

Version française abrégée
Loftusia Brady est un foraminifère benthique du Maastrichtien, connu dans les faciès de plate-forme externe de la Téthys. Il abonde dans les plates-formes arabo-iraniennes et en Turquie, tandis qu'il est rare en Croatie, Serbie, Grèce et Italie et qu'on ne le connaît pas dans des régions plus occidentales (Fig. 1).
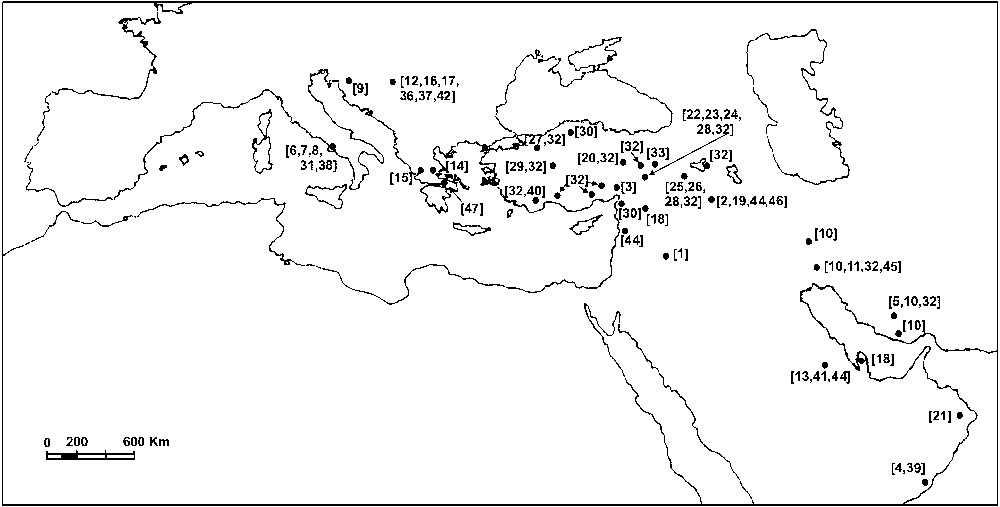
Geographical distribution of the genus Loftusia.
Distribution géographique du genre Loftusia.
Dans la majorité des cas, il est, soit remanié [7,9,14,40], soit présent dans des niveaux qui ne sont pas en continuité stratigraphique avec les couches encadrantes [15,20,22,27,29,42,43].
En Grèce, Loftusia est connu dans deux régions. En Grèce occidentale, dans les monts Valtou (zone du Gavrovo, MV, Fig. 2) [15], Loftusia s'observe dans un calcaire bréchique, en contact par failles avec des calcaires probablement cénomaniens et des brèches paléocènes à madrépores. En Grèce orientale, Loftusia est rencontrée au mont Kassidiaris (MK, Fig. 2) [14], en débris dans un calcaire bioclastique du Maastrichtien supérieur, surmonté en concordance par des sédiments paléocènes.
