-up (Fig. 4), just over the subtidal thrombolitic series, is an evidence of a second episode of sea level rise.
5 Conclusion
In the western Anti-Atlas, the Lower-Cambrian succession is subdivided in three lithostratigraphic units characterised, each one, by a particular microbialitic build-ups. The vertical distribution of these microbialitic structures is probably related to major marine ecosystem modifications. The stromatolitic series represents a long period in which a large area of the Anti-Atlasic platform was occupied by peritidal cyanobacterial mats. Laterally branched cyanobacterial filaments were appropriate growth forms of laminated microbialites under peritidal hydrodynamic conditions. In the overlaying series, abrupt occurrence of clotted and vacuolar thrombolites marks two major events. The first one is the appearance of calcified and mucilaginous cyanobacteria in the Lower-Cambrian Anti-Atlasic platform. The second one is the increase in the water depth from stromatolitic and evaporitic peritidal zone to subtidal thrombolitic environment. In the fossiliferous part of the Lower-Cambrian succession (archaeocyathan series), the flourishing archaeocyatic and calcimicrobe offshore reefal build-ups are evidences of a second episode of sea-level rise. The branching archaeocyathan framework contributes significantly to dendritic calcimicrobe occurrence.
1 Introduction
Dans l'Anti-Atlas (Fig. 1), la succession sédimentaire comprise entre les volcanites et les volcanoclastites du Précambrien III et les schistes à paradoxides du Cambrien moyen, est actuellement rapportée au Cambrien inférieur [7]. Elle se présente, dans le synclinal d'Issafène (Fig. 2), sous forme d'une alternance de faciès carbonatés et silico-clastiques. En plus des archéocyathes, la diversité et la distribution des microbialites identifiées dans les faciès carbonatés, ont permis de subdiviser cette succession en trois séries (Fig. 3A) caractérisées, chacune, par un type particulier de microbialites : (i) la Série à stromatolites, (ii) la Série à thrombolites et (iii) la Série à archéocyathes.
Ce travail, axé sur l'étude des faciès cambriens de la région d'Irherm (Figs. 1 et 2), se fixe comme objectif de discuter le mode de genèse des microbialites, afin de préciser le rôle de ces structures dans la reconstitution et l'évolution des paléoenvironnements. Le découpage lithostratigraphique ici proposé tient compte, non seulement des événements cambriens majeurs, mais également des changements environnementaux intervenus au début du Paléozoı̈que.
2 Série à stromatolites (Fig. 3B)
Cette série correspond au membre carbonaté de la formation Adoudou [7]. Sa limite supérieure, marquée par l'apparition des premières thrombolites, coı̈ncide avec la première anomalie positive des profils isotopiques (δ13C) [13,17,18]. Ces événements sont considérés comme des marqueurs d'une ligne temps isochrone qui souligne le passage Néoprotérozoı̈que–Cambrien inférieur [2,11].
2.1 Description des faciès
Les faciès reconnus dans cette série sont des dolomies à microbialites, qui alternent parfois avec des dolomies massives, des dolomies à texture vacuolaire, des dolomies bréchiques et des strates centimétriques d'argiles rougeâtres.
Les faciès à microbialites se présentent généralement sous la forme de bancs simples de dolomies noires ou beiges, caractérisés par la dominance des stromatolites, localement silicifiés. Les deux variétés morphologiques de lamines stromatolitiques identifiées sont des lamines en dômes (variété LLH [12]) et des lamines planaires (variété Sp). Pour la variété LLH, les lamines sont modelées en une succession latérale de dômes, de rayon généralement centimétrique à pluri-centimétrique. Pour la variété Sp, les lamines sont planaires à légèrement ondulées et se relient latéralement à des dolomies bréchifiées ou vacuolaires. L'aspect microscopique des microbialites montre une alternance de lamines sombres et de lamines claires. Les lamines sombres, dont l'épaisseur ne dépasse pas 1 mm, sont irrégulières et édifiées par une micro-dolosparite brune ou sombre. Les lamines claires, dont l'épaisseur peut atteindre facilement 5 mm, sont édifiées par une dolosparite plus ou moins limpide.
