

 CC-BY 4.0
CC-BY 4.0
1. Introduction
The Jurassic of Southern Vendée is a thin platform series which lies unconformably on the Paleozoic Armorican basement, of which it represents the southern cover (Figure 1). Its geographical position, near the Atlantic Coastline, makes it the westernmost sedimentary witness of the northern Aquitaine border. Thus, the associated ammonite faunas, are the westernmost representatives of the Western European faunal ensemble. The Pliensbachian deposits are privileged paleogeographical witnesses because of their proximity to the North-Western margin of the Bay of Biscay.
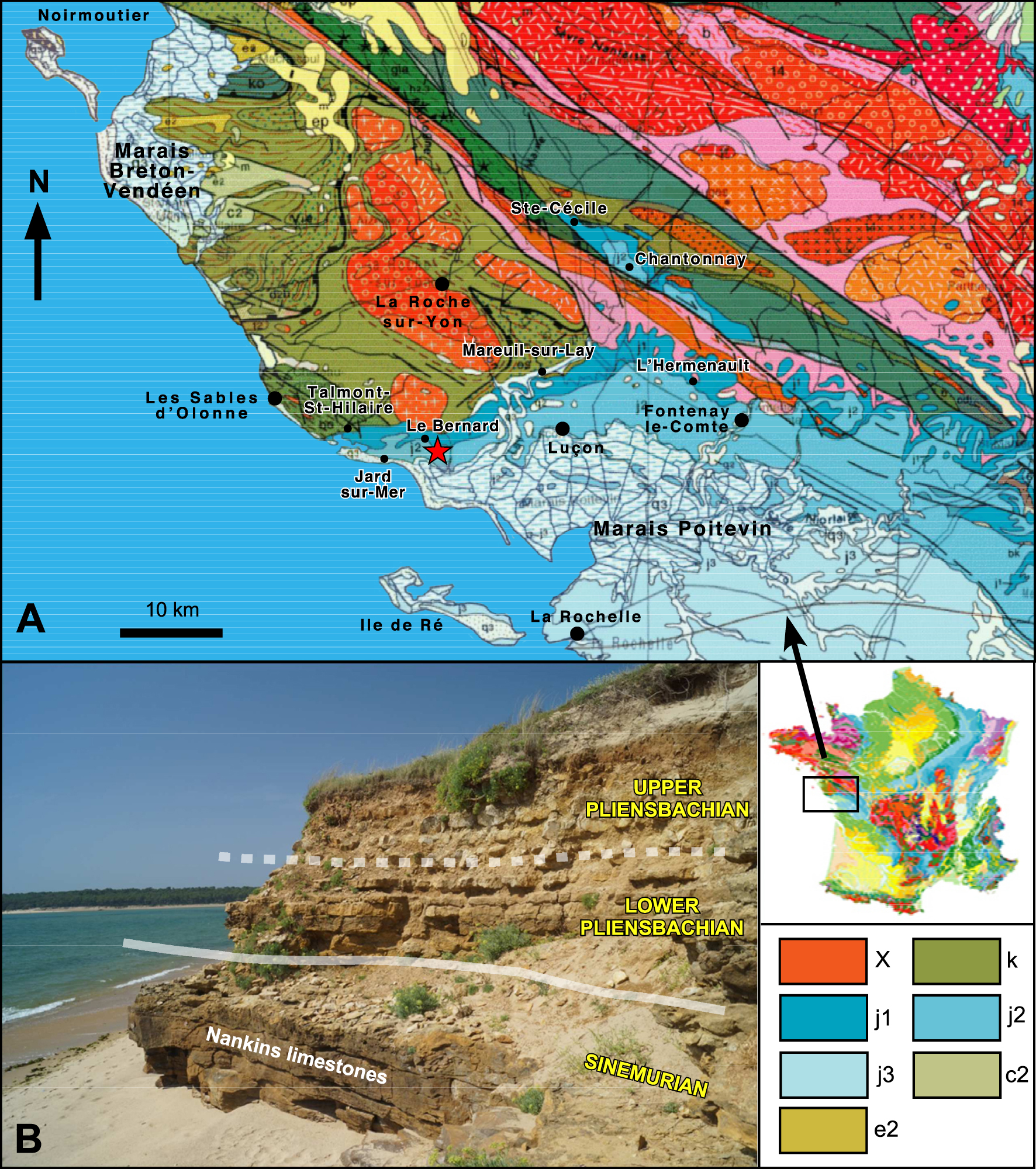
(A) Geographical and geological setting. Location of Southern Vendée. Simplified geological map (BRGM editions) and localities cited in the text. X: Metamorphic; k: Paleozoic; j1: Lower Jurassic (Lias); j2: Middle Jurassic; j3: Upper Jurassic; c2: Cretaceous; e2: Paleocene. Red star: Location of the Le Bernard section. (B) Photography of the Upper part of the Sinemurian “Nankins Limestones” and of the Pliensbachian clayed limestones and marls of the Payré Estuary (Jard-sur-Mer).
Located north of the Iberian plate, the Bay of Biscay is an essential structural element of the Iberia–Armorica–Newfoundland conjugate margin. The Rift of Biscay was set up from the Triassic or the Lower Jurassic [Olivet 1996; Rasmussen et al. 1998], at the same time as a mosaic of pre-drift basins which prefigured the North Atlantic Sea at the west of Iberia microplate (Figures 2, 8). In the absence of outcrop, we know nothing of the fauna of these essentially offshore basins, particularly of their macrofauna, and we have no element of comparison with the Jurassic of Vendée.