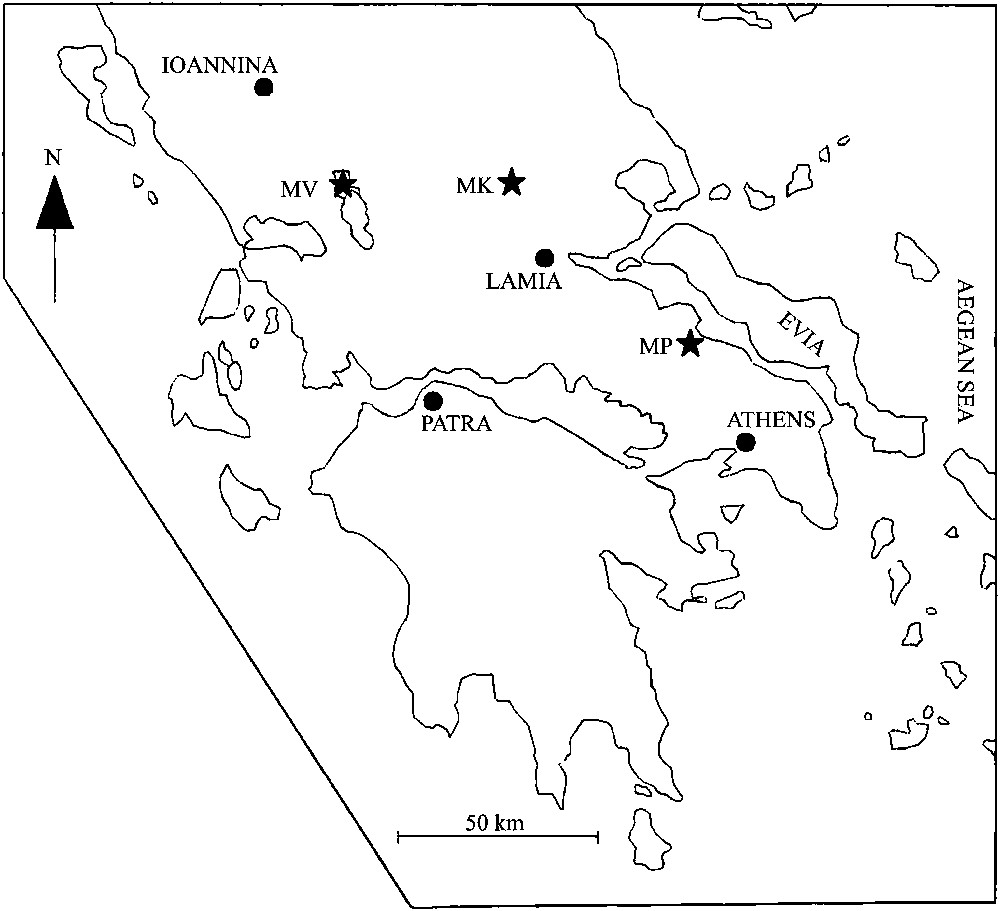
Geographical distribution of the genus Loftusia in Greece. MV: Mounts Valtou, MK: Mount Kassidiaris, MP: Mount Ptoon.
Distribution géographique du genre Loftusia en Grèce. MV : Monts Valtou, MK : mont Kassidiaris, MP : mont Ptoon.
Nous avons découvert, en Grèce orientale, au mont Ptoon (zone subpélagonienne, MP, Fig. 2) [47], in situ, des spécimens de Loftusia dans un niveau de calcaires du Maastrichtien supérieur, en continuité stratigraphique avec les couches inférieures et supérieures d'âge Maastrichtien supérieur également. Il s'agit d'une série carbonatée continue, du Maastrichtien au Paléocène, suivie du flysch. La coupe a été levée en Béotie, le long de la route qui va du village Kokkinon à Akraifnion, 2 km avant ce dernier (Fig. 3).
