

Abridged English version
Since 1978, the Moulay Bousselham lagoon, commonly called Merja Zerga, was declared a biological reserve with an international interest for avifauna. It is also one of the four Moroccan sites that were considered by the RAMSAR Convention for the conservation of humid zones of international interest. This ecosystem shows several ecological originalities. It presents also important social and economical activities due to the practice of traditional fisheries including fish and molluscs. In the beginning of the 1980s, the hydrological, sedimentological and biological state of this lagoon was studied by several authors. These studies showed the perpetual evolution of this lagoon with time, which was apparent by the spectacular changes of its topography, particularly in the outer reaches of the lagoon. In order to explain this situation, a good understanding of the biological and ecological functions of this lagoon is required. With this aim, a typological study was carried out which included monthly measurements of a series of physical-chemical parameters (seawater temperature, salinity, suspended matter, organic and mineral matter and chlorophyll a). Analysis of the Normalised Principal Components combined with Discriminant Analysis of hydrological data was applied to all data collected in all the stations at high and low tide by using the ADE-4 package. The same package was also used to carry out an analysis of variance (Anova-FF). The spatial-temporal analysis of the data approached by Principal Components Analysis (ACP) combined with the Discriminant Analysis (AD) showed the unimportant inter-stations variability, with a percentage of 11% and 9% of the total variance at high tide and low tide, respectively. The results of the inter-stations variability revealed that at high tide, the stations distribute themselves along the F1 axis that translates the functional expression of the salinity. At low tide, the distribution of the stations along the F1 axis reflects a progressive hierarchization from the downstream toward the uphill of the lagoon, also dictated by the salinity, which shows a meaningful spatial variation (). This configuration makes it possible to subdivide the site in three different zones, although this zonation does not appear strong enough, which justifies the weak value of the percentage of the inter-station variability (9%). These results underline that oceanic waters constitute the main motor of circulation of water inside the lagoon, with a renewal of about 98% at every tide cycle. In opposition to their spatial variation, the different measured parameters present a meaningful monthly variation (); nevertheless, the results of the analysis of the inter-months variability show that only the temperature and the salinity have a clean seasonal rhythm.
1 Introduction
Depuis 1978, la lagune de Moulay Bousselham, appelée communément Merja Zerga, est déclarée réserve biologique d'intérêt mondial pour l'avifaune. Elle est aussi classée parmi les quatre sites marocains retenus par la convention de RAMSAR pour la conservation des zones humides d'importance internationale. Cet écosystème présente plusieurs originalités écologiques [3] et connaît d'importantes activités socio-économiques, en raison de la pratique de la pêche artisanale des poissons et des mollusques [2]. Depuis les années 1930, la lagune a attiré l'attention des chercheurs ; toutefois, les études qui y sont effectuées sont restées pour longtemps peu nombreuses. Ce n'est qu'au début des années 1980 que ce site a fait l'objet de plusieurs travaux de recherche [3,5,8,14,17] visant à définir son organisation hydrologique, sédimentologique et biologique. Ces études ont mis en avant l'évolution perpétuelle de la lagune au fil du temps, qui se traduit par l'obstruction progressive du goulet qui assure l'échange entre la lagune et l'océan Atlantique. Cependant, le découpage de la lagune en différentes zones s'avère nécessaire pour une bonne compréhension du fonctionnement hydrologique. À cet effet, une étude typologique de la lagune a été entreprise sur la base des mesures mensuelles des paramètres physico-chimiques durant un cycle annuel.
2 Matériel et méthodes
2.1 Site étudié
Étendue sur une surface de 35 km2, la lagune de Moulay Bousselham est située à 125 km au nord de Rabat, entre 34°47′ et 34°52′ de latitude nord et entre 6°13′ et 6°18′ de longitude ouest (Fig. 1). L'alimentation et l'évacuation de ses eaux sont assurées par un réseau de chenaux, qui garantit l'échange entre la lagune et l'océan Atlantique, et ce, par le biais d'un goulet étroit d'une longueur de 50 m quand il est opérationnel. Par ailleurs, cet écosystème est alimenté par un ensemble d'apports en eaux douces : les débouchés de l'oued Drader, au nord-est, et du canal de Nador, au sud, y demeurent d'un apport important (Fig. 1). La profondeur de la lagune varie entre 0 et 2 m (en moyenne 50 cm) suivant le cycle tidal et les précipitations [14].
