1 Introduction
The landscape is the background from which the habitats used by animals and humans derive. Studying the interaction between the fauna and the environment requires therefore, in the first place, understanding the ways in which landscapes evolve through space and time. Earth scientists study this evolution at multiple scales while investigating the underlying geodynamical processes. Here we provide examples demonstrating that the knowledge and techniques derived from Earth sciences can be adapted to research in other fields, such as palaeoanthropology and archaeology.
Combining evidence in the fossil and archaeological record with a better knowledge of landscape processes, we show that it is possible to gain greater insight into past human and animal behaviour. The approaches we advocate are particularly concerned with placing the hominins (and humans) within the broader faunal community and, in turn, locating the community itself into a very much wider landscape than in traditional approaches. It requires the use of types of data sources and techniques that are commonly used in Earth Sciences but are less commonly employed in questions regarding human evolution and past land use (such as GIS data, fieldwork and satellite imagery).
We highlight and expand upon selected aspects of the reconstruction of ancient land use under the following topics. First, we consider the role of topography when reconstructing the habitats of early hominins in the past. Second, we consider the role of topography and edaphics in constraining the seasonal movements of large herbivores between areas of suitable grazing (Owen-Smith, 1988). We show how humans may have exploited these predictable animal movements for hunting purposes. We present two case studies as illustrations. In the first case, we discuss the example of landscapes around hominin sites in Africa. In the second case, we explore how humans could have exploited megafaunal movements in the southern Levant.
2 Topographic studies in Africa and potential insights into human evolution
It is widely accepted that a progressive, stepwise drying of the climate resulted in the gradual disappearance of tropical forest, starting in the Miocene and becoming well-established by 5 Ma. It affected the range of forest species in substantial parts of Africa, and specifically in East Africa (Bobe, 2006; Cerling et al., 1997; deMenocal, 2004). As forest cover reduced, fauna began to exploit more open savannah conditions characterised by grasslands, smaller tree and shrub vegetation types. Savannah supports today a larger fauna biomass per square kilometre than any other zone of the planet, but forest contains a larger diversity of species (Fig. 1, Maglio and Cook, 1978). Several faunal lineages show radiations as species began to exploit the new ecological niches associated with the increasing grassland (Bobe and Eck, 2001; Bobe and Behrensmeyer, 2004). This is also true for hominins; the fossil record indicates several species of early hominin genera (Sahelanthropus, Orrorin, Kenyanthropus Ardipithecus, Australopithecus), which may reflect different evolutionary strategies to living in new habitat types. Of these genera, only Homo has remained, becoming with time the top predator we know today (e.g., Wood, 2005).
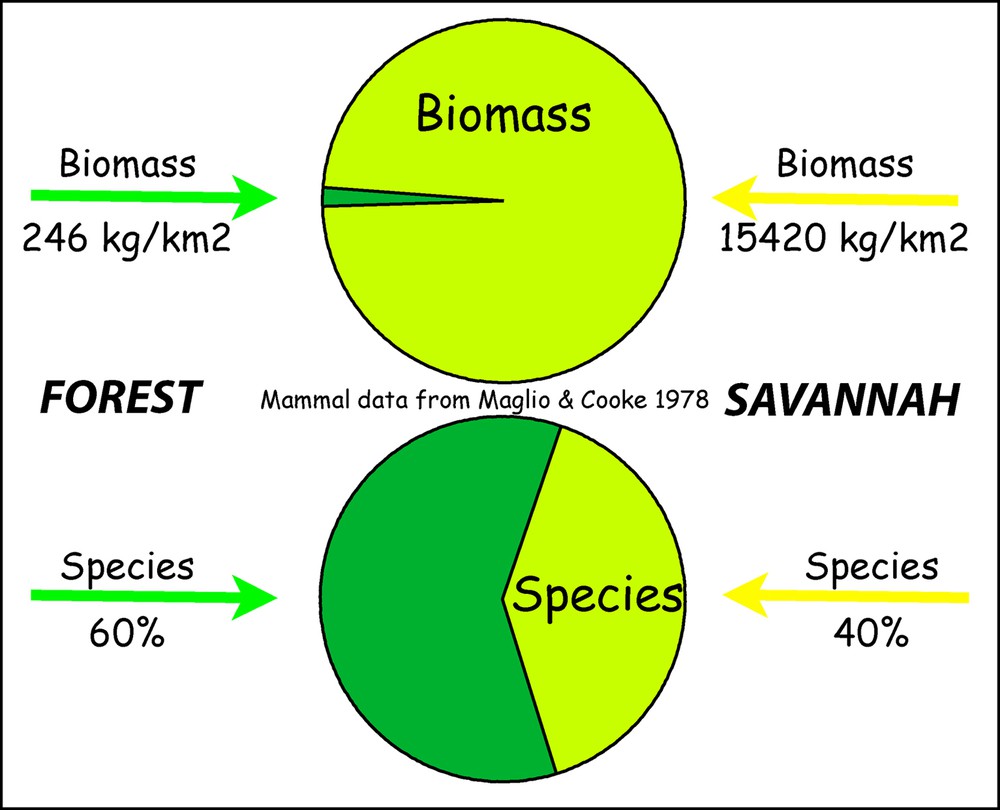
(Colour online). Mammal carrying capacity of forest and savannah (Maglio and Cook, 1978).