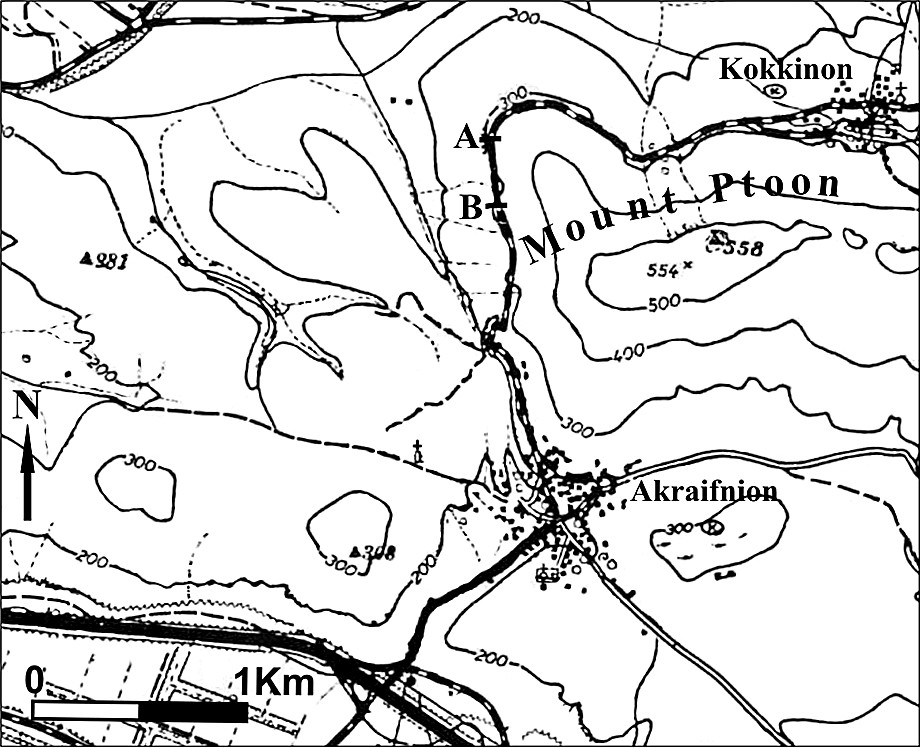
Location of the study section AB.
Situation de la coupe étudiée AB.
La coupe est matérialisée sur la Fig. 4. La série continue comprend, de bas en haut :
- – (A) 20 m de calcaires grainstone–packstone à rudstone à rudistes ;
- – (B) 7 m de calcaires grainstone–packstone à rudstone, datés du Maastrichtien par la présence de Siderolites calcitrapoides Lamarck. Ils contiennent encore des Orbitoides media (d'Archiac), Lepidorbitoides sp., Sirtina sp., Sulcoperculina sp., Goupillaudina sp., Nummofallotia sp., Rotaliidae, Melobesiae et des échinodermes. Dans la partie moyenne de la séquence, un niveau constitué exclusivement par d'abondantes coquilles de rudistes entiers est observé ;
- – (C) 3 m de calcaires packstone–grainstone, datés du Maastrichtien supérieur par Hellenocyclina beotica, associée à Orbitoides media, Orbitoides gensacicus (Leymerie), Lepidorbitoides sp., Siderolites calcitrapoides, Sulcoperculina sp., Nummofallotia sp., Goupillaudina sp., Rotaliidae, Melobesiae, des débris de rudistes, des échinodermes et des coraux. Des coquilles entières de rudistes sont observées dans la partie supérieure de la séquence ;
- – (D) 4 m de calcaires grainstone–rudstone à Loftusia sp. A (Figs. 5a, b et 6a, b). Datée du Maastrichtien supérieur par la présence d'Hellenocyclina beotica Reichel (Figs. 5b et 6b), l'association faunique du niveau (D) inclut encore d'abondants fragments de rudistes, Orbitoides media (Fig. 6b), Orbitoides apiculata Schlumberger, Orbitoides gensacicus (Fig. 6b), Omphalocyclus macroporus (Lamarck) (Fig. 6a et b), Siderolites calcitrapoides, Sulcoperculina sp., et des débris d'échinodermes.
- – (E) 37 m de calcaires wackestone–packstone–grainstone, attribués également au Maastrichtien supérieur et contenant la même association, mais sans Loftusia sp. A ;
- – (F) 13 m de calcaires packstone–wackestone du Paléocène, superposés en continuité et caractérisés par la présence de Globigerinidae, Melobesiae et de gros oursins. Dans la partie supérieure, des intercalations de marnes et de calcaires marneux marquent le passage au flysch.