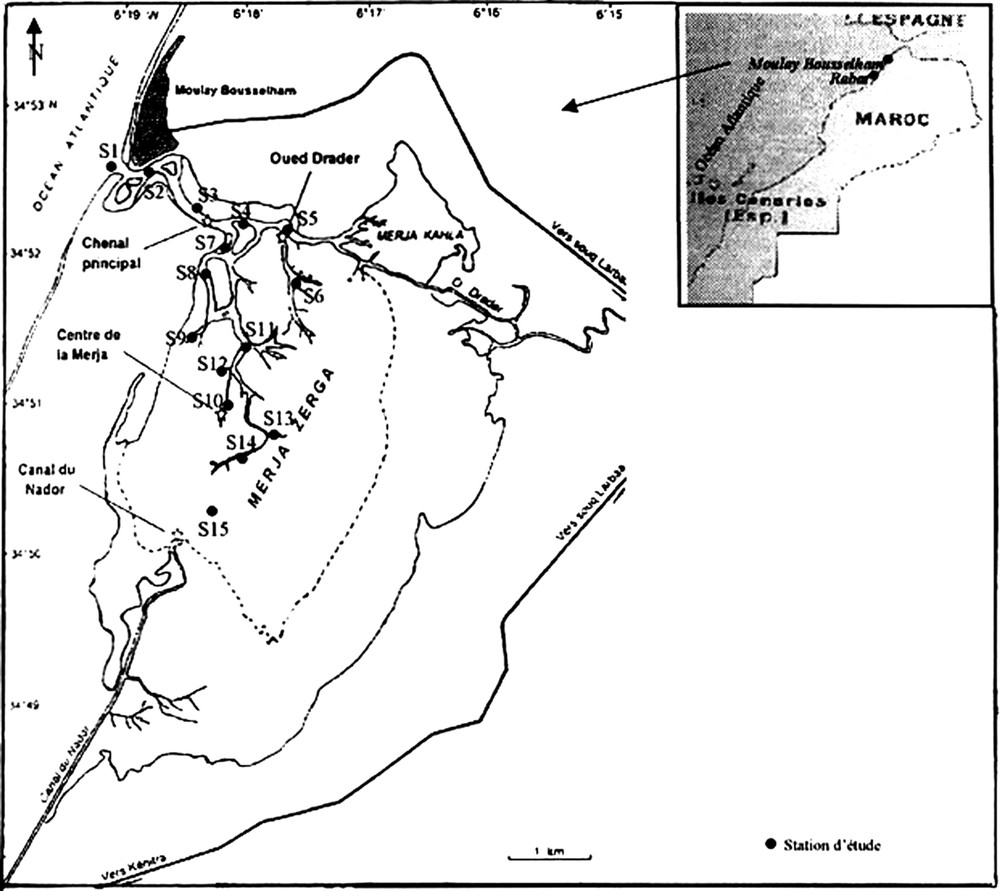
Localisation des stations d'échantillonnage d'eau pour l'étude hydrologique réalisée entre juillet 2001 et juin 2002.
Sampling network of the hydrological parameters during July 2001 and June 2002.
2.2 Prélèvement et méthodes d'analyses
L'étude de la typologie de la lagune a été basée sur l'analyse en composantes principales (ACP) combinée à l'analyse discriminante (AD). Six paramètres hydrologiques ont été choisis pour caractériser la lagune : la température de l'eau , la salinité mesurées in situ, les matières en suspension (MES), les matières minérales particulaires (MMP), la matière organique particulaire (MOP) et la chlorophylle a (CHLO), déterminées après traitement des échantillons d'eau.
Les prélèvements d'eau ont été effectués en surface à marée haute et à marée basse des mortes eaux durant un cycle annuel (juillet 2001–juin 2002) au niveau de 15 stations, réparties et numérotées par ordre croissant de l'aval vers l'amont de la lagune (Fig. 1).
Le traitement des données par ACP et par AD ainsi que la représentation graphique des résultats ont été réalisés avec le logiciel ADE-4. L'inertie totale de chacune des deux matrices des données – à marée haute et à marée basse – est décomposée en une part intra-stations et intra-mois et en une autre part inter-stations et inter-mois. Cette décomposition de l'inertie est faite en utilisant la métrique euclidienne [11]. Une analyse de variance (Anova1-FF) a été appliquée également en utilisant le même logiciel pour tester la signification des différences des paramètres hydrologiques entre les stations d'une part, et entre les mois d'autre part [18].