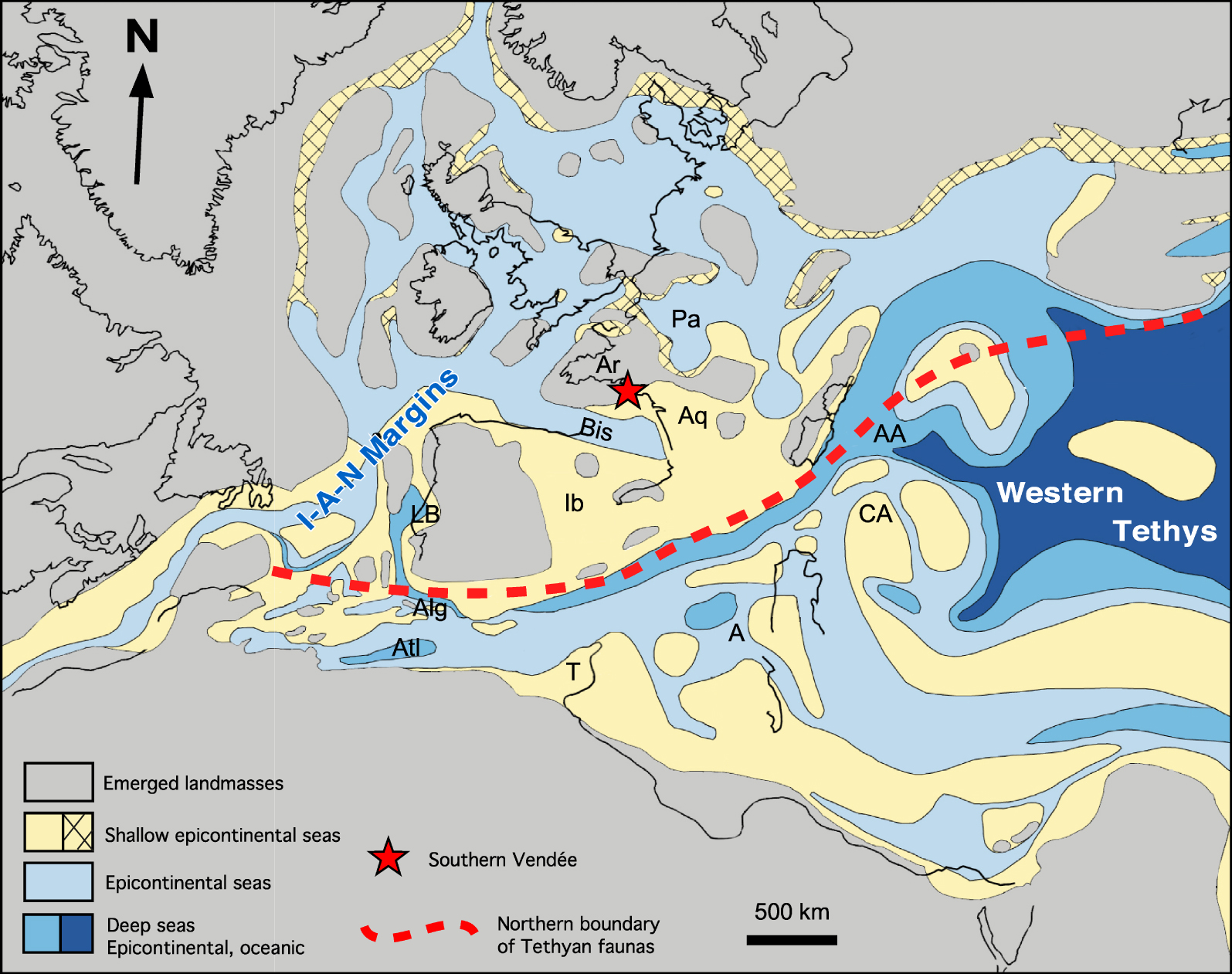
Paleogeographical framework for the Pliensbachian at the scale of Western Tethys [modified from Thierry et al. 2000]. Abbreviations. I-A-N Margin: Iberian-Armorica-Newfoundland Margins; Ar: Armorican Massif; Aq: Aquitaine Basin; Bis: Biscay Gulf; Pa: Parisian Basin; Ib: Iberian Basin; LB: Lusitanian Basin; Alg: Algarve; Atl: High Atlas; T: Tunisia; A: Apennines; AA: Austroalpine; CA: Calcareous Alps.
The Lusitanian Basin, located on the western Iberian margin (Figures 2, 8), is the only one to present rich Lower Jurassic onshore outcrops, whose ammonite faunas are now well known [Mouterde et al. 1983, 2007; Dommergues et al. 2010].
Belonging to the North-West European Type (Figure 2), these faunas are, on several occasions in the Upper Sinemurian, then in the Lower Pliensbachian, enriched by highly endemic faunas (genera, species). This remarkable endemism, although punctual and short, suggests a particular paleogeographical configuration of the conjugate “Iberia–Armorica–Newfoundland” passive margin of the Atlantic passive margin [Dommergues et al. 2010]. To date, its causes remain still poorly understood. It probably finds its origin in the relative isolation of the pre-existing Permian–Triassic west Iberian rift domain [Nirrengarten et al. 2018; Angrand et al. 2020]. Also, it has probably been favored by the early Jurassic first phase of rifting that created narrow and deep offshore basins, typically corresponding to asymmetric half grabens generated by tilted blocks tectonics (e.g. Lusitanian, Porto, Porcupine, Rockall Basins) [Olivet 1996; Rasmussen et al. 1998] (Figures 2, 8). Despite the lack of data about macrofauna in these offshore basins, it was very likely that all were interconnected, and that the Portuguese endemism was not restricted to the Lusitanian Basin alone, but could affect others neighbouring basins of “Iberia–Armoric–Newfoundland” [Dommergues et al. 2010].
The stratigraphic and taxonomic revision of the Pliensbachian ammonite faunas of Southern Vendée, that we have undertaken provides an element of response, by highlighting the presence, among the ammonites of the Vendean Lower Pliensbachian, of several taxa related to the Lusitanian endemism, of which the genus Dayiceras is the most emblematic element. The connection with the Lusitanian faunas is reinforced, in the Upper Pliensbachian, by the presence, in Vendée, of ammonites of Tethyan (Mediterranean) origin, abnormally numerous in North-Western Europe, but identical to those present in abundance at the same time, in the Lusitanian Basin.
This study presents the summary of a taxonomic revision of the Pliensbachian ammonite faunas of Southern Vendée based on material collected in situ, put in a rigorous biostratigraphic framework considering all paleobiogeographical and paleoecological aspects [Fauré and Bohain 2017; Bohain and Fauré 2022]. It offers a reliable spatio-temporal framework that we compare here with paleogeographic interpretations in the context of the pre-drift basins of the “Iberia–Armorica–Newfoundland” conjugate margins.
2. Stratigraphy
The Pliensbachian of Southern Vendée is a thin (4.5 m) condensed clayed limestone platform series, which overlies the “Sinémuro-Hettangian” dolomitic limestones called “Nankins Limestones”, or overlaps directly the Paleozoic Armorican basement (Figure 1A). From Bourgenay (Talmond-Saint-Hilaire) to Saint-Nicolas Cove (Jard-sur-Mer), the layers of Pliensbachian age are known for their emblematic coastal outcrops, which can be followed for several kilometres of cliff or foreshore (Figure 1B). The hills of the Vendean hinterland (regions of Luçon, Chantonnay, Sainte-Cécile, and L’Hermenault) are much unfavourable for their study because of the rather flat topography where outcrops are rare and ephemeral (Figure 1A).