2.2 Interprétation
L'aspect laminé des faciès stromatolitiques est la preuve d'une genèse périodique assurée par l'action combinée de l'activité métabolique des micro-organismes et des processus sédimentaires [15]. Dans la région d'Irherm, les lamines organiques ne sont pas bien préservées dans les édifices stromatolitiques. La structure interne permet, cependant, de supposer que les lamines sombres pourraient dériver de la dégradation de cette fraction organique. Cette dégradation ainsi que la nature exclusivement dolomitique de ces faciès suggèrent leur genèse dans des conditions environnementales similaires à celles reconnues actuellement dans le lagon de Coorong en Australie [4] et le lagon Lagoa Vermelha au Brésil [19]. En effet, la présence d'un substrat organique, enfoui et en voie de décomposition, permet la précipitation d'une dolomicrite, à l'origine de la genèse de lamines carbonatées sédimentaires. Ces lamines micritiques, ultérieurement transformées en dolosparite, seront, à chaque fois, séparées par des lamines organiques qui résultent de l'activité photosynthétique intermittente de la construction microbialitique. La dégradation progressive des lamines organiques, lors de l'enfouissement, s'accompagne d'une nucléation diagénétique d'une dolomite de remplacement pour donner, dans le cas extrême, une micro-dolosparite riche en matière organique, d'où la couleur sombre de ces lamines. Cette dégradation, possible uniquement si la matière organique n'est pas calcifiée, suppose que les micro-organismes générateurs sont des cyanobactéries non calcifiées.
Si l'activité microbialitique s'estompe à l'interface eau–sédiment, alors que la dégradation anaérobie du substrat organique enfoui se poursuit, des niveaux carbonatés, plus ou moins épais, prennent alors naissance par précipitation directe de la micrite dolomitique. Ces faciès, dont l'épaisseur dépend de la durée de l'arrêt de la construction cyanobactérienne en surface, peuvent être massifs et homogènes ou parsemés de cristaux d'anhydrite si le milieu devient confiné. L'augmentation de la taille et la coalescence latérale de minéraux évaporitiques, par accentuation du confinement du milieu, aboutit à l'édification de strates évaporitiques qui alternent avec les niveaux carbonatés. La dissolution ultérieure de ces évaporites aboutit à la genèse des brèches de dissolution et des dolomies vacuolaires.
L'association stromatolites–évaporites montre que le milieu de genèse est moins profond et subit un confinement occasionnel. Il s'agit de la zone intertidale et de sabkhas supratidales. L'aspect laminé de ces microbialites, où interfèrent les lamines organiques et sédimentaires, constitue une preuve d'une croissance latérale et périodique des micro-organismes constructeurs. Compte tenu de l'aspect mou et fragile des cyanobactéries, supposées non calcifiées, ce mode de croissance microbialitique latéral peut être considéré comme une forme d'adaptation à la faible tranche d'eau et aux conditions hydrodynamiques relativement agitées de la zone littorale (Fig. 4).
3 Série à thrombolites (Fig. 3C)
Cette série regroupe la formation Lie-de-Vin et la partie inférieure de la formation d'Igoudine [7]. Elle se caractérise par la prolifération de microbialites à texture grumeleuse ou vacuolaire dites thrombolites [2] et par l'apparition, pour la première fois, des cyanobactéries mucilagineuses calcifiées [11]. Sa limite supérieure, marquée par l'apparition des archéocyathes, coı̈ncide avec la deuxième anomalie positive du profil isotopique (δ13C) [18]. L'apparition des archéocyathes [3] dans l'Anti-Atlas serait liée à une transgression globale qui envahit l'ensemble de l'Altaı̈-Saı̈an (Asie centrale), la Mongolie, l'Espagne, l'Allemagne, le Maroc et l'Avalonia. Cette transgression a permis la migration des archéocyathes à partir de la plate-forme sibérienne où ils sont apparus depuis le Tommotien [3].
3.1 Description des faciès
Les faciès thrombolitiques dominants, en association avec les faciès stromatolitiques et les faciès argilo-gréseux, définissent des séquences sédimentaires de comblement métriques à plurimétriques.
Un faciès élémentaire à thrombolites est constitué par un banc décimétrique à métrique de calcaire dolomitique renfermant des thrombolites. Une thrombolite simple se présente sous forme d'un bioherme décimétrique à métrique et de forme subsphérique à elliptique. Sa texture peut être grumeleuse ou vacuolaire. La variété grumeleuse est formée par un assemblage de grumeaux à contour irrégulier plus ou moins net, le diamètre d'un grumeau ne dépassant pas 4 cm. L'aspect microscopique d'une thrombolite grumeleuse révèle une structure en mosaı̈que de taches irrégulières de micrite sombre et de taches nettement claires de dolosparite rhomboédrique pouvant renfermer des particules clastiques. Pour la variété vacuolaire, les vacuoles correspondent à des cavités de taille pluri-centimétrique et de forme irrégulière, qui peuvent être vides ou partiellement oblitérées par une calcite blanche.