3 Résultats
3.1 Variation spatio-temporelle des paramètres hydrologiques
Afin d'étudier la variation spatio-temporelle des paramètres hydrologiques, nous avons analysé, dans un premier temps, l'évolution de la valeur moyenne de chaque paramètre, calculée sur toute la lagune en fonction du temps. Dans un second temps, nous avons étudié la variation de la valeur moyenne annuelle de ces mêmes paramètres en fonction des stations. Les résultats sont présentés sur la Fig. 2.





Variations mensuelle et spatiale de la salinité (a), de la température (b), de la chlorophylle a (c), de la matière en suspension et minérale particulaire (d) et de la matière organique particulaire (e) au cours des deux cycles de marées.
Monthly and spatial variation of salinity (a), temperature (b), chlorophyll a (c), mineral particular matter and suspended matter (d) and particular organic matter (e) during two tidal cycles.
3.1.1 La salinité
Les taux de salinité relevés à marée haute et en aval de la lagune sont toujours supérieurs à ceux mesurés à marée basse et en amont (Fig. 2a). En effet, de la station S15 à la S1, la salinité varie entre 28 et 36‰ à marée haute et entre 9 et 31‰ à marée basse. Au cours du cycle annuel, la salinité moyenne des eaux de la lagune fluctue de 24‰ à 36,28‰ à marée haute et de 8‰ à 32,52‰ à marée basse (Fig. 2a).
3.1.2 La température
La variation de la température des eaux de la lagune suit étroitement la variation de la température atmosphérique. En effet, elle varie entre 15,72 et 22,33 °C à marée haute et entre 12,95 et 30,78 °C à marée basse (Fig. 2b). La température sur l'ensemble du bassin est relativement homogène, tant à marée haute qu'à marée basse : elle fluctue respectivement entre 17,6 et 18,7 °C et entre 20 et 23 °C (Fig. 2b).
3.1.3 La chlorophylle a
Mis à part le mois de janvier, où une concentration chlorophyllienne maximale (0,76 mg m−3) a été relevée à marée basse, la variation mensuelle montre que la biomasse en chlorophylle a est relativement élevée à marée basse (0,10–0,32 mg m−3), par comparaison avec celle déterminée à marée haute (0,07–0,20 mg m−3) (Fig. 2c). À l'échelle spatiale, à marée haute, la concentration chlorophyllienne varie faiblement d'une station à une autre, entre 0,10 et 0,17 mg m−3. À marée basse, elle évolue entre 0,15 et 0,47 mg m−3, sans décrire un gradient aval–amont (Fig. 2c). Le pic enregistré en janvier est dû aux déversements des rizières dans la lagune par le biais du canal de Nador.
3.1.4 La charge particulaire
La concentration des matières en suspension, essentiellement les particules minérales, oscille autour d'une valeur de 15 mg l−1 à marée haute et de 35 mg l−1 à marée basse (Fig. 2d). Au mois de novembre, nous avons enregistré une forte concentration en matières minérales, à la fois à marée haute (106 mg l−1) et à marée basse (252 mg l−1). Cette forte charge particulaire est liée aux coups de vents forts qu'a connus cet écosystème durant ce mois. En effet, étant donné la faible épaisseur de la tranche d'eau, particulièrement à marée basse, et la nature meuble du substrat, le vent a tendance à remettre en suspension les particules, tout en favorisant le brassage de la masse d'eau.
Les teneurs en matière organique varient entre 3 et 9 mg l−1 durant la période échantillonnée pour les deux cycles de marées, et pour toutes stations confondues (Fig. 2e), excepté en novembre à marée basse, où une teneur de l'ordre 24 mg l−1 a été enregistrée.
3.2 Typologie spatio-temporelle, rythme saisonnier du milieu
Les résultats de l'analyse discriminante mettent en évidence la faible variabilité inter-station du site étudié, aussi bien à marée haute qu'à marée basse (Tableau 1). Pour ce qui suit, on représentera les résultats de la variabilité inter-station (Fig. 3a et b), d'une part, et ceux de la variabilité inter-mois (Fig. 4a et b), d'autre part. Une fois calculées les composantes principales de l'ACP, les deux premiers axes ont été retenus pour la représentation graphique des résultats pour chaque type de variabilité. Ces deux axes expliquent plus de 83 % de l'information contenue dans la matrice de données (Tableau 2).
