

Abridged English version
The proteus is an amphibian with four members that have three fingers at the front and two toes at the rear. They are only used for walking and crawling on a humid ground, such as chitons and salamanders. P. Schlegel argued that the distribution of orientation of the proteus within the Earth's magnetic field shows some preference for the north–south direction. In order to test the hypothesis of magnetotactism, we carried out magnetic measurements on four animals from the underground Moulis laboratory (Ariège, France). No stable signal could be detected when measuring the Natural Remanent Magnetization (NRM) of the proteus. The proteus was thus given an anhysteretic remanence, which was also too weak for being detected. We thus produce a strong isothermal remanent magnetization. The front part of the animal, including its head, front legs and its upper body took a magnetic moment in the order of 10−10 A m2, whereas the rear part of the body is almost 100 times stronger. The stepwise acquisition curves of isothermal remanent magnetization (IRM) confirmed that the magnetization of the rear part of the proteus saturates after 0.2 T, and is thus associated with relatively soft coercivities that could be typical of magnetite. The presence of magnetite can be suspected also from the AF demagnetization curves of the IRM. In all cases, about 75% of the initial IRM was lost after 20 mT. Thus, at this stage, we had indications that remanent magnetism was present within the rear body and the rear legs of the animal, while no significant remanence was detected in the head. Low-temperature measurements were then performed from 1.7 up to 300 K using the Quantum design MPMS superconducting susceptometer of the Solid-State Physics laboratory of the Paris-6 University. In most cases, the signal was lower than the persistent paramagnetism of the gelatine capsule that maintained the sample within a straw used as a sample holder. After removing the effect of the capsule, the magnetization remains constant, with negative values. In contrast, the intestine and the rectal part show a much stronger signal and a different pattern. The magnetization remained positive and decreased between 1.7 and 60 K as a result of unblocking superparamagnetic grains. The most striking observation was the existence of a significant drop in the saturation magnetization () of the rectal part at about 120 K. This change is typical of the Verwey transition, in which the magnetite electron-spin lattice undergoes reorganization. Because these magnetite crystals were found in the intestine, they could have been ingested by the animals. The measurements performed on soil samples indeed provided parameters in good agreement with the parameters related to the saturation magnetization of the animal, with a similar viscosity. Noticeably also, the comparison between the demagnetization curves of the rectal parts of the animals and those associated with a variety of soil grains indicates that they likely have the same provenance.
All these experiments were performed with the most sensitive cryogenic magnetometers available. However they are not capable of detecting a few monodomain grains of magnetite. In this respect, none of the experiments are fully conclusive. Hopefully, we should be able to perform more sensitive measurements in a near future using a new generation of gradiometers.
1 Introduction
Il est maintenant établi que plusieurs vertébrés sont sensibles aux changements en direction et/ou en intensité du champ magnétique ambiant. La magnétosensibilité peut être due à des courants induits dans des électrorécepteurs par les mouvements de l'animal (ou les mouvements des masses d'eau) dans le champ magnétique ambiant, comme cela a été suggéré pour des poissons d'eau de mer [11]. Un deuxième mécanisme suggère un lien entre la magnétoréception et le système visuel [9,12–14]. Enfin, une hypothèse probable serait que des cristaux de magnétite biogénique soient utilisés comme de petites boussoles pour transmettre des informations sur le champ géomagnétique via le système nerveux.