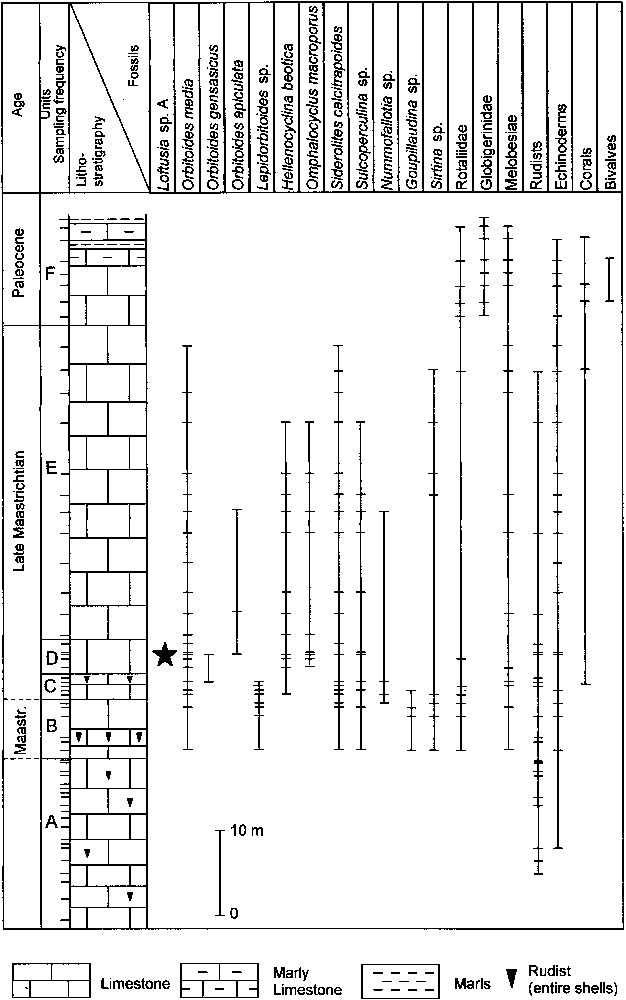
Stratigraphic column.
Colonne stratigraphique.

(a) Loftusia sp. A equatorial section. (b) Hellenocyclina beotica, Loftusia sp. A.
(a) Loftusia sp. A section équatoriale. (b) Hellenocyclina beotica, Loftusia sp. A.
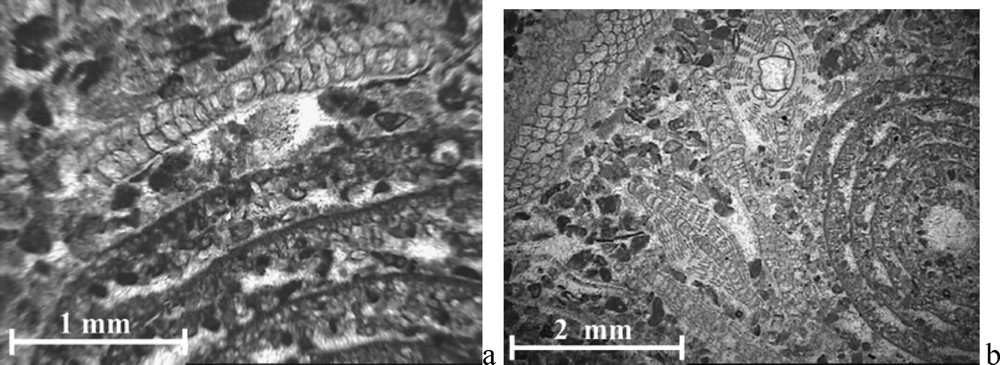
(a) Loftusia sp. A, Omphalocyclus macroporus. (b) Loftusia sp. A, Orbitoides media, O. gensasicus, Omphalocyclus macroporus, Hellenocyclina beotica.
(a) Loftusia sp. A, Omphalocyclus macroporus. (b) Loftusia sp. A, Orbitoides media, O. gensasicus, Omphalocyclus macroporus, Hellenocyclina beotica.
Il faut remarquer que le niveau à Loftusia sp. A est concordant à sa base et à son sommet. L'association faunique du niveau à Loftusia sp. A indique, ainsi que celle des niveaux qui l'encadrent, un âge Maastrichtien supérieur. La localisation de Loftusia dans un mince niveau du Maastrichtien supérieur pourrait expliquer la rareté de ses références en Grèce. L'association faunique montre un environnement de plate-forme externe en relation avec le récif. La proximité du récif est indiquée par l'abondance de débris et des coquilles entières de rudistes dans les niveaux sous-jacents. Cet environnement est connu comme étant favorable au genre Loftusia [15,34].