Pourcentages des différents types de variabilité obtenus par l'analyse discriminante
Percentage of different kinds of variability obtained by discriminated analysis
| Type de variabilité | Pourcentage de variabilité | |
| Marée haute | Marée Basse | |
| Intra-stations / Inter-stations | 89/11 | 91/9 |
| Intra-mois / Inter-mois | 70/30 | 35/65 |
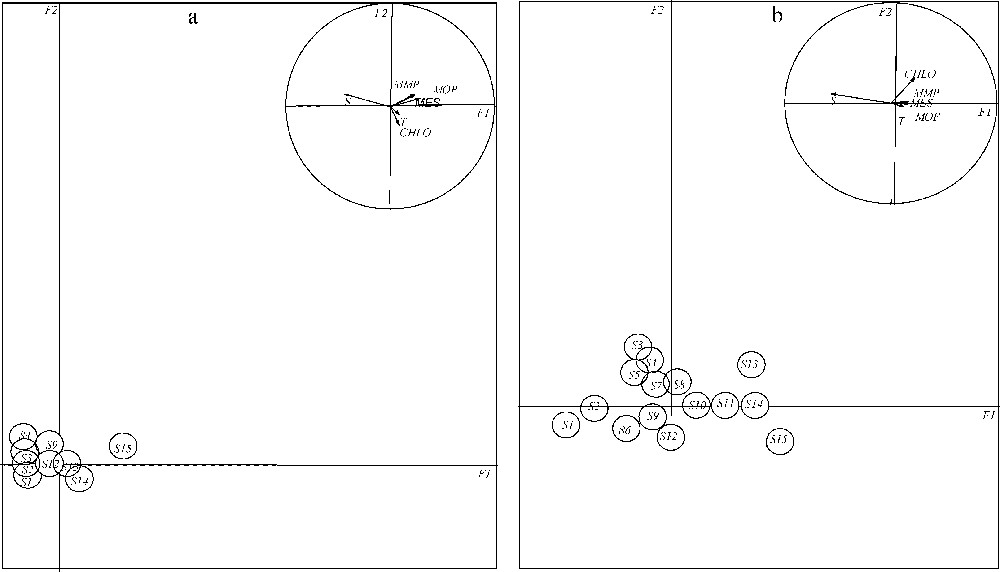
Résultats de la variabilité inter-stations à marée haute (a) et à marée basse (b) dans le plan factoriel F1 × F2.
Results of the inter-stations variability during high (a) and low tide (b) in the factorial plan F1 × F2.

Résultats de la variabilité inter-mois à marée haute (a) et à marée basse (b) dans le plan factoriel F1 × F2.
Results of the inter-months variability during high tide (a) and low tide (b) in the factorial plane F1 × F2.
Pourcentages d'inerties et d'inerties cumulées des deux premiers axes factoriels
Inertia and cumulated inertia percentage of the two first factorial axis
| Type de variabilité | Pourcentage d'inerties (%) | Pourcentages d'inerties cumulées des deux premiers axes (%) | |
| F1 | F2 | ||
| Inter-station | |||
| MH | 73,57 | 15,94 | 89 |
| MB | 76,79 | 12,22 | 89 |
| Inter-mois | |||
| MH | 55,05 | 27,73 | 83 |
| MB | 63,33 | 20,84 | 84 |
3.2.1 Variabilité inter-stations
À marée haute, les stations se distribuent le long de l'axe F1 (Fig. 3a), qui traduit la salinité. La station S15 s'individualise nettement des autres stations, qui dessinent à leur tour un gradient aval–amont en direction du canal de Nador (S12, S13 et S14).
À marée basse, la disposition des stations selon l'axe F1 reflète une hiérarchisation progressive de l'aval vers l'amont de la lagune, dictée également par la salinité. Cette configuration permet de subdiviser le site en trois zones différentes, bien que cette zonation ne soit pas très marquée (Fig. 3b), ce qui justifie la faible valeur du pourcentage de la variabilité inter-station (10 %) (Tableau 1). On distingue une zone située en aval comprenant les deux stations (S1 et S2), soumise à une nette influence marine, une zone intermédiaire ou zone de transition, regroupant neuf stations, qui s'étale du chenal primaire jusqu'aux extrémités du chenal tertiaire et secondaire (de S3 au S10, S12), et une zone amont, comprenant quatre stations (S11, S13, S14 et S15), alimentée par les eaux douces du canal de Nador. Les autres paramètres mesurés température, chlorophylle a, matière en suspension, matière minérale particulaire et matière organique particulaire ne contribuent pas à l'explication de la variabilité inter-station, aussi bien à marée haute qu'à marée basse.