Although it has been already described by Péneau [1923], Butel [1951, 1953], Gabilly [1964], and Dubar and Gabilly [1964], the Vendean Pliensbachian remained little studied until recent stratigraphic revisions by Alméras et al. [2010a, b], based on the study of brachiopods, and those of Fauré and Bohain [2017] and Bohain and Fauré [2022], on ammonites, of which we summarize here some of the results.
The Pliensbachian sedimentary sequence is made of detrital silico-arkosic sandstones or microconglomerates (less than 0.5 m thick), followed by an alternation of clayed limestone and bioclastic marl beds rich in ferruginous oolites (3 to 4 m thick), particularly in the coastal part of Vendée where sedimentation rate decreases. The richness of these layers in fossils, such as brachiopods and ammonites, as well as corals, bivalves, gastropods, and echinoids, has proven to be remarkable and the specimens are particularly well preserved. Despite the low rate of sedimentation, the condensation (2 m thickness for the Lower Pliensbachian; 2.50 m thickness for the Upper Pliensbachian), and the disturbance caused by hydrodynamics and bioturbation, the succession of the ammonites has been found in conformity. We have been able to identify [Fauré and Bohain 2017; Bohain and Fauré 2022] all the standard chronostratigraphic divisions of the North-West European Pliensbachian [Dommergues et al. 1997; Page 2003] (Figures 3, 4, 5).
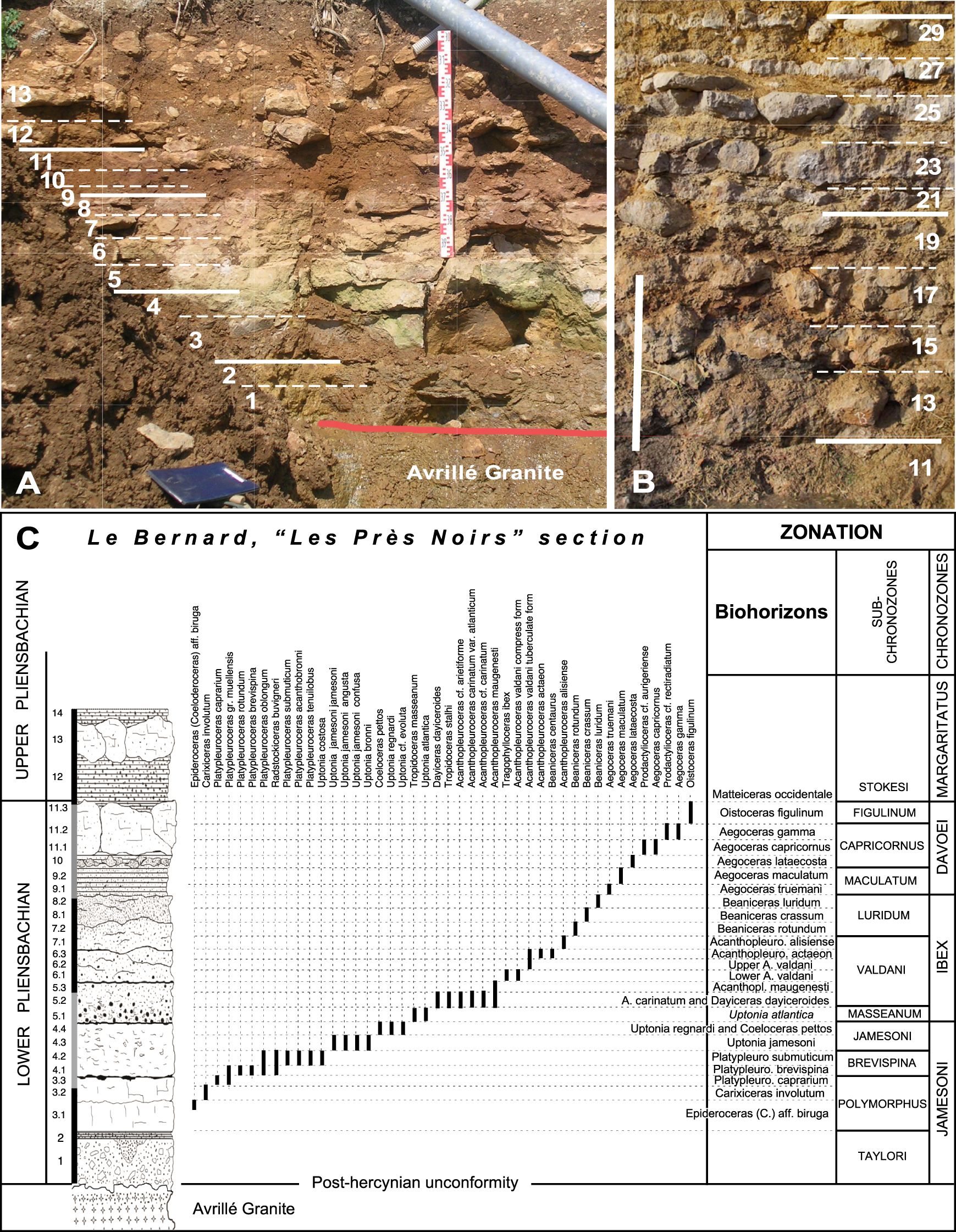
Le Bernard, “Les Prés Noirs” section. (A) Photography of the Lower Pliensbachian (scale: 1 m); (B) Photography of the Upper Pliensbachian (scale: 1 m); (C) Lithologic succession of the Lower Pliensbachian distribution of main ammonites (out of 87 taxa listed in this section. Complete inventory in Fauré and Bohain 2017, Figure 9) and chronostratigraphic framework.