Vingt lames minces ont été découpées dans les cinq échantillons collectés dans le banc à Loftusia. Malgré l'abondance relative des spécimens (2–26 spécimens par lame mince de 12 cm2), seulement cinq sections (quatre équatoriales et une axiale) conviennent pour les mesures (Tableau 1). Celles-ci montrent que les individus du gisement étudié appartiennent aux espèces de taille moyenne [34,35]. Nous n'avons trouvé que des formes mégalosphériques. Les individus observés présentent des ressemblances, mais aussi des différences avec les espèces L. anatolica Meriç et L. matsumarui Meric & Görmüs (Tableau 2). Nous n'avons pas décrit ici de nouvelle espèce, en raison du très petit nombre d'individus mesurables dans nos échantillons.
Measurements effected on Loftusia from Ptoon (eq: equatorial section, ax: axial section)
Mesures effectuées sur Loftusia du Ptoon (eq : section équatoriale, ax : section axiale)
| L (mm) | D (mm) | nc (mm) |
|
|
|
|||
| Specimen 1 (eq) | – | 3.67–4.3 | 1.2–1.3 | 4 | 0.4 | 0.33–0.37 | 6 | 14 |
| Specimen 2 (eq) | – | 3.1–3.81 | 0.87–0.92 | 5 | 0.27–0.33 | 0.26–0.33 | 4 | 11 |
| Specimen 3 (ax) | 8.48 | 3.97 | 1.24–1.78 | 5 | 0.23 | 0.3–0.33 | – | – |
| Specimen 4 (eq) | – | 3.58–4.85 | 1.36–1.67 | 4.5 | 0.25 | 0.33–0.39 | 6 | 13 |
| Specimen 5 (eq) | – | 5.63–7.44 | 1.8–1.93 | 6.5 | 0.31–0.37 | 0.27–0.3 | 5 | 16 |
Measurements of main parameters for Loftusia species (data from [34])
Mesures des paramètres principaux des espèces de Loftusia (données de [34])
| Species | L (mm) | D (mm) |
|
nc |
|
|
|
||
| L. baykali | 2.75–6.0 | 1.6–3.0 | 1/1.6 | 0.47–1.2 | 2–3 | 0.14–0.47 | 0.23–0.4 | 4–8 | 9–17 |
| L. coxi | 6.1 | 3.3 | 1/2? | 0.33–0.4 | 3 | 0.33–0.55 | 8? | 12–13 | |
| L. harrisoni | 1.6–5.5 | 1.53–5.0 | 1/1.92–1/0.7 | 0.8–2 | 1.5–3 | 0.39–0.61 | 0.3–0.42 | 6–7 | 8–13 |
| 3.2 [17] | 3.2 | 1/1.2–1/1.23 | 12–18 | ||||||
| L. ketini (A) | 1.8–5.3 | 1.4–2.5 | 1/2? | 0.5–1 | 1.5–2.5 | 0.35–0.39 | 0.31–0.39 | 4–5 | 9–10 |
| L. minor (A) | 2.0–7.5 | 0.9–2.9 | 1/2.16–1/3.4 | 0.28–0.7 | 2–4 | 0.15–0.39 | 0.23–0.42 | 8–9 | 13–15 |
| 5.96 | 2.15 | 1/2.79 | |||||||
| L. oktayi | 2.0–4.0 | 2.0–2.9 | 1/1.1 | 0.5–0.94 | 2.5–3 | 0.23–0.42 | 0.23–0.42 | 4–8 | 13–15 |
| L. turcica (A) | 5.2–9.8 | 1.4–2.7 | 1/3.6 | 0.2–0.84 | 2.5 | 0.2–0.4 | 0.24–0.52 | 4–8 | 9–12 |
| 6.7 | 1.87 | ||||||||
| L. anatolica | 16.1–28.8 | 4.7–6.1 | 1/3.09–1/3.9 | 0.9–1.4 | 6–13 | 0.23–0.42 | 0.23–0.47 | 9–11 | 18–23 |
| 1/3.5 [28] | |||||||||
| L. matsumarui | 9.7–16.9 | 3.3–5.3 | 1/2.4–1/3.6 | 0.3–0.7 | 3.5–4.5 | 0.15–0.23 | 4–6 | 16–18 | |
| 1/3.4 [12] | |||||||||