Le protée fut initialement découvert dans une source en Slovénie et signalé pour la première fois en 1689. Néanmoins, il n'a été réellement décrit que 80 années plus tard, par le physicien Joseph Nicolai Laurenti, sous le nom de Proteus anguinus. C'est un amphibien à quatre membres (équipés respectivement de trois et deux doigts à l'avant et à l'arrière), qui, comme pour les salamandres, lui servent à nager dans l'eau et à marcher sur les sols humides. Il peut être considéré comme un véritable fossile vivant, puisque ses ancêtres vivaient dans les marais du Crétacé. L'habitat du protée est confiné à des grottes, car il ne peut pas survivre en surface – il est notamment trouvé en abondance dans les eaux souterraines des grandes cavités creusées dans les roches karstiques de Slovénie, Croatie et Dalmatie. Sa taille varie de 20 à 35 cm, mais sa croissance se poursuit tout au long de sa vie. La couleur blanc–rose de l'animal résulte de la dépigmentation de la peau et de sa coloration par les globules rouges. Il est aveugle, en dépit d'un essai de développement de la vue, qui a vite avorté. Le protée ne pèse qu'une dizaine de grammes et peut survivre sans nourriture pendant plus de douze semaines.
Peter Schlegel [19] a effectué plusieurs expériences au laboratoire souterrain du CNRS à Moulis (France), dans lesquelles il a utilisé des stimuli électriques variés pour étudier les mécanismes d'orientation de l'animal. Il a observé une sensibilité aux courants alternatifs jusqu'à des basses fréquences de 0,1 Hz, plus particulièrement développée entre 25 et 40 Hz. L'animal effectue alors des mouvements nerveux de la tête et s'éloigne fréquemment de la source électrique. D'autres expériences consistant à approcher rapidement un aimant permanent de forte intensité magnétique ont, comme dans le cas précédent, engendré des mouvements nerveux et des réactions de fuite. Des tests supplémentaires ont montré que les protées étaient insensibles aux champs magnétiques induits par des courants continus. Il en conclut que le champ magnétique terrestre ne serait pas assez puissant pour exciter les récepteurs du protée.
Dernièrement, P. Schlegel a étudié les orientations du protée dans le champ magnétique terrestre, en enregistrant la distribution des orientations de six animaux placés pendant dix heures dans le champ magnétique terrestre, puis durant dix autres heures dans un champ pour lequel on a inversé la composante horizontale. Les distributions obtenues montrent une préférence directionnelle, mais aucun changement systématique d'orientation n'a été détecté après avoir inversé le champ.
La sensibilité électrique de l'animal étant écartée, de même que la magnétoréception chimique, puisque le protée est aveugle, sa sensibilité au champ magnétique, si elle existe, fonctionne donc avec un autre type de capteur.
L'influence du champ magnétique ayant été décelée dans deux groupes de salamandres cavernicoles (Eurycea lucifuga et Notophthalmus viridescens), la magnétoréception serait le mécanisme le plus plausible [1,3,8,16]. Pour le prouver, il faudrait pouvoir détecter la présence d'oxydes magnétiques à l'intérieur de l'animal. Une observation directe par observation en microscopie à transmission est impossible, si elle n'est pas d'abord guidée par des mesures magnétiques. Le but de cette étude consiste donc à détecter la présence d'oxydes magnétiques dans le protée par des mesures magnétiques à haute sensibilité, afin de mieux caractériser les mécanismes qui seraient associés au guidage et à l'orientation de l'animal.
2 Mesures
2.1 Généralités
Pour toutes les expériences, nous avons bénéficié de l'existence du laboratoire CNRS souterrain de Moulis (Ariège, France), situé au pied des Pyrénées françaises, à 110 km au sud de Toulouse, dans lequel des centaines de protées, certains d'entre eux âgés de plus de 20 ans, sont élevés dans de grands réservoirs d'eau.
