

1 Introduction
Very recent studies were carried out in Brazil, dealing with the indicators of physical properties of soils (Guimarães Santos et al., 2011) and with the soils’ structure as related to water movement (Bustillo et al., 2011). The study here presented concerns another subject of soil structure in the Brazilian ferralsols.
In the course of this decade, Balbino et al. (2001, 2002a, 2002b), Volland-Tuduri et al. (2004, 2005) and Reatto et al. (2007) have improved the characterisation of the physical properties of Brazilian ferralsols, particularly their micro-aggregation and porosity. These ferralsols (FAO, 2006), latossolos in the Brazilian taxonomy (EMBRAPA, 1999), occupy nearly half the surface area in the Cerrado biome, and a considerable part of this land in tropical regions has been brought into cultivation over the last century. The most recent descriptions (Balbino et al., 2001; Neufeldt et al., 1999; Volland-Tuduri et al., 2005; Westerhof et al., 1999) make it clear that, under natural vegetation, these ferralsols have a poorly-developed macrostructure and strong microstructure with micro-aggregates (50–300 μm in size) resulting from termite activity (Eschenbrenner, 1986; Oliveira et al., 2000; Reatto et al., 2009; Trappnel and Webster, 1986). However, larger aggregates of 500 to 1000 μm, arranged in small clods from 2 to 20 mm in size, have also been reported (Volland-Tuduri et al., 2005). These aggregates were attributed to geophagous earthworm activity following deforestation (Balbino et al., 2001; Chauvel et al., 1999; Grimaldi et al., 1993). Classic results on the effect of endogeic earthworms under tropical conditions have shown the formation of water-stable macro-aggregates. All the studies emphasized the enhancing and weakening effects on soil structure according to soil texture (Blanchart et al., 1999). General effects such as horizon compaction in specific situations were reported (Chauvel et al., 1999), but other results showed how the same earthworm species, Pontoscolex corethrurus, enhanced porosity and decompaction (Blanchart et al., 1999).
Most ferralsols feature fine porosity due to the assemblage of clay particles, mainly kaolinite, but the porosity resulting from clay packing varies when gibbsite is associated with the kaolinite (Balbino et al., 2002b). When some of these soils are cultivated, particularly the clayey-sandy or clayey ferralsols, there is a decrease in water retained between 1 and 3 kPa, and hence a decrease in the water available from macropore storage (Balbino et al., 2001). Consequently, there is considerable variation in the amount of available water, with values ranging from 0.048 to 0.189 g g−1. These variations in macro-porosity are associated with the way the soil is managed, rather than with clay content or bulk density (Balbino et al., 2002a). Earthworm populations are known to be very sensitive to changes in soil management (Lavelle and Spain, 2001). In a first step (Fregonezi et al., 2001), we presented results showing significant variations in macromorphology and bulk density down to the upper part of the B horizon in a yellow ferralsol under different types of cultivated pasture. In this context, the aim of the present study is to analyse which physical soil characteristics were affected by transformations and to address the role of soil macrofauna.
2 The site and its context
The study area, on a commercial farm, was located in the center of Brazil (19°20'S, 48°10′W, elevation 900 m) on a gently undulating plateau, with no sign of runoff or erosion on the surface (Fregonezi et al., 2001). The mean annual precipitation is 1574 mm, with 6–7 months of rain and a marked dry season. The mean annual temperature ranges from 20 to 24 °C. Clayey soils (Table 1) were identified as being yellow/red gibbsitic ferralsols (FAO, 2006) or dystrophic latosols (EMBRAPA, 1999) with kaolinite and gibbsite content ranging from 0.43 to 0.47 g g−1 of soil and from 0.25 to 0.30 g g−1 of soil respectively. Goethite, hematite and quartz were also found.