| L. sp. A | 8.48 [1] | 3.1–4.85 | 1/2.1 [1] | 0.87–1.93 | 4–7 [5] | 0.23–0.4 | 0.26–0.39 | 4–6 [5] | 11–16 |
| [4] | [5] | [5] | [5] | [5] | |||||
| 5.63–7.44 | |||||||||
| [1] |
La découverte de Loftusia sp. A in situ dans une série continue de la plate-forme de Grèce orientale (zone subpélagonienne) valide son attribution stratigraphique au Maastrichtien supérieur (coexistence avec Hellenocyclina beotica). Il en est de même en ce qui concerne la présence du genre dans cette plate-forme et dans les Hellénides en général. Les références antérieures concernant les plates-formes du Gavrovo et pélagonienne n'offrent pas cette certitude.
Cette découverte constitue ainsi une nouvelle contribution à la connaissance de la distribution du genre dans la partie occidentale de sa région paléobiogéographique dans la Téthys, où elle n'est pas aussi abondante que dans la région orientale (Fig. 1).
1 Introduction
Loftusia Brady is a Maastrichtian benthic foraminifer, known from outer platform facies of the Tethys. It was found in Italy [6–8,31,38], Croatia [9], Serbia [12,16,17,36,37,42], Greece [14,15,47], Turkey [3,20,22–30,32,33,40], Syria [18,44], Iraq [1,2,19,44,46], Iran [5,10,11,32,45], Saudi Arabia [13,41,44], Qatar [18], Oman [4,21,39] (Fig. 1).
It appears that the genus is abundant in Arabo-Iranian platforms and Turkey, rare in Croatia, Serbia, Greece, and Italy, and totally absent in regions further to the west (Fig. 1).
In the majority of the sites, Loftusia is either transported [7,9,14,40] or enclosed in beds that are not in stratigraphic continuity with the underlying and the overlying formations [15,20,22,27,29,42,43].
In Greece, Loftusia has been found in two localities: in western Greece, on Mounts Valtou (Gavrovo zone, MV, Fig. 2) [15]. Loftusia was found in a breccia that comes in contact, by faults, with limestones probably of Cenomanian age, as well as with Paleocene breccias with Madrepores. In eastern Greece, on Mount Kassidiaris (MK, Fig. 2) [14] Loftusia was found as debris in a bioclastic limestone of Late Maastrichtian age, overlain in conformity by bioclastic sediments of probable Paleocene age.
Recent stratigraphic investigations in eastern Greece, on Mount Ptoon (Subpelagonian zone, MP, Fig. 2) [47] revealed Loftusia, in situ, in a Late Maastrichtian horizon in stratigraphic continuity in a carbonate sequence of Maastrichtian to Paleocene, followed by flysch sedimentation.
2 Stratigraphic description
The section was realized in Boeotia, along the road that from Kokkinon leads to Akraifnion. More specifically it begins at the point where the road ‘Megali Rachi’ is being constructed, 2 km before Akraifnion (Fig. 3).