Les mesures magnétiques ont été effectuées sur quatre animaux fournis par J.-P. Durand, ou prélevés directement sur place à Moulis. Les animaux ont été immergés dans le glutaraldéhyde ou dans l'alcool, afin de les préserver dans les meilleures conditions. La dissection a eu lieu dans un environnement propre, en utilisant des matériaux non magnétiques, notamment des couteaux en céramiques. Aucune contamination ne peut donc raisonnablement être suspectée lors des manipulations. Les premières mesures magnétiques ont été conduites dans la pièce amagnétique de l'Institut de physique du Globe de Paris (IPGP), avec un dispositif supraconducteur à interférences quantique (SQUID). La limite de détection de ce magnétomètre est de 10−11 A m2. Un porte-échantillon spécial a été conçu pour réduire la quantité de matière nécessaire et le niveau de bruit de fond de l'appareil. L'absence de signal stable ne signifie pas l'absence de grains magnétiques dans le protée, car elle peut résulter du manque de sensibilité de l'appareil de mesure. Un unique grain de magnétite monodomaine porte une aimantation de 10−14 A m2, de plusieurs ordres de grandeurs plus faible que notre limite de détection. Rappelons toutefois que de la magnétite monodomaine a été détectée en utilisant ce type d'appareillage chez les abeilles [4], les pigeons [6,7,17,18], les saumons [10], les fourmis [15], le papillon monarque [2] et les anguilles [5].
La mesure de l'aimantation naturelle rémanente (NRM) par nos magnétomètres cryogéniques, n'a donné aucun résultat. Nous avons ensuite soumis les animaux à différents champs magnétiques susceptibles d'induire une aimantation rémanente en remobilisant les grains magnétiques. En premier lieu, nous avons appliqué un champ continu de faible intensité (45 mT), en présence d'un champ alternatif décroissant, d'une amplitude maximale de 80 mT, pour produire une aimantation rémanente anhystérétique (ARA). Là encore, le magnétomètre n'a pu détecter aucun signal. Nous avons alors appliqué un champ intense par paliers progressifs jusqu'à 1,2 T, en utilisant un électroaimant, pour produire une aimantation rémanente isotherme (ARI). La Fig. 1 montre les moments magnétiques mesurés après les premier (0,2 T) et dernier (1,2 T) paliers, pour des parties différentes d'un protée long de 15 cm. La partie du protée incluant la tête, les pattes avant et la partie supérieure du corps a un moment magnétique moyen de l'ordre de 10−10 A m2, tandis que la partie arrière est presque 100 fois plus forte. La stabilité du signal a été évaluée par un test de viscosité. L'indice de viscosité, défini comme le pourcentage d'aimantation perdu après 14 jours en champ nul, indique (Fig. 1) que la partie avant de l'animal présente un taux d'aimantation visqueuse à court terme significatif, tandis que la partie arrière est considérablement moins affectée. Ces deux parties ont donc des caractéristiques d'aimantation différentes. Une forte valeur de la viscosité est souvent liée à des grains multidomaines, qui ne portent pas d'aimantation rémanente stable. L'application d'un champ intense écarte les parois des domaines magnétiques. Leurs aimantations individuelles ne se compensent plus mutuellement et finissent par créer une aimantation résultante. Néanmoins, cette composante décroît rapidement après suppression du champ, en raison de la relaxation des moments magnétiques vers leur état initial.
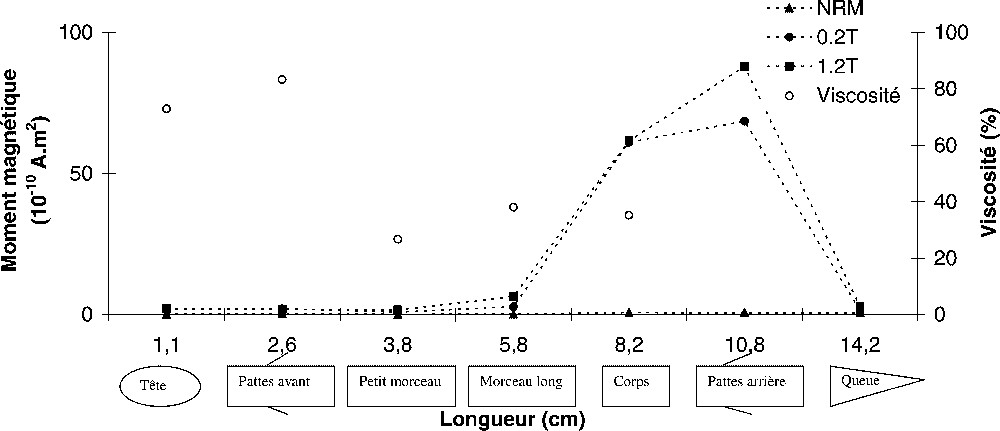
Moments magnétiques (symboles fermés) mesurés après avoir aimanté l'animal dans un champ de 0,2 T, puis de 1,2 T. Excepté en ce qui concerne le corps et les pattes arrière de l'animal, aucune aimantation n'est détectée. L'index de viscosité (symboles ouverts), plus élevé pour la partie avant de l'animal, indique l'absence de stabilité magnétique, et donc de rémanence.