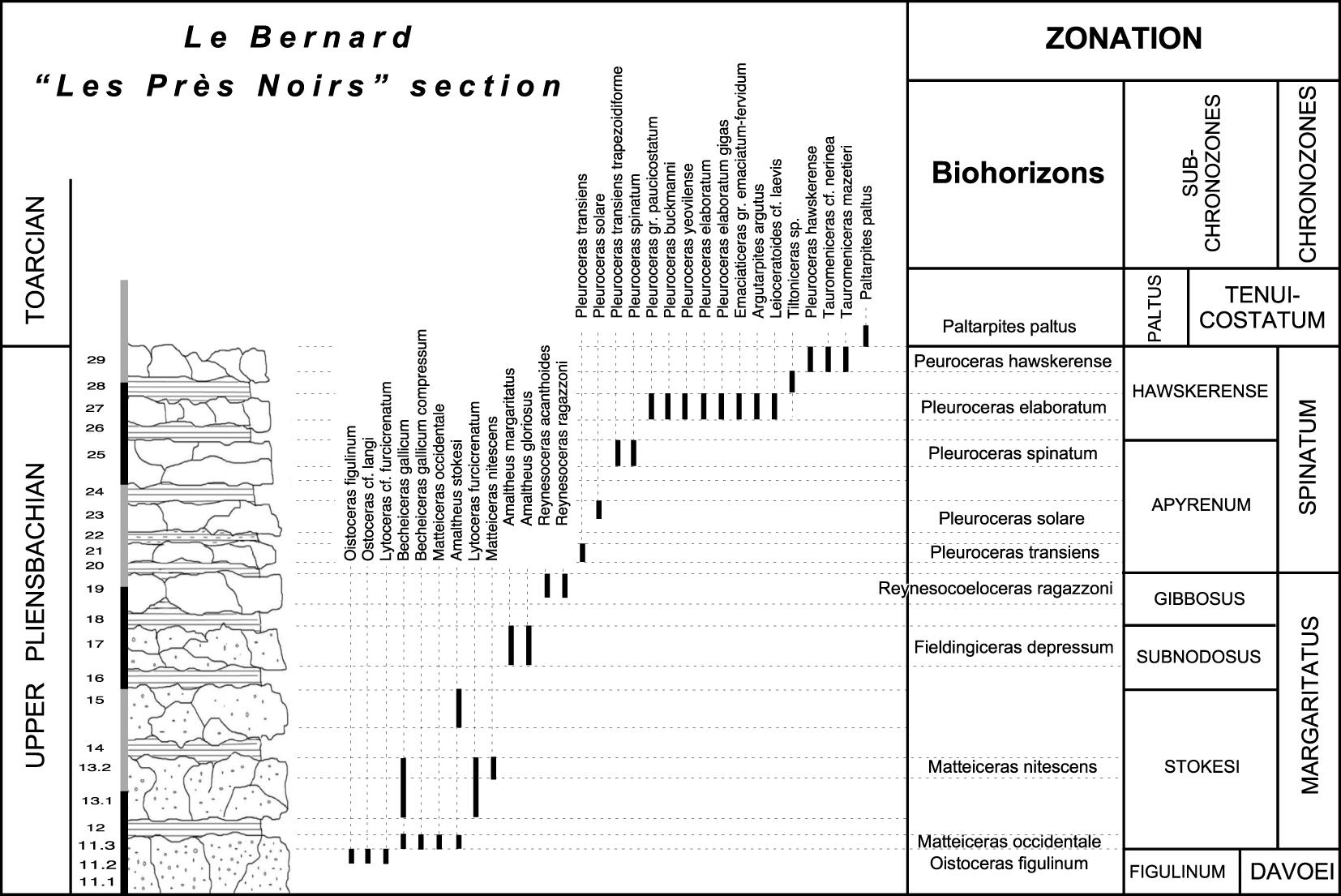
The Upper Pliensbachian of the Le Bernard, “Les Près Noirs” section. Lithologic succession, distribution of ammonites and chronostratigraphic framework.
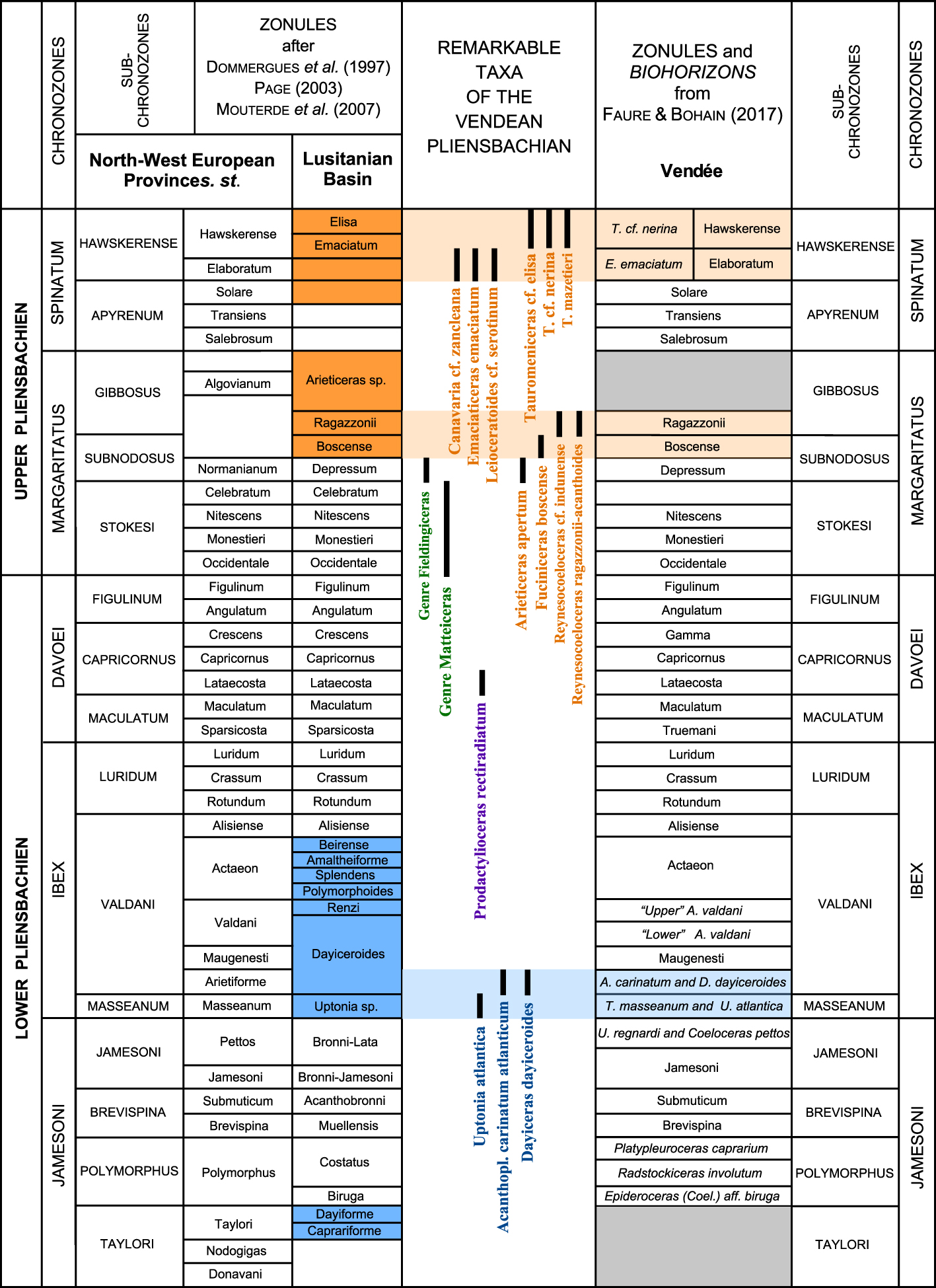
Ammonite standard zonation for the North-West European Province and the Lusitanian Basin [Dommergues et al. 1997; Page 2003; Mouterde et al. 2007]. Comparison with the zonule and biohorizon (in italic) succession adopted in Southern Vendée [Fauré and Bohain 2017]. In dark blue: periods of endemism of the Lusitanian faunas; in light blue: period of extension of Lusitanian endemism to the Vendée region; in dark orange: periods of expansion of Tethyan taxa in the Lusitanian Basin; in light orange: periods of Tethyan taxa presence in the Vendée Upper Pliensbachian; in grey: absence of fauna. Remarkable taxa column. Blue: species related to the Lusitanian endemism which are present in Vendée; purple: species common to Portugal, Vendée and Dorset; green: North-West European genera of “southern” affinity; orange: Tethyan taxa. Masquer
Ammonite standard zonation for the North-West European Province and the Lusitanian Basin [Dommergues et al. 1997; Page 2003; Mouterde et al. 2007]. Comparison with the zonule and biohorizon (in italic) succession adopted in Southern Vendée [Fauré and Bohain Lire la suite
3. Chronostratigraphic marks
The biostratigraphic precision offered by several sections of the Armorican occidental border, of which the Le Bernard, “Les Prés Noirs”, section is the main one, allowed us to determine certain points of the ammonite succession. In the Lower Pliensbachian, from the Polymorphus Subchronozone to the Figulinum Subchronozone, 24 distinct chronostratigraphic divisions, standard zonules, or local biohorizons, could be separated [Fauré and Bohain 2017] (Figures 3A, C). In the Upper Pliensbachian, 14 divisions could be separated thanks to Tethyan Hildoceratidae and Dactylioceratidae which offer the opportunity of a stratigraphic fineness that is not allowed by Amaltheidae alone [Bohain and Fauré 2022] (Figures 3B, 4).
3.1. Lower Pliensbachian (Figure 3C)
3.1.1. Jamesoni Chronozone
- Polymorphus Subchronozone: It has been divided into three distinct units, Epideroceras (Coeloderoceras) aff. biruga biohorizon, Carixicerasinvolutum biohorizon, and Platypleurocerascaprarium biohorizon; when the standard biozonation for North-Western Europe distinguishes only one (Polymorphus Zonule), and the Lusitanian standard two (Biruga and Costatus Zonules) [Page 2003].