From bottom to top we observed (Fig. 4):
- – (A) 20 m of grey, medium bedded to massive limestones, grainstone–packstone–rudstone with entire shells and debris of rudists;
- – (B) 7 m of light grey, massive limestones, grainstone–packstone–rudstone of Maastrichtian age, dated by the presence of Siderolites calcitrapoides Lamarck. They also include Orbitoides media (d'Archiac), Lepidorbitoides sp., Sirtina sp., Sulcoperculina sp., Goupillaudina sp., Nummofallotia sp., Rotaliidae, Melobesiae and echinoderms. In the middle of the succession entire shells of rudists constitute the exclusive faunal content;
- – (C) 3 m of light-grey, massive limestones, packstone–grainstone, of Late Maastrichtian age, dated by Hellenocyclina beotica Reichel. Entire shells of rudists are observed in the upper part while debris are observed all over the succession in association with Siderolites calcitrapoides, Orbitoides media, Orbitoides gensacicus (Leymerie), Lepidorbitoides sp., Sirtina sp., Sulcoperculina sp., Nummofallotia sp., Goupillaudina sp., Rotaliidae, Melobesiae, echinoderms and corals;
- – (D) 4 m of light grey, thickly bedded limestones, grainstone–rudstone, of Late Maastrichtian, with debris of rudists, Loftusia sp. A (Figs. 5a and b, 6a and b), Hellenocyclina beotica (Figs. 5b and 6b), Orbitoides media (Fig. 6b), Orbitoides apiculata Schlumberger, Orbitoides gensacicus (Fig. 6b), Omphalocyclus macroporus (Lamarck) (Fig. 6a and b), Siderolites calcitrapoides, Sulcoperculina sp. and echinoderms;
- – (E) 37 m of light-grey, thickly bedded limestones, wackestone–packstone–grainstone, with debris of rudists, Hellenocyclina beotica, Orbitoides media, Orbitoides apiculata, Omphalocyclus macroporus, Sirtina sp., Siderolites calcitrapoides, Sulcoperculina sp., Nummofallotia sp., Melobesiae, Rotaliidae, echinoderms and corals. The continuing presence of Hellenocyclina beotica dates this horizon as Upper Maastrichtian as well;
- – (F) 13 m of light grey, thickly bedded limestones, packstone–wackestone, of Paleocene age, which follow in continuity and are characterized by the presence of Globigerinidae, Rotaliidae, Melobesiae, spines and large shells of echinoderms, corals and debris of bivalves. In the upper part, thin bedded marly limestones and marls are followed by flysch sedimentation.
3 Micropalaeontological and palaeoenvironmental remarks
Order: Foraminiferida Eichwald 1830
Suborder: Textulariina Delage & Hérouard 1896
Superfamily: Loftusiacea Brady 1884
Family: Loftusiidae Brady 1884
Genus: Loftusia Brady 1870 (in Carpenter & Brady 1870)
Loftusia Brady is a benthic planispiral foraminifera. It has a fusiform, ovoid, globose or nautiloid test. The wall of the spire is perforate, calcareous and alveolar with an arenaceous endoskeleton. It is tightly coiled with regular whorls whose thickness increases gradually from the centre to the periphery. Primary, longitudinal, oblique septa divide the whorls into chambers with labyrinthic, endoskeletal structure. The last is composed of radially set pillars. Transverse secondary or partial septa may be formed from the fusion of pillars. Across the primary septa and arranged in transverse rows, there are numerous apertures.
In the study section, Loftusia sp. is found in relative abundance (2 to 26 specimens per sample of 12 cm2) in an Upper Maastrichtian (association with Hellenocyclina beotica) thin horizon of 4 m thickness. It is found neither in the underlying nor in the overlying beds of Late Maastrichtian age as well. That could explain the rarity of the references of the genus in Greece. Furthermore, the fact that the Loftusia horizon is in concordance with the underlying and the overlying beds has to be underlined.
Five samples were collected from the Loftusia horizon and 20 thin sections were made from them. Despite the abundant presence of Loftusia in the sections, only five well-oriented sections were observed (four equatorial and one axial) on which the measurements presented in Table 1 were performed. According to these measurements, the specimens from Mount Ptoon are placed among the medium-sized species of Loftusia [34,35]. Only megalospheric specimens are observed.