Magnetic moments (closed symbols) measured after magnetizing the animal in a 0.2-T and a 1.2-T field. Except for the body and the rear legs of the animal, no magnetization can be detected. The viscosity index (open symbols) is higher in the front part of the animal, which indicates the absence of magnetic stability and thus of remanence.
Les courbes d'acquisition d'IRM par paliers (Fig. 2a) ont confirmé que l'aimantation de la partie arrière du protée sature après 0,2 T, et est donc associée à des coercivités relativement faibles, qui peuvent être typiques de la magnétite. Les échantillons ont ensuite été désaimantés par champs magnétiques alternatifs décroissants (af). Dans tous les cas (Fig. 2b), à peu près 75 % de l'IRM initiale a été perdue après application d'un champ crête à crête de 20 mT, observation compatible avec la présence de magnétite.

(a) Acquisition par paliers d'aimantation rémanente isotherme (ARI ou IRM) jusqu'à un champ maximum de 1,2 T. Les résultats confirment la présence d'une aimantation stable dans le corps et les pattes arrière de l'animal. (b) Désaimantation par champ alternatif par paliers des échantillons saturés. Les coercivités sont homogènes et typiques de la magnétite. La forme irrégulière des courbes, caractérisées par un moment magnétique très faible, indique la limite des mesures et l'absence de composante.
(a) Stepwise acquisition of Isothermal Remanent Magnetization (IRM) up to a maximum field of 1.2 T. The results confirm the presence of stable magnetization in the body and the rear legs of the animal. (b) Stepwise alternating field demagnetization of the saturated samples. The coercivities of the samples are similar and typical of magnetite. The irregular shape of the curves, which are characterized by a very low magnetic moment, indicates the limit of the measurements and the absence of a reliable remanence component.
À ce stade, nous avons donc des indications sur la présence d'une aimantation rémanente stable dans la partie arrière du corps et les pattes arrière de l'animal, tandis que des composantes très instables ont été détectées dans la tête. Des expériences supplémentaires sont nécessaires pour tenter d'établir la nature des grains magnétiques et leur origine.