The key to how hominins may have adapted to living in novel, challenging environments may well lie in the types of landscapes they exploited. Here we refer to landscapes as the physical landscape Earth scientists are used to exploring (i.e. landform, rock and soils composition, etc.), and we refer to environments as the combination of vegetation and landscapes together. Within this context, “savannah” corresponds more to a vegetation biome as part of the environment, rather than to the physical landscape. While vegetation types are quite sensitive to climate change, features of the physical landscape are not affected to the same extent. Landscapes evolve in response to geodynamical processes such as active tectonics, volcanism, erosion and deposition processes. These processes shape landscape geomorphology, act on the geology and on soil composition, which can in turn affect the “nutrient potential” of a given area. The simple picture one might have of “savannah” becomes more complex when one starts considering the features of the underlying physical landscape. The physical landscape underlying the “savannah” can be made of various types of rocks and soils. It can be rough or flat, covered by volcanics or actively deformed by active tectonics. As a result, it can be more or less sensitive to climatic oscillations.
Habitats are the suitable environmental (vegetation and landscape) conditions that pertain to a particular species (Vrba, 1992). Suitable habitat regions need to meet the key habitat requirements of hominins, namely a range of forage (C3 browse and C4 grass food types), refuge from predation and a source of potable drinking water (e.g., Reynolds et al., 2011). There are specific geomorphological contexts that can provide these habitat requirements, in particular those affected by active tectonic activity.
2.1 Complex topography creates habitats that are suitable for hominins
King, Bailey and collaborators have shown that the locations of Palaeolithic sites are closely associated with topographically complex landscapes (Bailey et al., 2011; King and Bailey, 2006). Fig. 2 illustrates this for Lower Palaeolithic hominin sites. In the majority of cases, tectonic and/or volcanic activity was ongoing when hominins exploited them. These areas are associated with a wide variety of landforms (e.g., rocky outcrops, cliffs, gorges, ridges), hence offering plenty of look-out and observation points, shelter, protection on ledges from most predators and key strategic advantages for hunting (Bailey and King, 2011; King and Bailey, 2010) and others. Hominins, not being very fast, would have been more vulnerable in flat, featureless landscapes – for the same reason, throughout history, persecuted people have always moved to mountainous regions to escape cavalry, trucks or tanks.

(Colour online). Roughness (definition in Bailey et al., 2011) adjusted for altitude and latitude. Roughness is progressively de-emphasised above 1000 metres and at latitudes greater than 40°. The latter approximates inter-glacial conditions. The dots indicate the location of lower Palaeolithic sites outside Africa (see Winder et al., submitted).
Tectonic activity is particularly efficient at creating complex landscapes. Fig. 3A shows the effects of faulting on a riverine landscape: 1) cliffs and river gorges, which hominins and primates could exploit for safety and sleeping roosts, 2) catchment of drinking water and 3) mosaic heterogeneous habitats, possibly associated with a range of food types. The majority of stable isotope studies on enamel from hominin teeth suggest that they consumed mixed diets, with contributions from sedges, reeds and fruits from trees, or animals that fed on these items (Sponheimer and Lee-Thorp, 1999). A key advantage of tectonically active areas is that faulting or folding activity can disrupt water tables and create lakes and swamps that are constantly renewed as long as the activity persists. Faulting can thus provide a range of uplifted, dry areas where grazing animals would predominate, with down-dropped, wetter areas that would essentially provide habitat heterogeneity within a relatively small spatial range that would fit the existing knowledge about hominin habitats from many sites in Africa (see Reynolds et al., 2011 for review).

(Colour online). A. Normal and reverse faulting transforms landscapes. It creates a complex topography associated with features such as cliffs, marshy areas (in the presence of surface water, as shown here) and a range of wetter and drier vegetation zones for foraging. These key landscape features would be attractive to early hominins in Africa, such as Australopithecus africanus (after Bailey et al., 2011, discussed in Reynolds et al., 2011). B. The site of the Makapansgat site and its geomorphology illustrates the range of key landscape features created by faulting activity on the faults, as discussed in A. Masquer
(Colour online). A. Normal and reverse faulting transforms landscapes. It creates a complex topography associated with features such as cliffs, marshy areas (in the presence of surface water, as shown here) and a range of wetter and drier vegetation zones for ... Lire la suite
The landscapes around hominin sites in South Africa, such as Makapansgat and Sterkfontein share similar topographic features. Both sites were inhabited by Australopithecus africanus, a non-tool using hominin that exhibits generalised diets (Sponheimer and Lee-Thorp, 1999; Van der Merwe et al., 2003). Both sites preserve palaeoenvironmental evidence of heterogeneous habitats (also referred to as ‘mosaic habitats’) and swamps. The fossil species indicate high levels of biodiversity around the sites (Vrba, 1982; Reynolds and Kibii, 2011). The presence of aquatic and wetland species also suggest that swamps were present in the past, which could be related to active tectonics in these areas at the time when the sediments were being deposited (Reynolds et al., 2011).