The majority of Loftusia species are known from megalospheric individuals (A forms) and only a few species are dimorphic and include both megalospheric (A) and microspheric (B) generations (L. minor Cox, L. ketini Meriç and L. turcica Meriç & Avsar) [34]. Measurements of main parameters for Loftusia species known only from megalospheric forms as well as those of megalospheric (A) forms of dimorphic species are included in Table 2. In the same table are also included measurements effected on our specimens (L. sp. A), for comparative observations.
Based on the observed number of whorls we tend to compare our specimens with the species Loftusia anatolica Meriç and Loftusia matsumarui Meriç & Gormus. More specifically, two of our specimens present 4 and 4.5 whorls respectively, thus falling into the range of values for L. matsumarui, while one presents 6.5 whorls falling into the range of values for Loftusia anatolica. The latter is actually the specimen with the largest diameter. In the remaining two specimens, five whorls were counted. Such a number of whorls is not observed in the above-mentioned species of Loftusia or in any of the species (megalospheric forms) of the genus.
Regarding their diameter, four of our specimens are comparable to L. matsumarui, while one (specimen 5) has an even larger diameter than that of L. anatolica. In fact, its diameter is larger than that of every other so far described species of Loftusia (megalospheric forms). The length was possible to measure in only one specimen and was found smaller than the length of both the above mentioned species. Given that, the measurement of the
Concerning the nucleoconch, it is similar in size and even bigger than that of the typical L. anatolica, while the heights of both first and last tour measure within the range of values for that species. Finally, the number of septa in the first whorl falls into the range of values for L. matsumarui while the number of septa in the last whorl is smaller than that of L. matsumarui.
In summary, according to the above observations, no safe conclusion can be drawn concerning the species to which our specimens should be assigned. They present the greatest resemblance with the species Loftusia anatolica Meriç and Loftusia matsumarui Meriç & Görmüs from which, nevertheless, they appear to have important differences. The observed deviations could not lead us, for the present, to the identification of a new species due to lack of a sufficient number of specimens, therefore, we consider them as L. sp. A.
The Loftusia sp. A horizon overlies a horizon with abundant rudist shells and it also includes Hellenocyclina beotica, Orbitoides media, Orbitoides apiculata, Omphalocyclus macroporus, Siderolites calcitrapoides, Sulcoperculina sp. and echinoderms. The same fossils, but without the Loftusia, are found in the overlying horizon. The facies of the Loftusia horizon, as well as that of the surrounding levels, reflect a depositional environment in the outer platform. The vicinity to the rudist reef is indicated by the abundant debris and entire shells of rudists in the horizon that underlies the one with Loftusia. This environment is in agreement with what is reported in the international literature concerning the biotope of the genus Loftusia [15,34].
4 Conclusions
A Loftusia sp. A horizon of Late Maastrichtian age is for the first time found in situ in Greece (Boeotia), in a continuous undisturbed carbonate sequence of the eastern Greece platform (Subpelagonian zone), of Maastrichtian–Paleocene age, followed by flysch sedimentation. It is restricted in a relatively thin level (4 m), fact that could explain the rarity of references in Greece. Associated with debris of rudists, Hellenocyclina beotica, Orbitoides media, O. apiculata, O. gensacicus, Omphalocyclus macroporus, Siderolites calcitrapoides and echinoderms, it reflects an outer shelf environment in front of the rudist reef. The surrounding in conformity levels, of Late Maastrichtian age as well, include the same microfaunal association, but without Loftusia sp. A. The recovery of Loftusia sp. A in situ in a continuous sequence of the eastern Greece platform (Subpelagonian zone) assures the Upper Maastrichtian attribution of this probably new species, as well as the indubitable presence of the genus in this platform and the Hellenids in general. Its reports from the Gavrovo [15] and the Pelagonian [14] zones could not provide this certainty. Moreover, this fact provides new data on the distribution of the genus in the western part of its palaeobiogeographic area in the Tethyan region, where it is not as abundant as in the eastern part (Fig. 1).
Vous devez vous connecter pour continuer.
S'authentifier