Caractérisations physicochimiques des profils de sols étudiés sous végétation naturelle (CER), pâturage ancien à Brachiaria (CB), pâturage renouvelé à Brachiaria (RB) et pâturage renouvelé à Bachiaria-Stylosanthes (RBS).
| Site | Soil layer | Ferralic horizon | Clay | Db | pH w | CEC | Corg | |
| m | Starting from the soil surface at (m) | Colour (dry) | g kg−1 | g cm−3 | cmolckg−1 | g kg−1 | ||
| CER | 0–0.20 | 582 | 0.79 | 4.6 | 6.7 | 15.1 | ||
| 0.20–0.40 | – | 0.87 | 4.4 | 5.2 | 10.6 | |||
| 0.50–0.70 | 0.50 to 0.55 | 5 YR 5/6 | 640 | – | 4.5 | 3.9 | 8.1 | |
| 1.0–1.2 | – | 0.66 | 4.9 | 3.1 | 6.1 | |||
| 1.50–1.70 | – | – | 4.3 | 2.3 | 5.2 | |||
| 1.9–2.0 | – | – | 5.2 | 2.2 | 4.9 | |||
| CB | 0–0.20 | 506 | 0.87 | 5.3 | 6.9 | 16.4 | ||
| 0.20–0.40 | – | 0.82 | 4.9 | 6.4 | 14.8 | |||
| 0.50–0.70 | 0.50 | 5 YR 5/8 | 620 | – | 4.9 | 4.2 | 9.3 | |
| 1.2–1.3 | – | 0.69 | ||||||
| RB | 0–0.10 | 614 | 0.82 | 5.3 | 7.3 | 19.1 | ||
| 0.10–0.35 | – | – | 5.2 | 7.0 | 16.9 | |||
| 0.35–0.60 | 0.50 to 0.60 | 5 YR 5/8 | 628 | 0.82 | 4.9 | 4.8 | 11.2 | |
| 0.80–0.90 | – | 0.72 | ||||||
| RBS | 0–0.15 | 590 | 0.81 | 5.7 | 7.3 | 20.0 | ||
| 0.15–0.25 | – | 0.86 | 6.5 | 4.4 | 13.9 | |||
| 0.40–0.50 | 0.50 to 0.55 | 5 YR 5/8 | 645 | – | 4.7 | 4.5 | 10.2 | |
| 0.90–1.0 | – | 0.76 | – | – | – |
Observations were made at the end of the rainy season in three types of cultivated pastures and in a natural vegetation area: (i) a conventional, 20-year-old pure Brachiaria decumbens pasture, denoted CB, with low livestock carrying capacity and showing areas of exposed soil due to overgrazing; (ii) a 4-year old renewed pure Brachiaria decumbens pasture (RB) established on an old pasture which was part of the CB plots similar to (i) after soil preparation, liming and fertilisation; (iii) the RBS plot, a 4-year old renewed Brachiaria decumbens associated with the legume Stylosanthes guianensis cv. Mineirão; and (iv) the CER plot, an area of Cerrado vegetation and a typical open semi-deciduous forest-savannah used as a control. To prevent overgrazing of RB and RBS pastures, the farmer controlled animal numbers. All plots were in the same topographic position and elevation; pasture plots were adjacent and the CER plot was located at a distance of 1 km from the pastures.
3 Description of soils and analytical methods
In each plot, five monoliths measuring 0.25 × 0.25 m were sampled from the soil surface to 0.30 m deep, spaced 10 m apart (Anderson and Ingram, 1993). In the soil volume extracted, macroinvertebrates were counted and their biomass determined in the litter, 0–0.1, 0.1–0.2 and 0.2–0.3 m layers.
The soil morphology was described using the cultural profile method adapted to tropical conditions (Tavares Filho et al., 1999), with two pits per treatment. Ten different types of structure, i.e. homogeneous morphological units, were identified (Fig. 1 and Table 2). Undisturbed samples, a few dm3 in volume, were collected from different morphological types for physical analysis (Table 3). Moisture conditions were analysed for structures 1, 2, 3, 4, 7 and 9 (Table 4). Blocks of a few decimetres in size were sampled to take precise structure samples to the laboratory for analytical study. For each morphological type observed, samples were taken using 50 cm3 cylinders to establish water retention curves using the drying method on pressure plate apparatus (Richards and Fireman, 1943). Tensions of −1, −3, −10, −33, −1500 kPa were applied, with at least three replications for each tension. The volumetric water content was calculated.