2.2 Mesures à basses températures
L'évolution du moment magnétique lors de cycles à basses températures permet d'observer des transitions caractéristiques de la minéralogie magnétique. Nous avons utilisé le susceptomètre supraconducteur MPMS (Quantum Design) du Groupe de physique des solides de l'université Paris-6. Cet instrument, équipé de capteurs à squids, fonctionne à l'hélium liquide, à la fois pour la mesure et le refroidissement des échantillons en présence d'un champ appliqué pouvant aller jusqu'à 5,5 T. Sa sensibilité est de l'ordre de . Les mesures sont effectuées pendant le réchauffement entre 1,7 et 300 K. Un protée long de 20 cm, n'ayant subi ni ARA ni ARI, a été disséqué en 22 échantillons, qui ont été déshydratés afin d'éliminer le signal diamagnétique de l'eau. La distribution des morceaux et leurs caractéristiques magnétiques (diamagnétisme, ferrimagnétisme...) sont récapitulées dans le Tableau 1. Les variations de des différentes parties du protée en fonction de la température sont représentées sur la Fig. 3. Dans la plupart des cas (crâne, mâchoire, encéphale, peau, muscle, yeux, os, pattes avant), le signal est plus faible que le signal paramagnétique de la capsule en gélatine qui maintient l'échantillon (Fig. 3a). Après avoir éliminé l'effet de la capsule, l'aimantation reste constante, avec des valeurs négatives (Fig. 3b), probablement en raison du diamagnétisme de l'eau restant dans l'animal. En revanche, les intestins et la partie rectale montrent un signal plus fort et un comportement différent (Fig. 4). L'aimantation reste positive et décroît entre 1,7 et 60 K, en raison du déblocage de grains superparamagnétiques. L'observation la plus frappante est la chute significative de à 120 K pour la partie rectale (Fig. 4). Ce changement est typique de la transition de Verwey, au cours de laquelle les spins électroniques de la magnétite se réorganisent. Au-dessus de 120 K, la courbe apparaît moins régulière, mais le niveau d'aimantation est proche du bruit de fond de l'appareil. Les mesures à basses températures indiquent donc la présence de magnétite dans les intestins et la partie rectale de l'animal, localisation qui pose évidemment le problème de son origine « interne » ou « externe ».