Makapan Valley (South Africa) contains a series of fossiliferous deposits in caves along the valley (Fig. 3A). The Limeworks Cave is a site that contains fossils of Australopithecus africanus, dating back to approximately 3.0 Ma. It is a good case study to illustrate how the landscape features would have affected the fauna and environments present at the site when the species Australopithecus africanus was inhabiting this region. Fig. 3B shows the geomorphology of the Makapan Valley region and the cave sites, with the present-day tectonically-controlled Nylsvlei wetland. The landscape evidence from Makapansgat shows the combination of features, such as the river and the sedimentary plains, which would have offered foraging areas close to the relative safety of the river gorges, caves, and the characteristic cliffs of the Makapan Valley. Within the site region, there are also plains of varying sizes and wetlands within ranging distance of the valley itself. Presence of species such as the klipspringer (Oreotragus oreotragus) as well as several primate species indicates the presence of rocky outcrops and trees preferred by these species (Reynolds et al., 2011 and references therein). There is a close agreement between present habitat heterogeneous conditions and those inferred from the fossil fauna of Limeworks Cave at 3.0 Ma (Reynolds et al., 2011 and references therein). The high biodiversity of present and past species suggests that the wetland and rocky cliff areas were present during the time of hominin occupation, while studies of stable isotope of australopithecines from Makapansgat Limeworks Cave reveal a significant C4 (grass and sedge) signal, suggesting these vegetation types were within a reasonably accessible distance (Sponheimer and Lee-Thorp, 1999).
Similar landscape geomorphology and tectonic activity is identified at the fossil sites of Taung and the Sterkfontein Valley (South Africa). It suggests that tectonic motions created attractive, heterogeneous habitats, which Australopithecus most likely exploited preferentially (Bailey et al., 2011; Reynolds et al., 2011). It is very likely that sites repeatedly occupied over longer timescales (such as Sterkfontein) were tectonically-altered, and that this maintained the suite of attractive features. These landscapes are associated with high levels of habitat heterogeneity and less sensitive to climate changes than flat areas.
Although tectonic activity is the most effective at creating complex landscapes, other processes can have similar effects. In the Arabian desert, the small scale complexity associated with an old inherited relief, and more recent cemented sand dunes, retained palaeolakes under wetter climatic conditions (Petraglia et al., 2012). These would have been associated with transient savannah conditions. The landscape complexity in southern Africa around the “Cradle of Humankind” does not result from tectonic activity in the classic sense either. In this case, it appears to reflect an interaction between the complex geology of the Johannesburg Dome and headwater erosion caused by the Limpopo river system and its tributaries responding to the significant uplift of southern Africa by almost 1000 metres (see Fig. 9 in Bailey and King, 2011). Glacial erosion can also shape a complex landscape, although it is only relevant to the later periods in human evolution. Coastal processes associated with sea level changes can also create complex landscapes (e.g., Bailey and King, 2011). The coast of Saudi Arabia is a good example where hundreds of square kilometers have been exposed at lower sea stand while salt tectonics created rapidly evolving hills and depressions, dramatically impacting the river network (Devès et al., 2011). However, none of these processes affects the critical relationship between the land surface and the water table. They are therefore less likely to create environments where hominins might have a strategic advantage relative to predators and prey species throughout climatic oscillations.
2.2 From trees to cliffs: the Scrambler Man Hypothesis
Early hominins, such as Australopithecus, are considered to have exploited trees for security and progressively used the ‘savannah’ by means of anatomical adaptations progressively acquiring effective bipedalism. The energetically (relatively) efficient Homo body form appeared about 2 Ma ago (Wood, 1992). The most widely accepted view is that adaptation allowed more rapid movement on flat ground, known as the Endurance Running Hypothesis (Bramble and Leiberman, 2004) and freed the arms for carrying. Other possibilities such as thermoregulatory efficiency have also been proposed (Wheeler, 1993). Winder et al. (2013) propose an alternative hypothesis that considers the role of topography to be central to the evolution of the early Homo body form.
This “Scrambler Man” hypothesis would have favoured adaptations by allowing a progressive transition to the upright posture through climbing and scrambling activities. This view challenges previous models based on adaptations to forest or savannah (Fig. 4) in favour of physical incentives presented by steep rugged terrain. Pursuing his prey across rugged topography, “Scrambler man” could have acquired greater speed and agility over time. This hypothesis explains well the key anatomical changes observed over the course of human evolution (e.g., Fig. 7 of Winder et al., 2013).

(Colour online). Cartoon showing the evolution of hominin locomotor capabilities as predicted by the traditional “savannah” hypothesis (left) and the “complex topography hypothesis” (right). Both sequences begin with the predicted last common ancestor of Pan and Homo at about 6 Ma towards the top and culminate in Homo sapiens at the bottom (after Winder et al., 2013).
Complex topography has not only played a role in human evolution and hominins land use but also on animals. We will now consider landscape constraints that control animal movements through a given terrain and how humans living in the southern Levant may have exploited these predictable movements for hunting purposes.
3 The role of complex topography and soil edaphics in animal movement and its impact on hominins land use: the example of the southern Levant
The physical attributes of a landscape can affect animal movements. Our second case study concerns a combination of two such factors: 1) the roughness of the topography itself, and 2) the soil characteristics that indicate areas of good and poor pasture quality.
The majority of herbivores migrate depending on changes in available water, grazing and seasonal temperature variations. These are described as seasonal movements when there is little change in latitude. In modern landscapes, seasonal movements of wild animals are limited by humans via the fencing off of large areas. Historic documents record these types of migrations. Early European explorers to southern Africa recorded mass migrations of springbok (Antidorcas marsupialis) and other animals to areas of fresh grazing (Skinner and Louw, 1996). Seasonal resource variations are still exploited by transhumant shepherds who move their animals in the same way that species such as the red deer (Cervus elaphus) would have moved in the past without human intervention. Green (1998) has identified the routes followed by shepherds for northwestern Greece and identified where the migrating herds were intercepted by cattle rustlers. These are similar to places identified in a palaeoanthropological project where early humans intercepted red deer migrations, by means of strategically chosen bottleneck locations that split the herds off into smaller groups (Bailey et al., 1993).