Schematic ferralsol profile morphology and structural features under Cerrado natural vegetation (CER), old Brachiaria pasture (CB), renewed pure Brachiaria pasture (RB) and renewed Bachiaria-Stylosanthes pasture (RBS). See Table 2 for structure description.
Morphologie schématique des profils de sols et traits structuraux sous végétation naturelle (CER), pâturage ancien à Brachiaria (CB), pâturage renouvelé à Brachiaria (RB) et pâturage renouvelé à Bachiaria-Stylosanthes (RBS). Le Tableau 2 renseigne les descriptions des structures.
Morphologie de la structure et principaux traits décrits dans les profils de sols.
| Symbol | Homogenous structural volume (layer) | Internal features of the structural volume | Principal faunal features |
| 1 | Strong fine to very-fine granular (Bw horizon of ferralsols) | Microgranular structure and inter-micro-aggregates voids | Some termites and ants galeries and channels |
| 2 | In a matrix of fine granular structure, random dense irregular sub-rounded volumes | 2 types of fine to coarse aggregates, a/ dense with no visible inner porosity, b/ dense with fine to very-fine porosity, altered by termite activity | Termites galeries and voids |
| 3 | Strong fine granular structure and a weak to moderate medium subangular blocky structure | The blocky str. is composed by aggregates with fine to coarse porosity | Intense termite activity (channels and voids) |
| 4 | Subangular blocky structure, macro-porosity due to cracks in all directions | Small and medium rounded aggregates which can be isolated from coarse dense irregular sub-rounded volumes. Aggregates present a fine to coarse internal porosity. High root density | Earthworms in activity, dejections, galeries; termite galeries and channels |
| 5 | Granular, medium to coarse, no cohesion, macro-porosity due to inter-aggregates voids | Porous aggregates are associated by a dense roots system | Activity of worms, dejections and galeries |
| 6 | Subangular blocky | Macro-porosity by voids and cracks. Fine to medium aggregates without porosity | Fine termites galeries |
| 7 | Coarse angular blocky to fine and coarse subangular blocky, cracks | Non-porous aggregates | Non-observed faunal activity |
| 8 | Large platy (laminated) | No porosity | |
| 9 | Subangular blocky with random volumes of strong fine to very-fine granular | Porous rough aggregates. Higher termite activity in comparison to (2) | Termites and ants cavities, vertical channels |
| 10 | Fine to medium subangular blocky to granular fine, medium and coarse | Porosity by cracks and inter-aggregates, two types of aggregates, a/ porous and b/ dense, round rough faces porous associated to high root density | Earthworms dejections, high activity of termites and ants |
Propriétés physiques des structures : analyse granulométrique, densité réelle (Dp), densité apparente des volumes structuraux (Db vol), densité apparente des agrégats (Db ag), volume poral total de la structure (Vpt), volume poral total des agrégats (Vpt ag) et proportion de la masse du matériau micro-agrégé dans les agrégats (Φ ag).
| Site | Layer | Structurea | Sand | Silt | Clay | Dp | Db vol | Db ag | Vpt | Vpt ag | Φagb |
| m | g kg−1 | g cm−3 | cm3 g−1 | ||||||||