Caractéristiques magnétiques de plusieurs morceaux du protée
Characteristics of the remanent magnetism derived from the MPMS and Micromag measurements for several parts of proteus
| Morceaux de l'animal | Dissection | MPMS | Micromag |
| Tête | Crâne et mâchoire | Diamagnétisme | Aucun signal |
| Muscle et mâchoire | |||
| Épiderme du crâne et un peu de système nerveux | Aucun signal | ||
| Yeux et début d'encéphale | |||
| Restes de la tête | Aucun signal | ||
| Peau | |||
| Pattes avant | Pattes avant | Aucun signal | |
| Peau, os et début d'oesophage | |||
| Court (intestin antérieur) | Os, muscle haut des intestins | ||
| Intestin antérieur et glandes annexes | Signal très faible | ||
| Long (intestin moyen) | Peau, muscle, os et intestin moyen | ||
| Peau, muscle, os et intestin moyen | |||
| Reins, pancréas et intestin moyen | Signal très faible | ||
| Corps (intestin postérieur) | Peau, os, muscle | ||
| Peau, os, muscle | |||
| Intestin postérieur et duodénum | Ferrimagnétisme | Signal très faible | |
| Pattes arrière | Peau et muscles avant la queue | Diamagnétisme | Aucun signal |
| Pattes arrière | Diamagnétisme | Aucun signal | |
| Partie rectale | Ferrimagnétisme | Ferrimagnétisme | |
| Os avant la queue | Diamagnétisme | Aucun signal | |
| Queue | Peau et muscle de la queue | ||
| Os | Diamagnétisme |
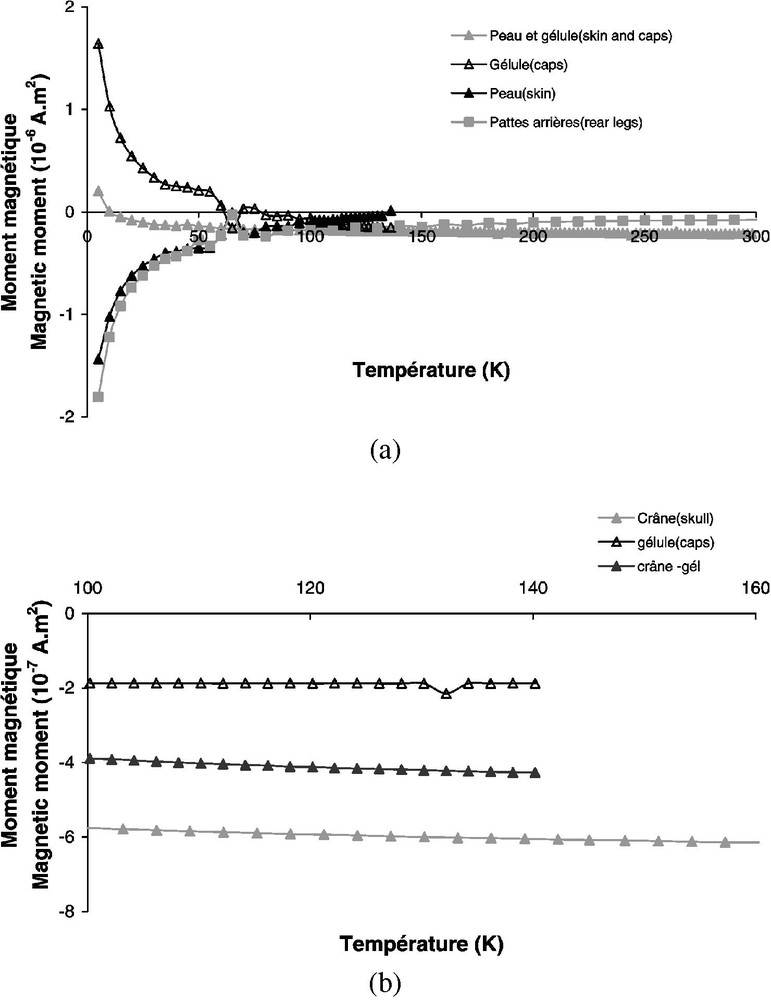
Mesures à basses températures. Variations de l'aimantation à saturation () avec la température. Aucune aimantation n'est plus élevée que celle liée au paramagnétisme de la capsule qui contient l'échantillon.
Low-temperature measurements. Changes in saturation magnetization () with temperature. There is no magnetization larger than the paramagnetism of the capsule used as a container for the sample.
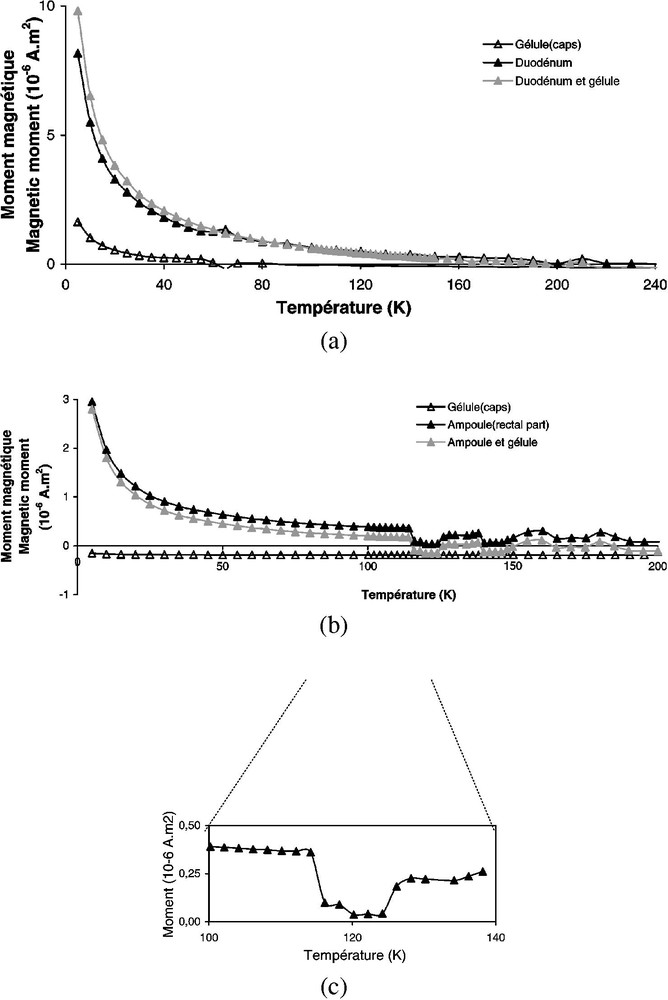
(a) Idem Fig. 3. Dans ce cas, l'intestin a une aimantation caractéristique qui est détectée avant et après soustraction du signal de la capsule contenant l'échantillon. (b) Idem Fig. 3a pour la partie rectale. (c) Signal mesuré entre 100 et 140 K, montrant une chute de associée à la transition de Vervey.