- Jamesoni Subchronozone: Its division into two units is modeled on the evolutionary sequence of the genus Uptonia, with the two successive species Uptonia gr. jamesoni [Sowerby 1812–1822] and Uptonia regnardi [Orbigny d’ 1842–1849] (Figure 6A):
- Jamesoni Zonule: Its limits are based on a new interpretation of the species Uptonia jamesoni and U. lata [Quenstedt 1845–1849], which we have already shown that they correspond, not to chronologically successive species, but to synonymous species [Fauré and Téodori 2019].
- Uptonia regnardi and Coeloceras pettos biohorizon: The association in Vendée of the two taxa Uptonia regnardi [Orbigny d’ 1842–1849] and Coeloceras pettos [Quenstedt 1845–1849] allows us to integrate the Pettos Zonule of the standard divisions into the Jamesoni Chronozone (Figure 5).

A1–A2, Uptonia regnardi [Orbigny d’ 1842–1849], Jamesoni Chronozone and Subchronozone, Uptonia regnardi and Coeloceras pettos biohorizon, Le Bernard, niv. 4.4, Neotype designed by Fauré and Bohain [2017], X78; B, C, D, Uptonia atlantica Fauré and Bohain 2017, Ibex Chronozone, Masseanum Subchronozone, Tropidoceras masseanum and Uptonia atlantica biohorizon, Le Bernard, niv. 5.1: B1–B3, Holotype, Y55; C1–3, Paratype, V70; D1–2, Z126; E, F, G, Uptonia atlantica Fauré and Bohain 2017, Jamesoni Chronozone, Masseanum Subchronozone, Peniche (Portugal): E, V73; F1–2, V72; G1–3, V75; H, I, J, K, Dayiceras dayiceroides [Mouterde 1951], Ibex Chronozone, Valdani Subchronozone, Acanthopleuroceras carinatum and Dayiceras dayiceroides biohorizon, Le Bernard, niv. 5.2: H1–2, Y58; I1–3, Z121; J1–2, Z122; K1–2, AU7; L1–3, Dayiceras dayiceroides [Mouterde 1951], Ibex Chronozone, Valdani Subchronozone, Dayiceroides Zonule, Peniche (Portugal): V84. The material belongs to the P. Bohain collection. It is kept in the collections of the Natural History Museum of Nantes (France). Scale bar 1 cm (all samples are natural size). Masquer
A1–A2, Uptonia regnardi [Orbigny d’ 1842–1849], Jamesoni Chronozone and Subchronozone, Uptonia regnardi and Coeloceras pettos biohorizon, Le Bernard, niv. 4.4, Neotype designed by Fauré and Bohain [2017], X78; B, C, D, Uptonia atlantica Fauré ... Lire la suite
3.1.2. Ibex Chronozone
- Masseanum Subchronozone: It corresponds to the acme of the species Tropidoceras gr. masseanum [Orbigny d’ 1842–1849]. In Vendée, it also contains the late Uptonia, U. atlantica Fauré and Bohain 2017, allowing us to individualize Tropidoceras masseanum and Uptonia atlantica biohorizons (Figures 3, 5).