| CER | 0–0.05 | 5 | 340 | 95 | 563 | – | 0.84 0.04 | 1.06 0.07 | – | – | 0.34 |
| 0.05–0.15 | 4 | 280 | 120 | 600 | 2.67 | 0.99 0.03 | 1.12 0.02 | 0.629 | 0.581 | 0.24 | |
| 0.15–0.35 | 3 | 290 | 138 | 573 | 2.60 | 1.03 0.12 | 1.09 0.02 | 0.604 | 0.581 | 0.29 | |
| 0.35–0.50 | 2 | 225 | 85 | 690 | 2.60 | 1.04 – | 1.11 0.03 | 0.602 | 0.573 | 0.25 | |
| 0.50–0.70 | 1 | 243 | 118 | 640 | 2.63 | 0.81 0.08 | 1.02 0.03 | 0.693 | 0.612 | 0.41 | |
| CB | 0–0.05 | 8 | – | – | – | – | – | – | – | – | |
| 0.05–0.20 | 6 | 268 | 228 | 505 | 2.60 | 0.99 – | 1.24 0.04 | 0.617 | 0.523 | 0.07 | |
| 0.05–0.20 | 7 | 280 | 213 | 508 | 2.63 | 1.09 0.10 | 1.31 0.03 | 0.584 | 0.502 | 0.00 | |
| 0.20–0.50 | 2 | 220 | 140 | 640 | 2.63 | 1.13 0.03 | 1.15 0.05 | 0.571 | 0.563 | 0.19 | |
| 0.50–0.70 | 1 | 223 | 128 | 620 | 2.63 | 0.97 0.06 | 0.98 0.10 | 0.632 | 0.627 | 0.49 | |
| RB | 0–0.05 | 10 | – | – | – | – | – | – | – | – | – |
| 0.05–0.35 | 4 | 230 | 156 | 614 | 2.56 | 1.04 0.02 | 1.12 – | 0.596 | 0.563 | 0.24 | |
| 0.35–0.55 | 9 | 243 | 130 | 628 | 2.60 | 0.89 0.10 | 1.08 0.03 | 0.658 | 0.585 | 0.30 | |
| 0.55–0.70 | 1 | 255 | 130 | 615 | 2.63 | 0.91 0.08 | 0.97 0.04 | 0.654 | 0.631 | 0.51 | |
| RBS | 0–0.10 | 10 | 290 | 145 | 565 | – | 0.97 – | 1.36 0.04 | – | – | 0.00 |
| 0.10–0.50 | 4 | 220 | 165 | 615 | 2.63 | 0.99 0.03 | 1.32 0.02 | 0.622 | 0.498 | 0.00 | |
| 0.35–0.50 | 9 | 178 | 178 | 645 | 2.63 | 1.07 0.07 | 1.21 0.05 | 0.595 | 0.540 | 0.11 | |
| 0.50–0.70 | 1 | – | – | – | 2.63 | – | – | – | – |
b Φagg: mass proportion of micro-aggregates < 0.84 mm class fraction calculated after Volland-Tuduri et al. (2005).
Teneur en eau volumique (cm−3 cm−3) dans les différents volumes structuraux pour les différentes classes de tensions.
| Structuresa by layer and site | Class tension (kPa) | |||
| −1 to −3 | −3 to −10 | −10 to –33 | −33 to −1500 | |
| Layer 0.05–0.35 m | ||||
| 4 - RB | 0.063 0.010 AB | 0.102 0.016 A | 0.033 0.002 B | 0.034 0.005 B |
| 4 - RBS | 0.012 0.004 B | 0.053 0.003 C | 0.019 0.009 B | 0.039 0.001 B |
| 4 - CER | 0.013 0.004 B | 0.064 0.013 BC | 0.020 0.007 B | 0.053 0.011 B |
| 3 - CER | 0.089 0.053 A | 0.067 0.003 BC | 0.034 0.016 AB | 0.051 0.0003 B |
| 7 - CB | 0.012 0.004 B | 0.092 0.011 AB | 0.061 0.006 A | 0.050 0.006 A |
| Layer 0.35–0.50 m | ||||
| 9 - RB | 0.100 0.022 A | 0.109 0.003 A | 0.027 0.002 B | 0.034 0.003 B |
| 9 - RBS | 0.022 0.003 B | 0.103 0.009 A | 0.032 0.008 AB | 0.039 0.006 AB |
| 2 - CERb | 0.019 B | 0.099 A | 0.021 B | 0.053 A |
| 2 - CB | 0.011 0.001 B | 0.097 0.012 A | 0.048 0.011 A | 0.049 0.012 AB |
| Layer 0.50–0.70 m | ||||
| 1 - CER | 0.028 0.008 A | 0.112 0.037 A | 0.022 0.004 A | 0.026 0.004 A |
| 1 - All pastures (CB, RB, RBS) | 0.050 0.034 A | 0.123 0.013 A | 0.043 0.009 A | 0.032 0.018 A |
a Table 2.
b No replicates.
Bulk densities of the layers and structural volumes were measured using the cylinder method and the proportions of structural volume types determined. Particle density was determined with a pycnometer and particle size distribution using the pipette method (EMBRAPA, 1997). The bulk density of the aggregate classes was measured using the clod method (Mathieu and Pieltain, 1998).