3.2 Interprétation et discussion
Les thrombolites sont interprétées comme des structures organo-sédimentaires édifiées par les cyanobactéries [8,15]. Elles se distinguent des stromatolites par l'absence de la lamination interne, indice d'une croissance périodique [15]. Dans l'Anti-Atlas occidental, leur structure interne témoigne de l'occurrence simultanée d'une phase réellement construite par des communautés microbiennes, représentée par la phase micritique sombre et d'une phase non construite sous forme de carbonates chimiques et de particules silico-clastiques. Les micro-organismes édificateurs correspondent, vraisemblablement, aux cyanobactéries calcifiées et individualisées sous forme de coccoı̈des, eux-mêmes organisés et préservés sous forme de colonies irrégulières. L'apparition des cyanobactéries calcifiées, à la base de la formation Lie-de-Vin de l'Anti-Atlas occidental, a été mise en évidence par Latham et Riding [11]. La partie claire du faciès thrombolitique correspond, en revanche, au remplissage des espaces inter-grumaux par les particules détritiques ou par une calci-sparite dont la dissolution tardive confère une texture vacuolaire à certains édifices thrombolitiques. Pour certains auteurs [1], ces espaces inter-grumaux dérivent plutôt de la destruction de la portion non calcifiée de la matière organique. En effet, selon ces mêmes auteurs, les cyanobactéries, supposées calcifiées et mucilagineuses, auraient la capacité de secréter un mucus organique non calcifié (EPS = extracellular polymeric substance), dont la nature chimique acide empêche sa calcification immédiate. La dissolution ultérieure de ce mucus inter-columnaire par d'autres microbes hétérotrophiques aboutit à la formation des espaces irréguliers dont la taille dépend de la quantité du mucus secrété. Ces espaces peuvent rester vides ou remplis par une calcite blanche, dont la dissolution tardive confère un aspect vacuolaire à certains niveaux thrombolitiques (texture vacuolaire). En cas d'enfouissement rapide de ces constructions, la destruction lente du mucus s'accompagne de l'apparition abondante de la dolomite authigène [22]. Cette dolomite, qui renferme, souvent, des reliques de la matière organique contribue au remplissage de ces espaces de phagocytose (texture grumeleuse).
L'absence de périodicité dans l'édification des thrombolites, comme en témoigne leur structure interne non laminée [15], est une preuve de leur genèse sous une tranche d'eau permanente. Le milieu de formation est, probablement, la zone subtidale (Fig. 4), par comparaison avec les Bahamas où les thrombolites actuelles ont été décrites dans le domaine subtidal [6]. L'apparition quasi constante des stromatolites au sommet des édifices thrombolitiques suggère une croissance des microbialites, d'abord verticalement dans la zone subtidale, puis latéralement dans la zone intertidale. Cette substitution verticale des thrombolites par les stromatolites montre que la charpente microbienne des thrombolites n'est pas assez rigide au point de pouvoir résister aux conditions hydrodynamiques agitées de la zone intertidale.
Pour les thrombolites, la calcification des colonies microbiennes et leur renforcement par le mucus peuvent être considérés comme une autre forme d'adaptation des microbialites aux conditions du milieu, devenu assez profond et moins agité.
4 Série à archéocyathes (Fig. 3D)
Cette série regroupe le membre de Tiout, et les formations d'Amouslek et d'Issafène [7]. Sa limite supérieure est une surface légèrement érosive, surmontée par un niveau brèchique et lumachellique, d'épaisseur variable. Ce niveau est comparable à la « Brèche à micmaca » du Cambrien moyen [9]. Des niveaux lumachelliques, similaires à celui mentionné ci-dessus, ont été décrits au sommet du Cambrien inférieur, dans des régions différentes du globe. Leur intercalation systématique dans des argilites noires, dépourvues de toute trace d'activité organique est un marqueur d'extinction massive des trilobites et d'autres espèces à la fin du Cambrien inférieur et au début du Cambrien moyen [14].
4.1 Description des faciès
Les faciès constitutifs correspondent à la masse édifiée par les calcimicrobes et les archéocyathes sous forme de biohermes et de biostromes. Ces faciès se relient latéralement et se substituent verticalement à des faciès carbonatés détritiques, en grande partie composés de calcimicrobes.