(a) Same as in Fig. 3. In this case, the intestine has a characteristic magnetization, which is detected before and after subtraction of the signal of the capsule containing the sample. (b) Same as in Fig. 3a for the rectal part. (c) Signal measured between 100 and 140 K, showing the drop in associated with the Vervey transition.
3 Origine de la magnétite
Les protées du laboratoire souterrain de Moulis sont élevés dans des aquariums dont le sol est le même que celui de la grotte. De petites particules du sol, et donc de magnétite, peuvent être mises en suspension dans l'eau par les mouvements de l'animal ou ingérés, par exemple avec les vers qui lui servent de nourriture. Si les cristaux de magnétite détectés dans les intestins ont été ingérés par l'animal, ils doivent présenter des caractéristiques similaires aux sédiments. Nous ne pouvons cependant pas attendre une correspondance directe entre les sols des aquariums et les intestins de l'animal, car la nature des particules du sol peut varier sur quelques centimètres. Par ailleurs, le nombre relativement faible de paramètres magnétiques accessibles en raison du très faible niveau d'aimantation rend la comparaison entre les deux familles de grains encore plus difficile. Les mesures effectuées sur les échantillons de sols sont en bon accord avec les paramètres d'aimantation à saturation décrits plus haut pour les protées, et ils présentent une viscosité similaire (environ 15 %). De même, les courbes de désaimantation af de la partie rectale de l'animal (Fig. 5a) et des sédiments (Fig. 5b) présentent les mêmes spectres de champ de déblocage. Ces résultats suggèrent donc que les grains de magnétite trouvés dans les animaux sont en fait des grains provenant du sol de la grotte.
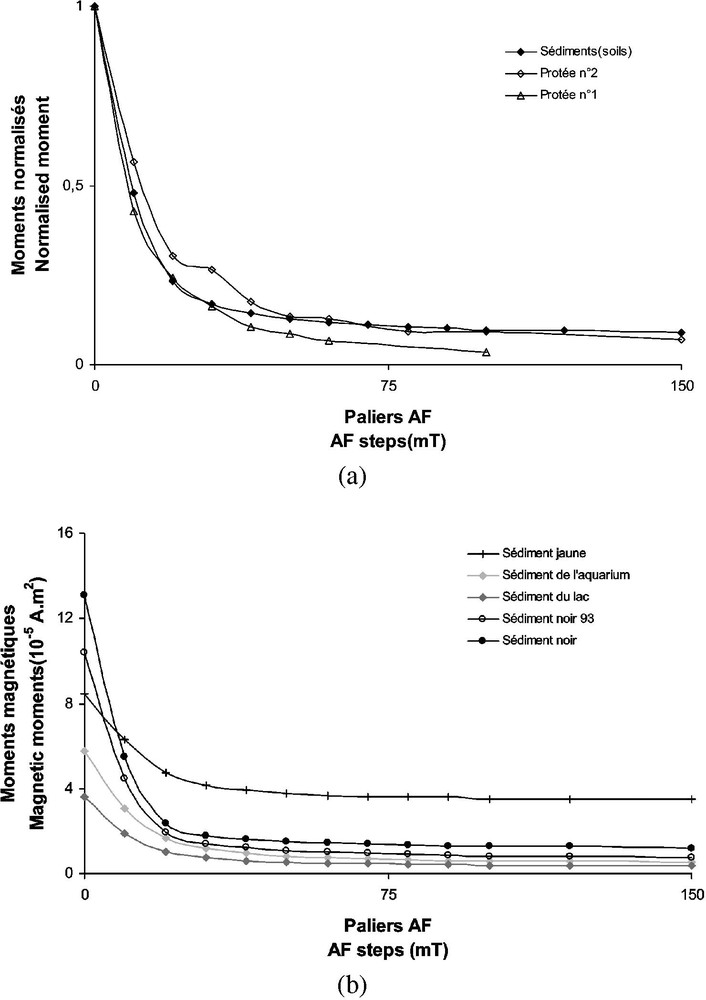
(a) Comparaison entre les courbes de désaimantation AF de la partie rectale de deux protées et celles des sédiments de la grotte. (b) Courbes de désaimantation de divers sédiments trouvés dans la grotte qui auraient été ingérés par l'animal. Il n'y a pas de différence majeure entre la plupart de ces courbes.