3.1 The role of soils composition on animal migration
Soil attributes, referred to as edaphic factors, can affect animals movements by enhancing the “nutritional” qualities of certain vegetation. Edaphic factors, or the shorthand term “edaphics”, concern the ability of the regolith (i.e. soils and subsoils) to supply, by plant take-up, the nutrients necessary for herbivore growth, health and reproduction. These edaphic factors are critical to the growth of young animals and the continued health of all the animals. The availability of soluble phosphates is especially critical, as this supports bone development. Simple phosphorus or phosphate levels in soils and subsoils are not, on their own, guides to the edaphic quality of the soils. Tricalcium di-orthophosphate, the main constituent of animal bones, is only soluble in ionised acidic water, and its release into soils in a form which can be taken up by plants is therefore normally very slow. So for high edaphic quality in relation to phosphates, not only is an adequate source of the minerals required, but the conditions must be such that the minerals can actually be taken up by the fodder which the herbivores eat (Henkin et al., 1995). Even where the regolith provides abundant sources of the main minerals and nutrients in a form which can be taken up by plants, specific trace elements may be missing, for example, elements, such as selenium, cobalt, copper or potassium (Burrows et al., 1979; Corah, 1996; Formigoni et al., 2011; Kadim et al., 2003; Ruess, 1984). In such cases, animals may need to make occasional movements towards sources of fodder supplying them. Nowdays, fodder supplement and fertiliser providers can handle this requirement without the need to move domestic animals.
It is important to distinguish between “fertility”, in the sense of plant growth, and “edaphics”. A given regolith might support abundant vegetation, but this may be of poor quality when viewed as a food and trace element source for herbivores. Over wide areas of the Mediterranean, for example, until the recent substantial fall in grazing pressures resulting from changes in human economics, the edaphically poorer areas often carried more abundant plant vegetation that was less exploited by animals, while the vegetation in the edaphically richer areas was bitten down hard, reflecting their differential attractiveness to herbivores (Sturdy and Webley, 1988; Sturdy et al., 1997).
3.2 Locating sites with respect to megafaunal movements in the southern Levant
We have discussed hominin use of complex topography to gain a strategic advantage relative to other animals. Individual hominins lack the characteristic features of major predators, such as speed, strong teeth, and large size. Cursorial predators rarely hunt large animals such as elephant, rhinoceros, and for the species of large ungulates such as buffalo and wildebeest, predation is typically restricted to young, old or weakened individuals. Yet Homo appears to have exploited adults of large species in large numbers (Grimstead, 2010; Shipman et al., 1981).
In the Levant, fossils of very large fauna fossils are found in several Palaeolithic sites indicating that they were eaten by hominins (Owen-Smith, 1988 defines megafauna to be over 1000 kg in adult body weight but notes that other authors go as low as 45 kg – in his book on mega fauna, he includes Asian rhino less than 1000 kg). In the following, we propose to explore the way in which edaphic factors may have constrained large herbivores seasonal migration in addition to landscape complexity. We then compare the location of some key sites with respect to the routes that we can infer from this approach.
If animals are predated they will seek to avoid places where it has occurred in the past and “attempt” to be unpredictable. A modern analogy is the manner in which diplomats in countries where there is a danger of terrorist attack actively avoid following regular routes. However, this is not always possible for diplomats or for the species that hominins preyed on. Migration hunting depends on positioning the available hominins where animals are due to pass. If the route varies, this strategy fails. An example of the cost of this strategy failing is the short-lived and unsuccessful attempts of Barren Ground Eskimo to adopt this approach after the introduction of guns to the Canadian North (Hurst, 2004; Mowat, 1952). The animals chose alternative routes.
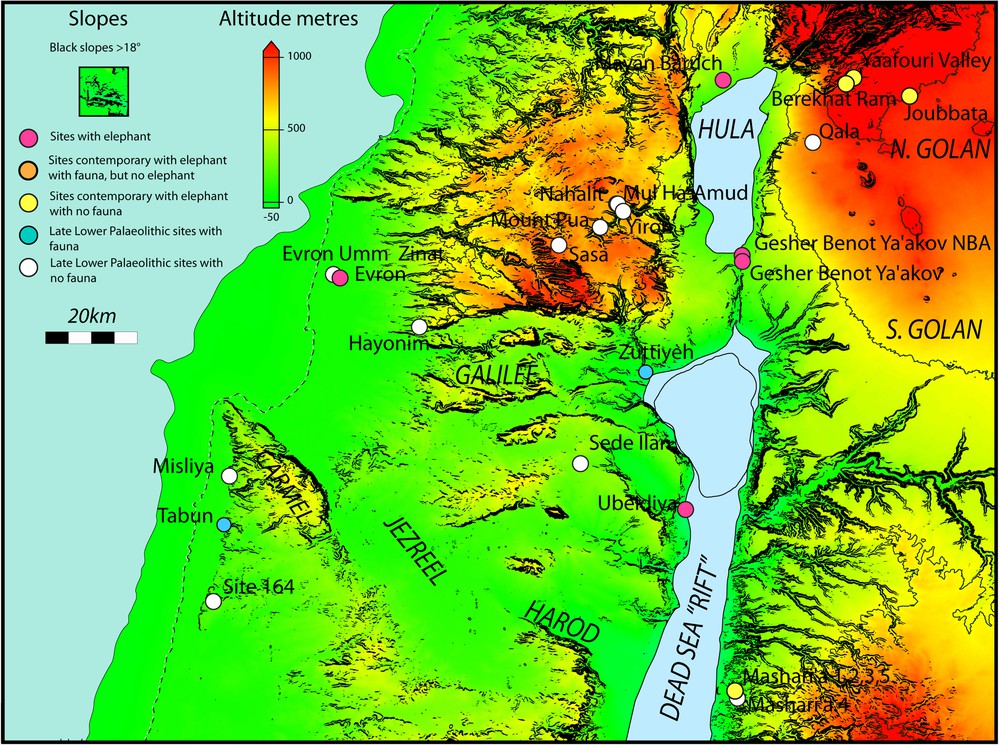
Lower Palaeolithic sites in the Levant, dating back to between about 1.5 Ma and 0.3 Ma, are shown in Fig. 5. We focus on the rectangular area in the figure, which is particularly rich in sites exhibiting evidence for Homo erectus exploiting megafauna including elephant species. We refer to various species of elephant but it should be understood that similar environmental constraints applied to other very large herbivores.