- Valdani Subchronozone: It could be divided into six successive units (Figure 3C):
- Acanthopleuroceras carinatum and Dayiceras dayiceroides biohorizons, equivalent to the Arietiforme Zonule of the standard; which combines primitive Acanthopleuroceras (A. arietiforme [Oppel 1853], A. carinatum [Quenstedt 1885], A. carinatum atlanticum Dommergues and Mouterde 1981) and the late Polymorphitidae Dayiceras dayiceroides [Mouterde 1951].
- Maugenesti Zonule. Dayiceras is not seen in Vendée these days, while the genus remains abundant in the Lusitanian Basin.
- Valdani Zonule. It is subdivided into two distinct biohorizons based on the predominance of two morphotypes of the species Acanthopleuroceras valdani [Orbigny d’ 1842–1849], which were however not distinguished at a specific level [Fauré and Bohain 2017].
- Actaeon Zonule and Alisiense Zonule, without local particularity.
3.1.3. Davoei Chronozone
- Maculatum Subchronozone: The two standard zonules are based on the evolution of primitive forms of the genus Aegoceras (Figures 3C, 5):
- Truemani Zonule. We substituted Aegoceras truemani Fauré and Bohain 2017 (Figure 5) for the standard index ammonite “Aegoceras sparsicosta” [Trueman 1919], because we consider the latter taxon as the peramorphic form (“Androgynoceras” form) of a Beaniceras from the Luridum Subchronozone.
- Maculatum Zonule. Without modification.
- Capricornus Subchronozone:
- Lataecosta Zonule. In Vendée, it is the level of Prodactylioceras rectiradiatum [Wingrave 1916].
- Gamma Zonule. We have substituted Aegoceras gamma Dommergues 1979 (Figure 5) for the standard index ammonites “Aegoceras crescens” (Hyatt 1867 in Trueman 1919), a species whose generic attribution (Aegoceras or Oistoceras) was ambiguous.
- Figulinum Subchronozone: Vendée data did not suggest any modification. It is limited at its top by the first concomitant appearance of the Amaltheidae [Amaltheus stokesi Sowerby 1812–1822] and Harpoceratinae (genus Matteiceras).
3.2. Upper Pliensbachian (Figures 3B, 4)
3.2.1. Margaritatus Chronozone
- Stokesi Subchronozone: Primitives Amaltheus [Amaltheus stokesi, A. bifurcus [Howarth 1957–1958], A. wertheri Lange 1932] are associated with Harpoceratinae which are grouped in the subgenus, or genus, Matteiceras [Wiedenmayer 1980] whose succession makes it possible to recognize, in Vendée, three standard zonules (Occidentale, Monestieri and Nitescens Zonules) for Southern Europe (Figures 4, 5).
- Subnodosus Subchronozone: It is the interval of existence of the group of Amaltheusmargaritatus Montfort de 1808, A. gloriosus [Hyatt 1867] and A. subnodosus [Young and Bird 1828]. The Harpoceratinae, grouped in the genus Fieldingiceras [Wiedenmayer 1980], makes it possible to identify the Depressum Zonule in which appears the first Arieticeratinae of Tethyan affinity, Arieticeras apertum [Monestier 1934] (Figure 4).
- Gibbosus Subchronozone: Amaltheus gibbosus [Schlotheim 1820] is associated with A. margaritatus. Two standard zonules defined in Southern Europe [Page 2003] and based on taxa of Tethyan origin, are identified (Figures 4, 5):
- Boscense Zonule, with Fuciniceras boscense [Reynès 1868].
- Ragazzonii Zonule, with the brief expansion of the genus Reynesocoeloceras [R. gr. acanthoides [Reynès 1868]—ragazzonii [Hauer 1856] and R. gr. indunense Meneghini 1867–1881], two Dactylioceratidae of Tethyan origin, which are a good time mark for the lower subchronozone. These taxa are present, at an identical level, in Portugal [Mouterde et al. 2007] and in Grands-Causses [Meister 1989].
An important discontinuity marks a gap in the sedimentary record that lasts during the Middle et Upper part of the Gibbosus Subchronozone (Figure 5).
3.2.2. Spinatum Chronozone
- Apyrenum Subchronozone: The index Pleuroceras apyrenum [Buckman 1909–1930] is rare, so P. solare [Phillips 1829] appears a better indicator of the subchronozone. The three standard Salebrosum, Transiens and Solare zonules are present (Figure 5). The last one contains P. spinatum [Bruguière 1789] and Amaltheus margaritatus which persists up to the top of the zonule. Hildoceratidae of Tethyan origin are missing in Vendée.
- Hawskerense Subchronozone: The Pleuroceras are still very predominant, but the subchronozone is well marked by two waves of Arieticeratinae and Harpoceratinae belonging to Tethyan genera or species.
- Elaboratum Zonule, with the index species Pleuroceras elaboratum [Simpson 1884], P. gigas Howarth 1957–1958, P. buckmani [Moxon 1841], P. yeovilense Howarth 1957–1958 and P. paucicostatum Howarth 1957–1958. This is the level in Vendée of the Mediterranean taxa Emaciaticeras emaciatum [Catullo 1853]—fervidum Fucini 1931 (Figure 7B), E. cf. archimedis Fucini 1931 (Figure 7F), Canavaria cf. zancleana [Fucini 1931], Leioceratoides cf. serotinum [Bettoni 1900] (Figure 7D) and L. cf. laevis [Haas 1913] (Figure 7E).
- Hawskerense Zonule, with Pleuroceras hawskerense [Young and Bird 1828] and the Tethyan taxa Tauromeniceras cf. elisa [Fucini 1931] (Figure 7C), T. cf. nerina [Fucini 1931] and T. mazetieri [Dubar 1927] (Figure 7A).