3.2.2 Variabilité inter-mois
À marée haute, dans le plan factoriel F1 × F2, la répartition des mois se fait suivant l'axe F1, qui exprime, de la droite vers la gauche, l'opposition des mois chauds à basses salinités aux mois froids à fortes salinités (Fig. 4a). À marée basse, bien que leur dispersion soit plus importante, les mois s'ordonnent de la même manière qu'à marée haute, excepté le mois de novembre, qui se détache, les autres mois se répartissant d'une façon longitudinale le long de l'axe F2. Suivant cet axe, on peut subdiviser ce groupe en trois sous-ensembles. Dans ces valeurs positives, un premier sous-ensemble représente la saison d'été (juillet, août, septembre), durant laquelle les eaux de la lagune sont chaudes et moins salées, un sous-ensemble intermédiaire à la saison du printemps rassemble les mois d'avril, mai, juin, et un troisième comprend les mois de décembre, janvier et février, correspondant à la saison d'hiver, pendant laquelle les eaux sont froides et salines (Fig. 4b).
4 Discussion
L'étude typologique entreprise durant un cycle annuel dans la lagune de Moulay Bousselham a permis de définir les grands traits de son fonctionnement hydrologique. Les résultats de l'ACP et de l'AD ont montré que le pourcentage de la variabilité inter-station est faible, de l'ordre de 10 % pour les deux cycles de marées (Tableau 1). Ce pourcentage de variabilité est attribué seulement à la salinité qui montre une variation spatiale significative (à ) (Tableau 3).
Résultats de l'analyse de variance des paramètres hydrologiques
Variance analysis results of hydrological parameters
| Paramètres | Stations | Mois | ||||||
| Marée haute | Marée basse | Marée haute | Marée basse | |||||
| F | P | F | P | F | P | F | P | |
| Température | 0,22 | ns | 0,38 | ns | 68,81 | *** | 190,7 | *** |
| Salinité | 3,30 | *** | 5,97 | *** | 13,71 | *** | 19,52 | *** |
| Chlorophylle a | 0,83 | ns | 1,52 | ns | 3,18 | *** | 8,98 | *** |
| Matière en suspension | 0,90 | ns | 0,49 | ns | 1,83 | ns | 32,18 | *** |
| Matière minérale particulaire | 0,93 | ns | 0,48 | ns | 1,74 | ns | 33,05 | *** |
| Matière organique particulaire | 1,23 | ns | 0,54 | ns | 1,74 | ns | 27,75 | *** |
De telles structures hydrologiques spatiales déterminées à chaque cycle de marée montrent que malgré l'étroitesse du goulet, les eaux océaniques constituent le moteur principal de circulation des eaux intra-lagunaires. Le volume des eaux océaniques oscillant dans la lagune est estimé à , soit un renouvellement d'environ 98 % seulement contre 2 % d'apports d'eaux continentales, véhiculés par les oueds afférents, essentiellement ceux provenant du canal de Nador () [8]. Quoique les apports continentaux soient négligeables par rapport aux apports marins, leur effet est plus ressenti à marée basse, tout en décrivant un gradient aval–amont. À ce gradient décroissant hyalin se manifestent des gradients écologiques des différents peuplements qui colonisent cet écosystème [3,4], tant dans la composition qualitative des peuplements que dans les paramètres de structure (richesse spécifique, densité, biomasse), depuis la communication avec la mer jusqu'aux zones les plus internes de la lagune. Bazairi et al. [3] ont déterminé trois peuplements macrozoobenthiques en allant de l'aval vers l'amont de l'écosystème. Le premier est un peuplement à Cerastoderma edule, qui se présente sous trois faciès, le faciès à Nephtys cirrosa (groupe SS1), le faciès à Nassarius reticulatus (noyau SS2) et le faciès à Scrobicularia plana (noyau SS3). Le deuxième est un peuplement à Scrobicularia plana (groupe SS4). Le troisième est un peuplement à Tapes decussatus–Scrobicularia plana–Melita palmata (groupe SS5) (Fig. 5). De même, l'indice de diversité de Shannon–Weaver varie selon un gradient aval–amont entre 1,83 bits (groupe SS4) et 2,56 bits (groupe SS1) [3].

Représentation graphique des nuages des points stations et espèces dans le plan factoriel F1 × F2 de l'AFC appliquée en zone subtidale. Ced : Cerastoderma edule, Nci : Nephtys cirrosa, Nre : Nassarius reticulatus, Spl : Scrobicularia plana, Tde : Tapes decussatus et Mpa : Melita palmata.