En affleurement, les biostromes se présentent, généralement, sous forme de bancs carbonatés à patine noire ou blanchâtre, épais de 1 à 2 m et d'extension latérale régionale. L'aspect microscopique de ces faciès montre, en plus de quelques archéocyathes solitaires, des particules micritiques uniformément réparties dans un fond microsparitique plus ou moins limpide. Ces particules de taille variant de 0,1 à 0,3 mm ont une forme subsphérique souvent irrégulière et présentent un contour plus ou moins net. Ces masses micritiques, souvent isotropes, se branchent latéralement pour former un réseau ramifié comparable à celui des thrombolites.
Les biohermes apparaissent sous la forme de lentilles de 2 à 30 m d'épaisseur et d'extension latérale régionale. La masse construite des biohermes est un calcaire noir ou blanchâtre, dont la structure interne diffère de celle des biostromes par la fréquence des archéocyathes coloniaux et des particules détritiques silico-clastiques dans la phase inter-columnaire à patine claire.
4.2 Interprétation et discussion
L'abondance, au sein de la masse interne des biostromes, de la phase micritique riche en matière organique suggère que les cyanobactéries calcifiées interviennent activement dans l'édification de la masse récifale. Les particules micritiques constitutives ressemblent aux calcimicrobes réniformes qui caractérisent les groupe Renalcis et Tarthinia [16]. Ces constructions récifales, où les cyanobactéries constituent les principaux organismes constructeurs et où les archéocyathes ne jouent qu'un rôle secondaire, sont fréquents au Cambrien inférieur [10,20]). Elles correspondent aux faciès boundstone selon la terminologie adoptée par Wood [20] et prolifèrent, généralement, dans des eaux profondes et calmes (Fig. 2), comme en témoigne la dominance des cyanobactéries [21].
L'aspect branchu des archéocyathes, synonyme du faciès bafflestone [5], est une caractéristique des organismes bioconstructeurs filtreurs [20]. De telles structures récifales prolifèrent sous des conditions hydrodynamiques relativement agitées [20] et, de préférence, le long des bordures de pentes de plate-forme (Fig. 4). L'abondance des archéocyathes, au sein des biohermes, offre une opportunité au développement des calcimicrobes épibiontes. L'aspect branchu et rigide des archéocyathes détermine, en effet, des espaces irréguliers au sein de la bioconstruction servant de lieux favorables au développement des calcimicrobes épibiontes [20]. Ces calcimicrobes calcifiés, qui se développent sous forme de colonies dendritiques et d'arbustes minuscules attachés aux murailles des archéocyathes, sont facilement arrachés et étalés autour du récif pour former, en association avec les oolithes, des bancs détritiques péri-récifaux. Il semble, par conséquent, que la forme dendritique des calcimicrobes soit étroitement liée à la prolifération accrue des archéocyathes branchus qui jouent un rôle de support rigide et offrent des espaces abrités pour leur développement.
5 Conclusion
La succession des faciès sédimentaires du Cambrien inférieur de la région d'Irherm (Anti-Atlas occidental) est formée de trois séries sédimentaires caractérisées, chacune, par la dominance d'un type particulier de microbialites. La répartition dans le temps de ces structures résulte, probablement, des changements de l'écosystème marin au Cambrien inférieur dans la bordure anti-atlasique du craton ouest-africain.
La Série à stromatolites se caractérise par les seuls faciès microbialitiques à structure laminée. L'aspect mou et fragile des micro-organismes constructeurs, supposés non calcifiés, est l'une des causes principales de leur développement latéral sous forme de microbialites laminées. Il s'agit, par conséquent, d'une forme d'adaptation microbienne aux conditions environnementales à l'aube du Cambrien inférieur. La Série à thrombolites se caractérise par la prolifération des constructions microbiennes à texture grumeleuse. L'apparition des ces structures est favorisée, d'une part, par l'approfondissement du milieu, faisant succéder aux environnements péritidaux des environnements franchement subtidaux et, d'autre part, par la calcification et le renforcement, par le mucus, des colonies calcimicrobiennes. L'épanouissement des biohermes thrombolitiques par croissance verticale des calcimicrobes est une autre forme d'adaptation aux conditions du milieu, devenu assez profond et moins agité.
À l'issue d'un second approfondissement du milieu, qui s'inscrit dans le cadre d'une élévation continue du niveau marin relatif, apparaissent des constructions récifales à archéocyathes et à microbialites. L'aspect rigide et branchu du squelette des archéocyathes est, probablement, l'un des précurseurs de l'acquisition de la forme dendritique par l'une des communautés microbiennes génératrices de récifs de bordure de plate-forme.