Significant Lower Palaeolithic sites in the southern Levant, plotted on to a relief map. The sea levels shown are the present-day, −50 m (orange contour) and −100 m (red contour). Lake Lisan (proto Dead Sea) is shown at its maximum extent (light blue) and the current extent (darker blue). This can be taken to approximate high levels of Lake Amora, a general name for fluctuating lakes that have filled the valley over the last ∼ 1 Ma. Changing lake levels illustrate the sensitivity to climate changes. Note that the coastal plain becomes very narrow for all sea levels in what is now Lebanon restricting access to the north. Briefly sea level could have been a few metres higher than at present. Masquer
Significant Lower Palaeolithic sites in the southern Levant, plotted on to a relief map. The sea levels shown are the present-day, −50 m (orange contour) and −100 m (red contour). Lake Lisan (proto Dead Sea) is shown at its maximum extent (light ... Lire la suite
3.2.1 Landscape characterisation
Fig. 6A shows the topography of the region as a simple map of altitude. There is an altitude range from east to west of nearly 1000 metres across the region. Except for very brief periods when it was ∼ 10 metres higher) the sea level was below the present and the coastal plain would have been wider. The region is broken up by steep slopes with black shading showing slopes steeper than 18° (about 30%) impassable to large animals. The complexity of the topography results from tectonic activity associated with the Jordan (Dead Sea) valley fault system (Devès et al., 2011). The fault is mainly strike-slip and hence imposes mostly horizontal displacement. The overall shape of the Jordan valley is created by modest vertical displacement (relative to the more pronounced horizontal displacement) and it is likely that the depth of the Jordan valley has not changed greatly over the last 1.5 Ma (Matmon et al., 1999, 2000a,b, 2003).
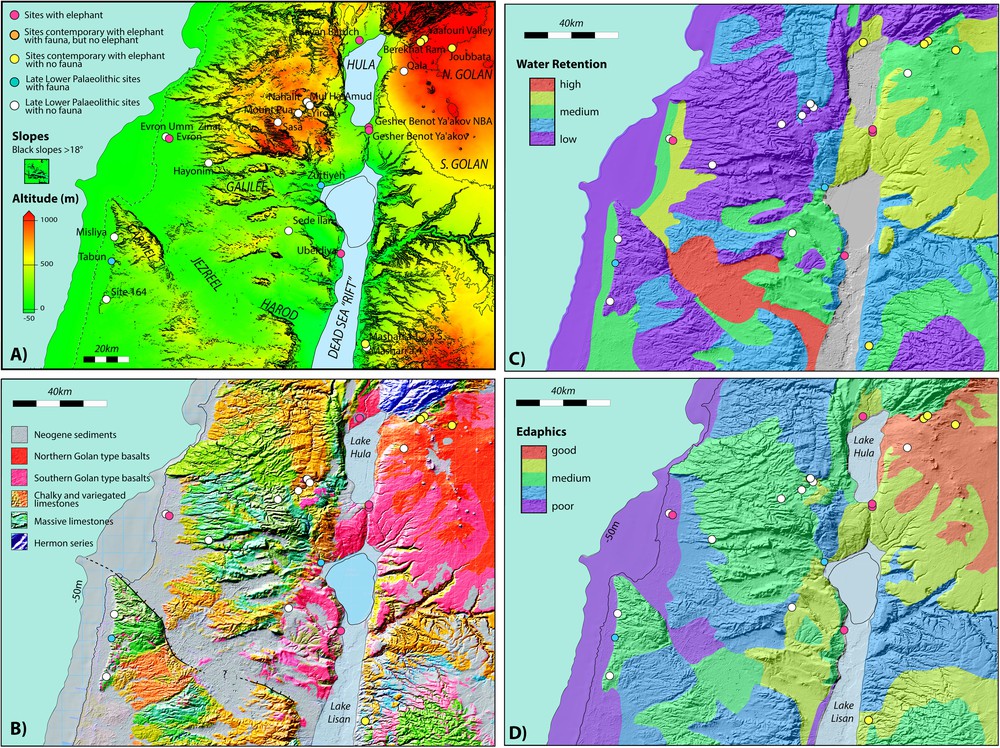
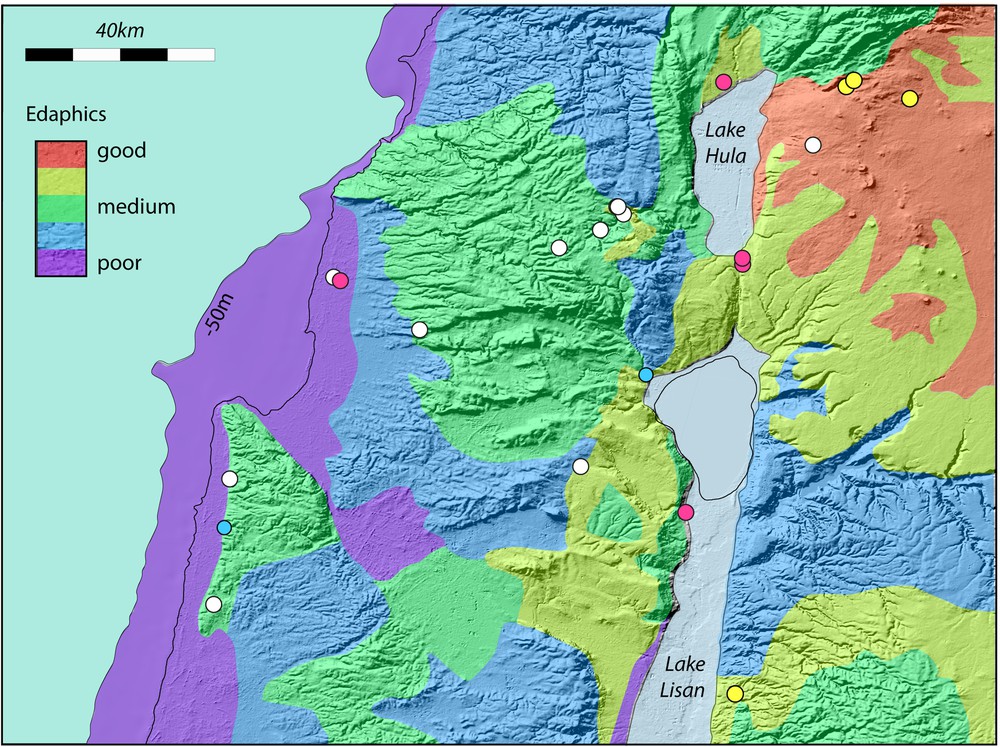
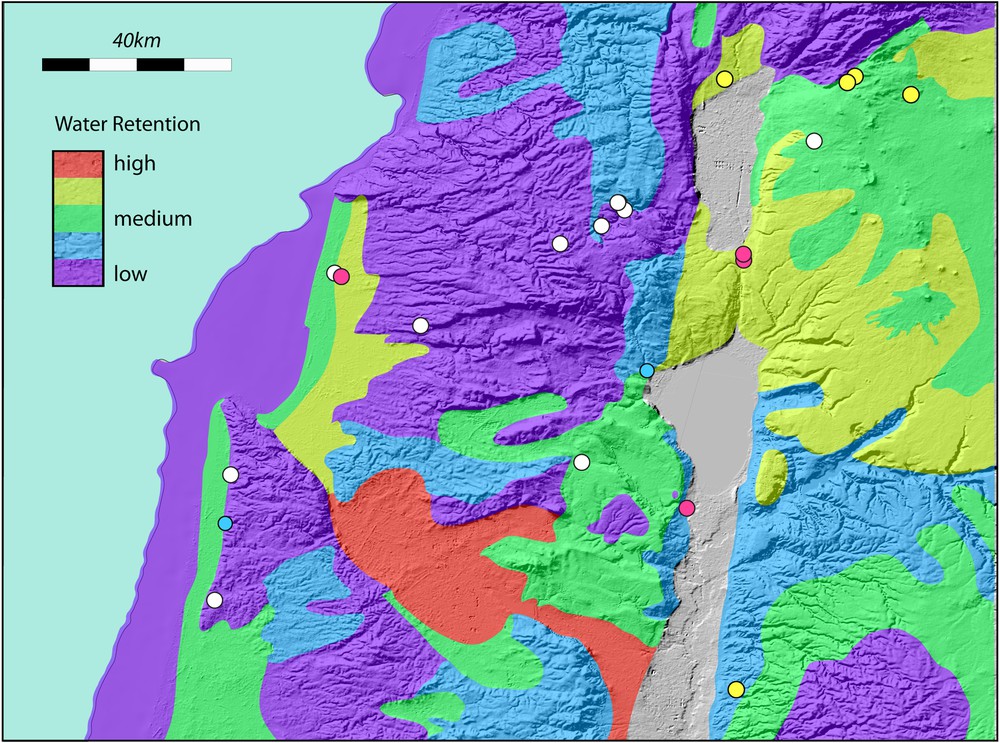