(a) Comparison between the stepwise af demagnetization curves for the rectal parts of two proteus and sediments of the cave. (b) Demagnetization curves for a variety of sediments found in the cave that could be ingested by the animal. There is no marked difference between most of the curves.
4 Conclusion
Les mesures d'aimantation rémanente et de susceptibilité en champ fort, effectuées à l'aide d'un magnétomètre cryogénique sur quatre protées prélevés au laboratoire souterrain du CNRS à Moulis, n'ont pas permis de détecter de signal d'aimantation stable dans la tête et dans la partie avant de l'animal. Les seules parties aimantées mesurées dans l'abdomen sont très vraisemblablement dues à l'ingestion de particules magnétiques provenant du sol de la grotte, qui présentent effectivement les mêmes caractéristiques. L'hypothèse la plus probable est que ces grains ont été transportés et ingérés par le protée. Le protée absorbe en effet des vers de vase (des larves de Chironomes) dont la peau est recouverte de particules du sol et qui sont donc ingérées par l'animal. Toutefois, ces résultats ne démontrent pas pour autant l'absence totale de particules aimantées dans le corps de l'animal. En effet, malgré leurs très hautes performances, la sensibilité des deux magnétomètres cryogéniques utilisés demeure presque trois ordres de grandeurs en deçà de l'aimantation d'un unique grain de magnétite monodomaine. Il est toutefois intéressant de souligner que ces résultats contrastent avec les mesures effectuées sur la salamandre Notophtalmus viridescens, qui ont mis en évidence un signal à saturation de 10−9 A m2 dans le corps de ces animaux [1], et dont le magnétotactisme a été formellement reconnu. Bien que l'intensité du champ magnétique terrestre ne soit pas très élevée, on ne peut pas écarter l'hypothèse qu'un petit nombre de grains de magnétite soit à l'origine du magnétotactisme du protée. Dans ce cas, le moment magnétique résultant serait en dessous de la sensibilité de mesure de notre magnétomètre. Pour remédier à ce problème, il faudrait pouvoir placer les bobines de détection à proximité immédiate des échantillons. Il existe à l'heure actuelle un seul appareil capable de cette performance, dont la diffusion prochaine devrait permettre de donner un nouvel essor à ce type de recherche. La détection de grains de magnétite (ou d'autres minéraux) demeure en effet essentielle pour localiser les particules magnétiques, puis pour les analyser en microscopie, afin de valider ou réfuter l'hypothèse du magnétotactisme chez plusieurs animaux.
Remerciements
Les auteurs sont particulièrement reconnaissants à M. Durand, du laboratoire souterrain de Moulis, pour les avoir invités à travailler sur ce sujet et reçus sur place pour les échantillonnages. Ils expriment leur gratitude à Laurent Legrand du Groupe de physique des solides de l'université Paris-6 pour l'utilisation du susceptomètre supraconducteur MPMS et à Isabelle Arnulf de l'UFR de biologie de l'université Paris-7 pour son aide dans la dissection.