The results were tested using Analysis of Variance and Comparison of Means (Tukey) at P < 0.01. Soil bulk density, bulk density of structural features and bulk density of aggregates and all macrofauna data, biomass and density were subjected to principal component analysis (PCA), using the SPAD software program (SPAD, 1998), retaining only those components with eigenvalues > 1.
4 Results
4.1 Morphology and porosity of the structural elements observed
Under natural conditions, from the upper limit of the Bw horizon to the surface (Fig. 1 and Table 2), the reference soil (CER) presented a characteristic succession of layers, with a strong fine to very-fine granular structure (1), two intermediate layers in which a weak to moderate medium subangular blocky structure developed in the granular matrix (2 and 3), and the upper layers showed typical subangular blocky (4) and granular structures produced by faunal activity (5). We compared the structures found between 0.05 and 0.50 m depth, where the macromorphological transformations are the most significant (Fregonezi et al., 2001) (Fig. 1, Tables 2 and 3). We found that the soil of the old CB pasture, compared with that under native vegetation (CER) had undergone major modifications down to a depth of 0.5 m, resulting in a general loss of visible porosity and a change in the type of subangular blocky (4) and granular structures (5) to coarse to fine angular blocky and coarse subangular blocky with cracks (7), and a platy structure in the upper layers (8) (Fig. 1, Table 2). The transformation of this old CB pasture into new pastures (RB or RBS), observed after 4 years of development, resulted in recovery of a fine porosity, with the formation of new small and medium-sized rounded aggregates included in a subangular blocky structure, and a fine to medium subangular blocky to fine to coarse granular structure in the upper layer (structures 4 and 10, Table 2).
4.2 Water contents obtained at −1 to −1500 kPa
The volumetric water contents of the structural volumes are shown in Table 4. The relationship between tension applied and pore class (given in equivalent pore radius) was as follows: −1 and −3 kPa = 150–47 μm; −3 and −10 kPa = 47–15 μm; −10 and −33 kPa = 15–4.7 μm ; −33 and −1500 kPa = 4.7–0.094 μm. In the 0.5–0.35 m layer and for the pore class 47–150 μm (−1 to −3 kPa), structural volumes 3 of CER and 4 of RB showed the highest values; for the 15–47 μm pores (−3 to −10 kPa), the highest values were in structural volumes 4 of RB and 7 of CB; for the 4.7–15 μm pores (−10 to −33 kPa) structural volumes 7 of CB and 3 of CER showed significant variation; finally, for the finest pores, structural volumes 3 and 4 of CER and 7 of CB were notable but not significantly different.
In the 0.35–0.50 m layer, the values obtained in structures 9 of RB were significantly the highest in the porosity range −1 to −3 kPa (47–150 μm), whereas there were no particularly significant differences in the other pore classes.
In the micro-aggregated structures, structure 1 (CER, CB, RB and RBS), the values were equivalent in all pore range.
4.3 Soil fauna
The largest biomasses, observed in the monoliths sampling, were founded in the CER, followed by those in the RB and RBS sites (Table 5). Termites and ants were present everywhere, whereas earthworms, which were virtually absent in the CER and CB (where only a few individuals were observed) were well represented in the RB and RBS plots. Results showed that earthworm density was particularly high in the 0.1–0.2 m layer in the RBS plot. The PCA correlation matrix showed a negative correlation within the Db values and termite biomass values (Table 6). The PCA produced an optimized factorial model with three factors accounting for 79.8% of the total variance (Fig. 2). Factor 1 explained 41.0% of the variance and was negatively correlated with soil bulk density (−0.80), aggregate bulk density (−0.60) and positively with Isoptera biomass and density (0.95; 0.65) and biomass and density of Formicidae and Coleoptera. Thus, factor 1 represented the effects of the soil Isoptera, Formicidae and Coleoptera on bulk density, since 58% of the data on factor 1 were contributed by their biomass. Factor 1 clearly differentiated the contribution of insects (termites, ants, coleoptera) from that of earthworms. Factor 2, accounting for 20.3% of the variance, was positively correlated with earthworm biomass (0.61) and aggregate bulk density (Db ag); and negatively with bulk density of structural features (−0.68). The Db ag is accounted for 45.5% in factor 2. The faunal survey in RBS, 0.1–0.2 layer, shows high earthworm density, in contrast to the RB 0–0.1 layer in which insects were dominant. Factor 2 differentiated CER, in terms of the contribution of termites and ants, from the pasture upper layers.