A1–A2, Tauromeniceras mazetieri [Dubar 1927], Spinatum Chronozone, Hawskerense Subchronozone and Hawskerense Zonule, Le Bernard, niv. 29: AT9; B, Emaciaticeras gr. emaciatum [Catullo 1853]—fervidum Fucini 1931, Spinatum Chronozone, Hawskerense Subchronozone and Elaboratum Zonule, Talmont-Saint-Hilaire, Coteau des Draillards (=Le Bernard niv. 27): ER2 (×0.75); C1–C2, Tauromeniceras cf. elisa [Fucini 1931], Spinatum Chronozone, Hawskerense Subchronozone and Zonule, Sainte-Cécile, La Maison Neuve: FR7; D, Leioceratoides cf. serotinum [Bettoni 1900], Spinatum Chronozone, Hawskerense Subchronozone and Elaboratum Zonule, Sainte-Cécile, Coteau de Bellevue: EL8; E, Leioceratoides cf. laevis [Haas 1913], Spinatum Chronozone, Hawskerense Subchronozone and Elaboratum Zonule, Talmont-Saint-Hilaire, Bas de la Brunetière (=Le Bernard, niv. 27): EW7; F1–F2, Emaciaticeras cf. archimedis Fucini 1931, Spinatum Chronozone, Hawskerense Subchronozone, Elaboratum Zonule, Sainte-Cécile, Coteau de Bellevue: AG7. Scale bar 1 cm (all samples are natural size, except B). Masquer
A1–A2, Tauromeniceras mazetieri [Dubar 1927], Spinatum Chronozone, Hawskerense Subchronozone and Hawskerense Zonule, Le Bernard, niv. 29: AT9; B, Emaciaticeras gr. emaciatum [Catullo 1853]—fervidum Fucini 1931, Spinatum Chronozone, Hawskerense Subchronozone and Elaboratum Zonule, Talmont-Saint-Hilaire, ... Lire la suite
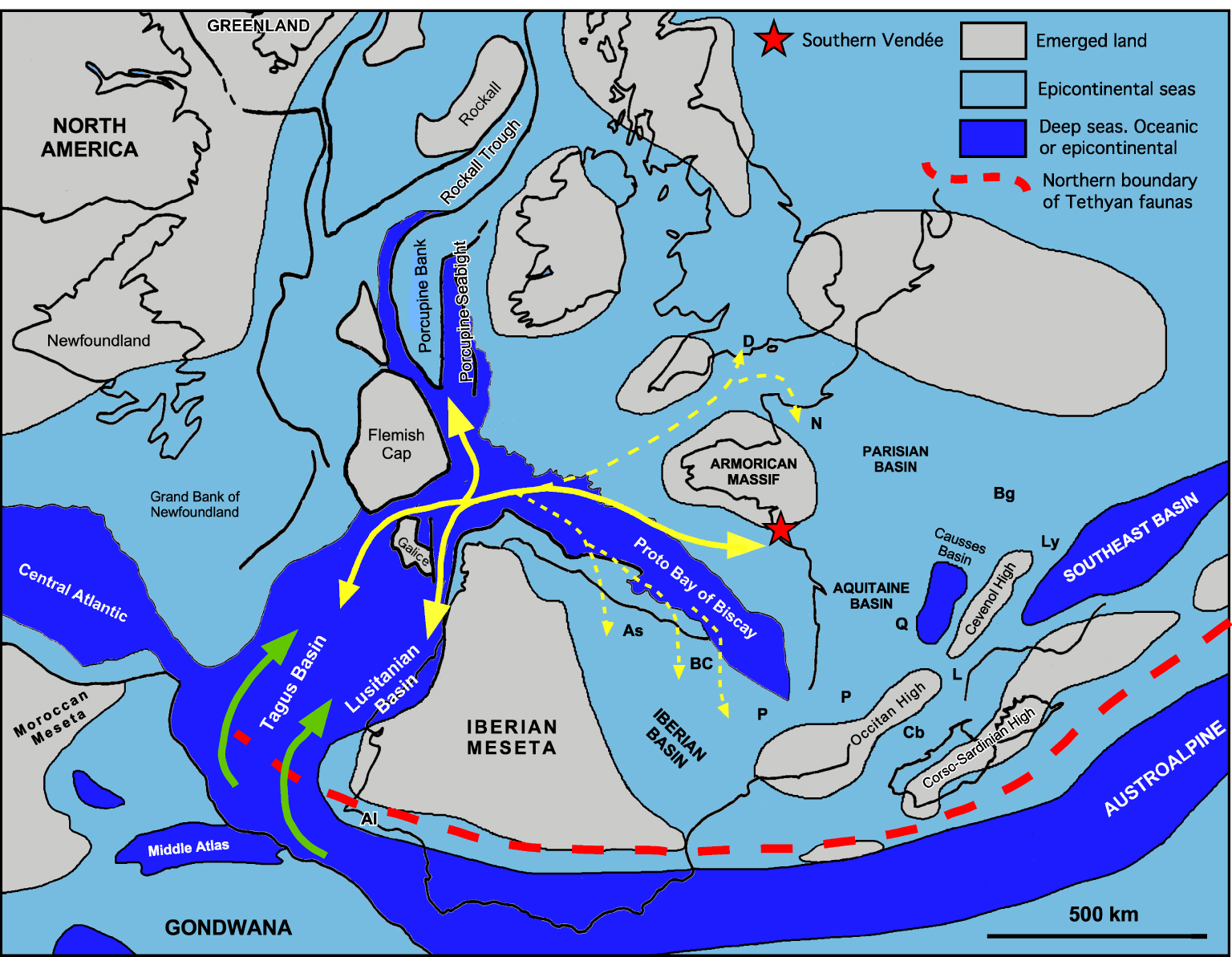
Paleogeographical reconstruction of the Atlantic region and western Tethys for the Jurassic and relative movements of Iberia and Europe. Example of partial closure of peri-Iberian oceanic basins with limited strike-slip movement between Europe and Iberia, according to Olivet [1996, Figure 12] under the assumption of basins inherited from either the Paleozoic or the Triassic. Green arrows: migration route of Upper Pliensbachian Tethyan taxa from the Mediterranean realm to the Atlantic area. Continuous yellow arrows: Lower Pliensbachian. Hypothetical main exchange route of Uptonia atlantica and Dayiceras dayiceroides between an Atlantic source of evolution, the Lusitanian Basin, and Southern Vendée (red star). Upper Pliensbachien: Same hypothetical migration route of Tethyan taxa inside the Atlantic area. Dotted yellow arrows: Lower and Upper Pliensbachian. Hypothetical way of dispersal of isolated individuals to the northern European platforms. Abbreviations. Al: Algarve; As: Asturias; BC: Basco-Cantabrian Basin; P: Pyrenees; Cb: Corbières; L: Languedoc; Q: Quercy; Ly: Lyon Area; Bg: Burgundy; D: Dorset; N: Normandy. Masquer
Paleogeographical reconstruction of the Atlantic region and western Tethys for the Jurassic and relative movements of Iberia and Europe. Example of partial closure of peri-Iberian oceanic basins with limited strike-slip movement between Europe and Iberia, according to Olivet [1996, ... Lire la suite
We demonstrate here that the standard Elaboratum and Hawskerense zonules of the Northwestern European standard are respectively equivalent to the Emaciatum and Elisa zonules defined in Portugal, a region in which amaltheids are scarce [Page 2003; Mouterde et al. 2007] (Figure 5).