Subtidal stations and significant species on the first two axes of correspondence analysis. Ced: Cerastoderma edule, Nci: Nephtys cirrosa, Nre: Nassarius reticulatus, Spl: Scrobicularia plana, Tde: Tapes decussatus, and Mpa: Melita palmata.
Les valeurs de salinité relevées dans l'ensemble de l'écosystème sont voisines de celles trouvées par Bazairi et al. [3], Benbakhta [4] et Benhoussa [5]. Cependant, en période de fermeture de la passe en 1991, la salinité a été plus basse (6‰) [17]. Dans les lagunes atlantiques sud-marocaines (Oualidia et Sidi Moussa), les salinités à marée basse sont de l'ordre de 25 à 32‰ [6,10]. En Méditerranée, dans la lagune de Nador, les eaux ont tendance à une sursalure (de 34 à 43,5‰), liée principalement à une évaporation importante dans certaines zones peu profondes [1,13]. Par ailleurs, au cours des deux cycles de marées, l'ACP et l'AD ont montré que les autres paramètres hydrologiques (T, CHLO, MES, MMP et MOP) ne contribuaint pas, dans la lagune de Moulay Bousselham, à la variabilité inter-station. Ce résultat met en évidence la dynamique des eaux, qui a tendance à jouer un rôle de brassage des différentes masses d'eaux, empêchant tout établissement d'une stratification verticale et diminuant le gradient aval–amont. Dans l'ensemble du site, la biomasse chlorophyllienne est relativement faible vis-à-vis des valeurs habituellement recensées dans les bassins paraliques périméditerranéens [12,15] ; elle varie entre 0,10 et 0,17 mg m−3 à marée haute et entre 0,15 et 0,47 mg m−3 à marée basse, ce qui traduit le non-confinement global du bassin [13]. Ces valeurs sont très faibles par rapport à celles déterminées dans le même écosystème par Rharbi [19] entre 0,2 à 9,2 mg m−3 et par Benhoussa [5] (entre 0,02 et 9,63 mg m−3). Rharbi a expliqué cette richesse chlorophyllienne par l'élévation anormale de la température qu'a connue la lagune à la fin de 1989 et durant le printemps 1990.
Comparée à d'autres sites marocains, la biomasse en chlorophylle a dans la lagune de Moulay Bousselham est relativement faible par rapport à celle relevée dans la lagune de Oualidia, où elle fluctue entre 0,7 et 1,18 mg m−3 [10]. En Méditerranée, dans la lagune de Nador, elle varie entre 0,3 mg m−3 et 2,8 mg m−3 [1,13]. Par ailleurs, la concentration en chlorophylle a dans la lagune de Moulay Bousselham est voisine de celle mesurée dans le lagon sud-ouest de Nouvelle-Calédonie (0,2 et 0,4 mg m−3) [7] et dans le lagon de Takapoto – Tuamotu, Polynésie française – (inférieures à 0,3 mg m−3) [16]. Cependant, en rade de Brest (France), la biomasse chlorophyllienne fluctue largement entre 0,66 et 8,85 mg m−3 [9].
Contrairement à leur variation spatiale, les paramètres mesurés présentent une variation mensuelle significative (à ) (Tableau 3). Cependant, les résultats de l'analyse de la variabilité inter-mois montrent que seules la température et la salinité possèdent un rythme saisonnier net (Figs. 4a et b). Les valeurs de la salinité sont minimales en été et sont maximales en hiver, contrairement à celles qui ont été soulignées dans les travaux antérieurs [3,4,8,19]. Cette situation inversée dépend étroitement du régime hydrologique du site, déterminé, quant à lui, par l'amplitude des marées, ainsi que par les conditions climatiques, notamment l'importance des arrivées d'eaux douces du continent, dépendant de l'abondance des pluies.
5 Conclusion
Le renouvellement de la lagune de Moulay Bousselham en eau d'origine marine est considérable et constitue le moteur principal de la circulation des eaux intra-lagunaires. De même, cette étude a relevé la complexité du fonctionnement de cet écosystème, qui réside dans l'instabilité temporelle des paramètres hydrologiques régis par les facteurs climatiques et dans l'alimentation permanente par les eaux continentales. Ainsi, cette étude permettra d'adopter une meilleure approche méthodologique visant à réduire le nombre de stations et d'augmenter la fréquence d'échantillonnage, en vue d'une compréhension approfondie du fonctionnement hydrologique du site pour garantir sa protection.