A. The Carmel, Galilee and Golan region showing significant Lower Palaeolithic sites. The current coastline (dashed) and the coastline at −50 m are shown. Lake Lisan is shown at its maximum extent. This can be taken to approximate high levels of Lake Amora. The current Sea of Galilee is outlined. Site colours as in Fig. 5. Slopes greater than 18° (∼ 30%) are indicated by black shading. B. Simplified geological map of the Carmel, Galilee and Golan regions, plotted on a relief map. Only the Haifa and Gilboa faults are shown. It is the modest, but ongoing motion on these faults that maintains marshy conditions in the Jezreel and Harod valleys. The morphology of the region is dominated by faulting that has occurred over the last 5 Ma and longer. More information is provided by references in the text. The map is modified from the Geological Map of Israel. 1:200,000. State of Israel, Ministry of National Infrastructures, Geological Survey of Israel. Jerusalem 1998. C. Interpreted principal edaphic characteristics of the Carmel, Galilee and Golan regions, plotted on a relief map. Edaphic characteristics are based on the geology. D. Simplified map of relative water retentiveness of soils in the Carmel, Galilee and Golan regions. Typical soil thickness as well as the water retentiveness characteristics were used to construct the figure. Masquer
A. The Carmel, Galilee and Golan region showing significant Lower Palaeolithic sites. The current coastline (dashed) and the coastline at −50 m are shown. Lake Lisan is shown at its maximum extent. This can be taken to approximate high levels of Lake ... Lire la suite
Rivers may have cut deeply into the valleys adding to the steep topographic features generated by tectonics. It is clear however that the topography of the region has not changed substantially over the time period of interest here (Matmon et al., 1999, 2000a,b, 2003). With the exception of small-scale features (less than about 1 km) and some volcanic features in the Golan, most features would have been similar to what they are today. More importantly, the places where the landscape is complex today should represent well the places where the landscape was complex at that time. On this basis it is reasonable to use the present landforms to represent the earlier environment. The present coastal plain has dunes and marshes and small outcrops of limestone areas sub-parallel to the coast and it is likely that the wider plain at lower sea levels was similarly endowed. By restricting animal movements these may have provided opportunities for hominin exploitation. They are not however major features and we will focus the following discussion on the important complexity of the inland region.