Moyenne et erreur standard (en italique) des densités (D, ind dm−3) et des biomasses (B, mg dm−3) de macro-invertébrés du sol des quatre sites étudiés.
| CER | CB | RB | RBS | |||||
| D | B | D | B | D | B | D | B | |
| Isoptera | 39.99 10.46 | 75.89 44.03 | 8.54 2.71 | 2.66 2.27 | 53.56 19.07 | 60.22 25.92 | 15.86 10.17 | 21.07 17.23 |
| Formicidae | 1.26 0.73 | 0.36 0.19 | 0.10 0.05 | 0.07 0.04 | 2.36 1.59 | 0.88 0.43 | 0.96 0.60 | 0.45 0.38 |
| Earthworm | 0 | 0 | 0 | 0 | 0.09 0.02 | 0.19 0.10 | 0.49 0.20 | 6.55 3.38 |
| L. coleopteraa | 0.15 0.04 | 0.92 0.36 | 0.10 0.04 | 0.40 0.27 | 0.31 0.04 | 2.36 0.42 | 0.25 0.07 | 0.57 0.24 |
| A. coleopteraa | 0.01 0.01 | 0 | 0.06 0.04 | 0.04 0.04 | 0.14 0.03 | 2.17 1.93 | 0.18 0.06 | 0.38 0.18 |
| Others | 0.49 0.11 | 5.01 4.80 | 0.49 0.11 | 2.00 0.16 | 0.23 0.05 | 3.67 1.31 | 0.23 0.05 | 1.72 0.53 |
| ∑ | 41.43 10.16 | 82.19 42.66 | 9.29 2.87 | 5.17 2.14 | 56.68 19.95 | 69.50 27.40 | 17.97 10.94 | 30.74 20.93 |
a A: adults (adultes); L: larvae (larves).
Corrélations des variables de l’ACP.
| Soil fauna | Bulk density | ||
| D b | Db vol | Db ag | |
| Biomass | |||
| Termite | −0.748 | −0.600 | −0.640 |
| Ant | −0.548 | −0.294 | −0.189 |
| Earthworm | 0.158 | −0.146 | 0.554 |
| L. coleoptera | −0.282 | −0.143 | −0.273 |
| A. coleoptera | −0.160 | 0.098 | −0.142 |
| Density | |||
| Termite | −0.438 | −0.758 | −0.393 |
| Ant | −0.505 | −0.291 | −0.297 |
| Earthworm | −0.006 | −0.143 | 0.548 |
| L. coleoptera | −0.330 | −0.220 | −0.010 |
| A. coleoptera | −0.289 | 0.006 | 0.364 |
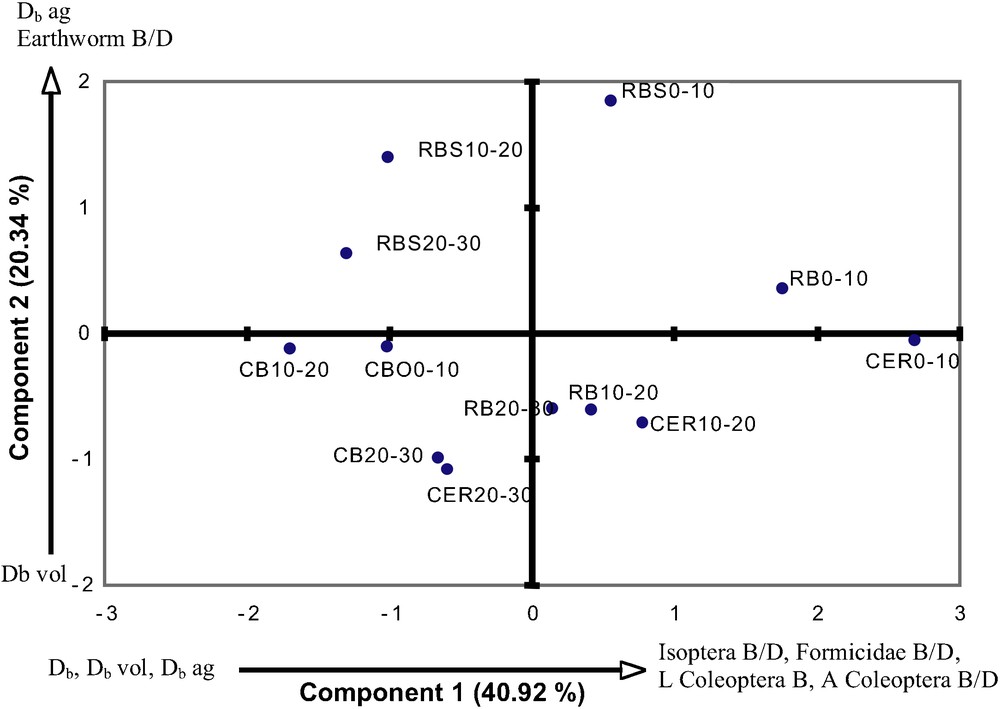
Principal component analysis of soil layers bulk density (Db), structural volume bulk density (Db vol), aggregate bulk density (Db ag) and fauna data (Isoptera, Formicidae, adults and larvae of Coleoptera, Earthworm; B: biomass, D: density) measured in the 0–0.3 m layers. The percentage of variance is noted for each component.
Analyse en composantes principales des variables densité apparente des horizons (Db), densité apparente des volumes structuraux (Db vol), densité apparente des agrégats (Db ag) et de la faune du sol (Isoptera, Formicidae, adultes et larves de Coleoptera, Earthworm ; B : biomasse, D : densité), mesurées dans les couches 0–0,3 m. Le pourcentage de variance est noté pour chaque composante.
5 Discussion
5.1 Soil and structure
Results showed a fairly good textural homogeneity at each depth among the ferralsols studied; values for particle density and soil colour confirmed this homogeneity (Tables 1 and 3). Textural variations were caused by material brought back up by termites and ants and subsequent mixing, which we were able to observe in the field. Brazilian ferralsols are normally known for featuring weakly developed fine granular macrostructure, which expresses a stable microstructure with aggregates of 80 to 350 μm in size (Furian et al., 1999; Lima and Anderson, 1997; Neufeldt et al., 1999). A more detailed analysis was performed to separate this microstructure into individual structural units (Balbino et al., 2001), and to quantify them (Volland-Tuduri et al., 2005). The structures of the CER soil were similar to those previously observed (Balbino et al., 2002a; Reatto et al., 2007; Volland-Tuduri et al., 2005).
5.2 Structural evolution within pasture cultivation
Some studies show that when ferralsols are transformed into pasture or intensively farmed, the structure is altered and bulk density values increase (Curmi et al., 1993; Kondo and Dias Junior, 1999; Stone and Silveira, 1978; Tavares Filho and Tessier, 1998). This was not observed in our study, bulk density values not showing any significant variation at the soil horizon scale for the different types of usage (Table 1). However, if we examine the bulk density values of the structures (Dbvol), results showed an increase in Dbvol from CER to CB, and a drop from CB to the two recent pastures, RB and RBS (Table 3).