The geology of the region is shown in Fig. 6B. Combining fieldwork, tectonic and geology cartography (Devès and King) with the experience gained by colleagues (Webley and Sturdy) in other parts of the Mediterranean, it is possible to outline important features of soils, including the degree to which water can be retained, and the quality of the soils. We propose that the soils composition and the associated vegetation may have controlled the movements of the large herbivorous mammals (megaherbivores) during the Lower Palaeolithic period. Our aim here is to demonstrate that important information can be extracted by adopting a broad landscape approach (a detailed study has been done in Devès et al., 2014).
3.2.2 Possible seasonal migration routes
Large animals such as elephants move in groups and consume large volumes of vegetation. The elephant (Loxodonta africana) consumes, for instance, at least 200 kg of vegetation daily and for this reason alone must keep moving (Estes, 1991). At first sight, it seems curious that very large herbivores could find sufficient range and fodder in the relatively confined areas of the southern Levant. The fossil evidence provides however prima facie evidence of their presence in the area. It is not possible to know the exact distribution and exact evolution of vegetation and vegetation types throughout the period of interest. It would have changed with climatic oscillations. Today's landscape can however, under given assumptions, provide a good proxy to what the landscape was “on average” in the past. As we discussed above, the morphology of the landscape does not appear to have changed dramatically since the times of occupation by hominins. Erosion and deposition rates are small. It is hence reasonable to consider that the soil composition can be deduced from the lithology (geology) to a first order at least, representative of the overlying soils composition. One can also argue that whatever the vegetation cover is, and has been, places with good edaphics are better for browsing and grazing than places with bad edaphics. We also have the chance to deal with a region where there is nearly a 1000-meter altitude difference between the low and the high lands. It seems reasonable to assume that the latter has played, not the same, but a similar role throughout climatic variations. Although the higher areas enjoy higher precipitation, this can turn to snow, producing winter conditions, which animals such as the elephants and rhinoceros of our period could probably not endure. Today, winters in the northern Golan and in the Upper Galilee are certainly harsh enough to make them unattractive to many of the herbivores we are considering. In the wetter and cooler phases of the Pleistocene, this situation would have been accentuated.
Our assessment of the edaphic potential of the region is shown in Fig. 6D. It is based on the underlying geology, assessments in similar Mediterranean regions (Sturdy et al., 1997), descriptions of Israeli soil types (Shapiro, 2006) and our own field observations. In general, beach or coastal sedimentary deposits resulting from lowered sea levels develop poor soils. Other sedimentary deposits can be good depending on the geology of their source rocks. Chalks are poor and limestones are often moderately good. Basalts are good in general and some basalt soils in the northern Golan have been known to be excellent since biblical times (This is reflected in the reluctance of the cattle-raising tribe of Reuben to cross the Jordan, mentioned in the biblical accounts of the Israelite “conquests” of Canaan (Numbers 32: 1 ff, Joshua 1: 12 ff), and the famed size and strength of the “Bulls of Basan” from the southern Golan). Even within regions of poor soils, windblown material or the disintegrating corpses of dead animals can improve the quality of local soils. However, while this can be exploited by smaller, solitary herbivores, these small-scale improvements will not support, nor be attractive to, large animals moving in abundant herds.
The ability of soils to retain water is an additional important aspect for supporting herbivores. During dry periods, soils that cannot retain water become useless while water-retentive soils can be exploited over much longer time periods. Fig. 6C shows a map of water retentiveness. Soils that develop on land revealed by falling sea levels do not usually retain water. Limestones that develop very thin soils are also poor during dry seasons. Fine-grained sediments from streams, rivers or slope-wash can be water retentive. Land that has at times been marshy and contains a high proportion of organic material can also retain water over long periods of time. Basalts can erode to form pockets of water-retaining soil.