The changes that occur can be defined by calculating pore volumes. The total pore volume of the structures (Vpt) was calculated based on particle density (Dp) and bulk density (Db) (Table 3). It has been suggested that there is a close linear relationship between the pore volume due to the assemblage of clay particles and the clay content in ferralsols (Balbino et al., 2002b). Using this relationship (Table 3), we estimated that the mean pore volume associated with the clay of the soil studied is 0.185 ± 0.013 cm3 g−1. This calculation shows that 28 and 32% of the total soil porosity (Vpt) were due to the organisation of the clay particles. At the soil surface, the changes that had taken place in the old pasture (CB) compared with the soil under natural conditions (CER) are related to the development of physical barriers to infiltration, with laminar structures at the surface (Fig. 1, Table 2) and a noticeable reduction of pore volumes above 47 μm down to a depth of 0.35 m (Table 4). This result confirms earlier observations (Balbino et al., 2001; Carvalho et al., 1998; Grimaldi et al., 1993). It is notable that the transformation of this ancient pasture to RBS and RB showed a similar change in structures 4, similar to the structure 4 described for the CER (Fig. 1, Tables 2, 3 and 4). In the 0.35–0.50 m layers, the renewal of CB into RB significantly changed the porosity > 47 μm and contributed to a total porosity greater than RBS (Table 4). In the micro-aggregate layers, we found a wide variation of the increase in the porosity for the pore classes > 15 μm. This is explained by the presence of small aggregates whose porosity was comparable in all the plots (Table 3), but the total porosity of the whole horizon was highest in CER (0.693 dm3 kg−1), as shown by the bulk density values (Table 3). Much of the intra-aggregate and inter-clod porosity is due to vigorous rooting (Table 2). In the RBS, there was a clear reduction in the internal porosity of the macro-aggregates. This was one of the characteristics of these coarse aggregates, previously described as compact to very compact and whose bulk density values (Dbag) were higher than those of the structural volume from which they came (columns Dbvol and Dbag, Table 3). Consequently, the water supply from these soils is provided by a coarse network of inter-clod voids and aggregates characteristic of an assemblage that is piling up.
5.3 Structure and soil fauna
Coarse aggregates, described as compact to very compact (Tables 2 and 4), are structures resulting from the activities of two species of earthworm observed which have strongly colonised these plots (Fig. 2, Table 5). This result is in line with previous work (Blanchart et al., 1997, 1999) describing compacting worms. The colonisation was compatible with the higher carbon contents observed in the surface horizon of RB and RBS in comparison to the other sites (Table 1).
Usually, cultivating the soil leads to the disappearance of macropores formed by biological activity (Borges et al., 1999). In ferralsols on basalt, development of the structure into micro-aggregates at the surface and subsurface following farming practices has been demonstrated (Tavares Filho, 1995). If macropore shrinkage can be attributed to the introduction of cultivation, our study clearly shows that the drop in porosity is largely offset by rearrangement into elementary structures, the macro-aggregates observed in structures 4 and 10 (Table 2). What therefore characterised these ferralsols was this juxtaposition of a microstructure with coarse aggregates. The mass proportion of micro-aggregates (< 0.84 mm, Table 3) varied with bulk density (Φ<0.84 = 1.97 [1/Db] − 1.52) (Volland-Tuduri et al., 2005). The mass proportion of micro-aggregates, denoted 1 (Fig. 1, Table 2), estimated using this equation, indicated the absence of micro-aggregate structures in the macro-aggregates of RBS and CB according to the Dbag values, and their significant presence in the structures of CER and in the deep layers of CER, CB and RB (Φag in Table 3). Their absence from RBS was due to earthworms that sorted and totally reorganised the microparticles as they passed through the intestine and were compacted during excretion (Barois et al., 1993; Chapuis-Lardy et al., 1998). The insects, termites and ants tended to create porosity in transporting the soil and tunnelling, thereby helping to reduce soil bulk density, as illustrated by the PCA (Fig. 2). In contrast, endogeic earthworms promoted the formation of large stable macro-aggregates (Lavelle and Spain, 2001) that eventually led us observe a close relationship between the bulk density of the soil layers and the structure of the earthworm community (Rossi, 2003). This was observed in PCA and the correlation matrix where the bulk density values of soil layers and aggregates were negatively correlated with termite biomass and density (Table 6).
6 Conclusions
We can see that transformations in structure are accompanied by changes in porosity and particularly pore size. Comparing the soil under natural vegetation with soil under pasture, these changes are substantial in the surface layer. Under pasture on which a satisfactory amount of organic matter is returned to the soil, we found that these changes affecting the structure and pores are due to compact agglomerates produced by earthworms. This reduces the available soil water, but the very loose assemblage of the whole structure maintains very strong void porosity, ensuring good infiltration and supply of water to deeper layers. In the final analysis, the variations observed between pastures are purely due to the abundance of these structures, partly attributable to soil-feeding earthworms. In addition, the high plasticity of these soils should be underlined, since the structures have been favourably modified in the medium term, inasmuch as the observation of ancient pasture has revealed serious constraints to good root growth.