Considering the different landscape and soil properties summarised above (in Fig. 6) one can suggest possible routes for large animals, in particular elephants. This is synthesized in Fig. 7. Starting with the spring season, the animals would have moved eastwards from the coastal plain towards the more edaphically productive areas to take advantage of the best nutrition of the year. They would follow the growth on such limestone areas as were available (for example, the easier ground south of Carmel between the coastal plain and the Jezreel valley), eastwards towards the more phosphate-rich basaltic soils. Henkin et al. (2007) stress that the first flush of new forage is the most nutritious, and here altitudinal differences would give large herbivores an incentive to follow the new forage at higher altitudes as it became available. This enables us to visualise the route the animals would have followed. The low-lying basalt soils of the eastern Galilee would be the location of the earliest occurrences of nutritious grazing at the beginning of the season, after which animals would have moved on to the Golan, crossing the Jordan valley where they could. The highest point of the southern Golan (close to the Acheulean sites of Berekhat Ram and its neighbors) represents the limit of the new forage. After reaching the northern Golan, much of the summer would be spent in making use of the available grazing east of the Jordan River. At times in the Pleistocene, recent lava flows would limit access to certain specific areas, while the lower nutritional value of the southern Golan, compared to its northern part, would be offset by the availability of fodder through the summer.

(Colour online). Reconstructed seasonal movements of large herbivores in the Middle Pleistocene of the Carmel, Galilee and Golan regions.
With the late summer/dry-season approaching, the animals would move back towards the limited resource of the Harod-Jezreel areas, where water-retentive soils, and marshlands provided important dry-season fodder. The winter rains would then make the coastal plain at low altitudes the obvious main grazing area for winter.
However, navigating through the complex topography (Fig. 7), in particular the steep slopes, would have limited the possible routes that animals would have taken between these areas of seasonal grazing.
Based on these reconstructed herbivore movements across the landscape, one can reinterpret the location of some important Lower Palaeolithic sites. Three of the sites with direct evidence of megaherbivore fossils, namely Gesher Benot Ya’aqov, Ma’ayan Baruch and ‘Ubeidiya can be positioned in relation to megafaunal preferential routes. At Gesher Benot Ya’aqov, for instance, crossing the Jordan valley has never been easy and, particularly so, at times when the Jordan valley was fully occupied by a lake as far north as Lake Kinneret. The site is beside where road crosses today and where a crusader fort (destroyed by Saladin) was built to control the strategic route from the coastal towns to Damascus. Ma’ayan Baruch is located in an area of good edaphics, with some variable topography and close to Lake Hula. Ubeidiya was at the edge of a lake. However the key to the site seems to be the region to the west: a sedimented basin with a rim of basalt, which hosted rich and water-retentive soils that would have offered attractive grazing for animals. Sites such as Berekhat Ram, Yaafouri valley and Joubbata were contemporary with the “elephant” sites and in a region of excellent edaphics. However, the area has been subsequently covered by more recent volcanic activity, which limits the interpretations we can make. The site of Evron Quarry appears nowadays difficult to assess but was probably in a region accessible to megaherbivores that may have been associated with the coastal plain features mentioned previously. Several Late Lower Palaeolithic sites can be seen in Fig. 7, but these are clearly located in regions that would have been inaccessible to elephant and other megaherbivores. Perhaps this indicates the start of a transition to predating smaller animals in the Middle Palaeolithic as the megafauna disappeared. Similar behavioural shifts from large game hunting to trapping and snaring smaller animals have been noted in other contexts in the Middle Stone Age in South Africa (Wadley, 2010).
4 Conclusions
Fossils and archaeological remains are embedded in landscapes that evolve dynamically due to various types of geomorphological processes. By understanding the processes controlling the evolution of the landscapes in the Levant and Africa, we were able to suggest how early hominins and humans could ensure their survival by exploiting specific landscape characteristics.
Habitats in complex topography offers several attractive features: relative shelter from predation, strategic advantages for hunting large animals, a variety of food resources and access to drinking water even under unfavorable climatic conditions. Hominins could use their strategic advantage in complex terrain to predate other animals, even if they lacked the speed or power of other predators, such as lions and hyaenas. A number of factors required prey species to move predictably. We examined the role of soil edaphics in the southern Levant. Trace elements in soils create rich grazing and browsing in areas that herbivorous animals must periodically visit. These features allow reasonable reconstructions of the seasonal movements of prey animals. The positions of key fossils sites appear to lie on these routes, suggesting that hominins were aware of the patterns of animal movements and found ways to exploit this knowledge of migration routes.
The role of the complex topography and soil factors as a means of controlling prey seasonal migration and favoring hunting activities has received almost no attention in the palaeoanthropological literature, but can provide, as shown here, fascinating insights into past human landscape use.
Landscapes form an important component of habitats used by animals and humans. Our understanding of the ways that animals and humans used their landscapes in the past could be considerably furthered by studying the features of the physical landscape itself and would undoubtedly benefit from the knowledge and techniques of Earth sciences.
Acknowledgments
The authors thank Geoffrey Bailey. Many of the ideas presented here come from our joint work. The project is supported by the European Research Council (ERC) through Advanced Grant 269586 ‘DISPERSE: Dynamic Landscapes, Coastal Environments and Human Dispersals’ under the ‘Ideas-specific’ Programme of the 7th Framework Programme (FP7). We appreciate the help provided by Amotz Agnon, Kate Raphael, Shai Shir and other Israeli colleagues. Nan Godet has received financial aid from the Southern Levant Human Environment Project. This is IPGP contribution 3618 and DISPERSE contribution 22.
Appendix A Supplementary data