4. Paleobiogeographic affinities of the Ammonites
The Pliensbachian Ammonitina under study are clearly related to the North-West European Paleobiogeographic Bioprovince (Figure 2) and the standard biozonation can be applied without amendment at the level of the chronozone and the subchronozone (Figure 5). The presence, in the Lower Pliensbachian, of taxa representative of the endemism prevailing in the Lusitanian Basin, then in the Upper Pliensbachian, of numerous Tethyan taxa, confers clear originality to the ammonite faunas of Southern Vendée.
4.1. Vendean lower Pliensbachian
In Vendée, the Lusitanian affinity ammonites are numerous within the lower part of the Ibex Chronozone (Masseanum and Valdani Subchronozones); a time-interval during which the endemism is precisely the most marked in the Lusitanian Basin [Phelps 1985; Dommergues 1987] (Figure 5). These taxa are for the first time brought to light outside the Lusitanian Basin:
- Uptonia atlantica Fauré and Bohain 2017 (Figure 6B–G) (Polymorphitidae). It is the “Uptonia sp.” of Mouterde et al. [1983], identified by these authors in the Masseanum Subchronozone of the Peniche section (Portugal) (Figure 5). In Vendée, this well-established species clearly succeeds to U. regnardi and, as in Portugal, extends the monophyletic clade of the genus Uptonia, into the Masseanum Subchronozone (Ibex Chronozone).
- Dayiceras dayiceroides [Mouterde 1951] (Figure 6I–L) (Polymorphitidae). This is the first species belonging to the genus Dayiceras. This species, which still shows many characters of Uptonia, extends into the Valdani Subchronozone the lineage of the Polymorphitidae. In Vendée [Fauré and Bohain 2017], as in Portugal [Mouterde 1951, 1967; Mouterde et al. 1983], it occurs at exactly the same level (Arietiforme Zonule) (Figure 5).
- Acanthopleurocerascarinatumatlanticum Dommergues and Mouterde 1981 (Tropidoceratidae). First Acanthopleuroceras at the base of the Valdani Subchronozone. This micromorphic species is considered by Dommergues and Mouterde [1981] as a geographical subspecies of A. carinatum [Quenstedt 1885], which would have undergone possible constraints specific to the Lusitanian Basin.
While in Western Europe (France, Germany, Spain, England), the Polymorphitidae lineage died out at the top of the Jamesoni Chronozone, in Southern Vendée, as in Portugal, it persists until the Masseanum Subchronozone and the lower part of the Valdani Subchronozone (Ibex Chronozone) with the two species Uptonia atlantica and Dayiceras dayiceroides. These two species were previously only known in the same stratigraphic level in the Peniche and São Pedro de Muel sections (Portugal) where they were considered to be particularly representative of the Lusitanian endemism [Mouterde et al. 1983; Phelps 1985; Dommergues 1987].
D. dayiceroides is however the only Dayiceras represented on the Armorican border whereas, in the Lusitanian Basin, the genus diversifies up to the top of the Actaeon Zonule. For reasons unknown, no ammonite relating to the Lusitanian endemism is recognized in Vendée after the Arietiforme Zonule while the endemic continues, very marked, in Portugal, until the top of the Actaeon Zonule (Figure 5).
4.2. Vendean upper Pliensbachian
Most of the Upper Pliensbachian ammonite assemblages of Southern Vendée are representative of the North-West European paleobiogeographic Province, but the presence of numerous Hildoceratidae (Harpoceratinae, Arieticeratinae) and Dactylioceratidae of Tethyan origin, gives to the Vendean ammonite associations a very “southern” character. We find them, in greater or lesser abundance, at several stratigraphic levels (Figure 5):
During the Stokesi and the Subnodosus subchronozones, Harpoceratinae are represented by the genera, Matteiceras and Fieldingiceras. Although these taxa have a Tethyan origin, they are totally endemic to the North-West European Province. They, nevertheless, predominate in the southern part of the European platform, near the North Tethyan margin [Dommergues and Mouterde 1980; Dommergues 1987; Meister and Stampfli 2000].
Within their maximum distribution area, we will first mention the Lusitanian Basin where they clearly outclass the Amaltheidae [Mouterde et al. 2007]. They are also very frequent in the meridional part of the European craton: Iberian Ranges [Comas-Rengifo 1985], Basque-Cantabrian Basin, Asturias [Rodriguez-Luengo et al. 2012], Pyrenees [Fauré 2002; Fauré and Téodori 2019], Grands-Causses [Meister 1989], Quercy [Fauré and Brunel 2019], Corbières [Fauré 2006], Languedoc and Cevenol Margin [Mattei et al. 1971; Cassel 1997] and Provence [Lanquine 1935]. They also reach the Lyonnais [Rulleau 2007], Burgundy [Dommergues and Mouterde 1980], Lorraine [Maubeuge 1971], as well as England [Howarth 1992] and Southern Germany [Fischer 1975] where these are therefore few specimens, or even isolated individuals.
Their abundance in Vendée, away from the north Tethyan margin, must be emphasized.
During the Gibbosus and Hawskerense subchronozones, the genera Leioceratoides (Harpoceratinae), Arieticeras, Emaciaticeras, Tauromeniceras (Arieticeratinae), Reynesocoeloceras (Dactylioceratidae), and the species they represent, are this time authentically Tethyan taxa, recognized at an identical stratigraphic level in the Mediterranean Province such as the Maghrebian margin of Africa (Morocco, Algeria, Tunisia), the Betic Ranges, or the Apennines.
Their frequency in Southern Vendée, far from the north Tethyan margin, is remarkable. Except for the Lusitanian Basin which is occasionally, at the end of the Spinatum Chronozone, integrated into the Mediterranean Province [Mouterde et al. 2007], these ammonites are still few in the southern part of the North-West European platforms: Basque-Cantabrian Basin, Asturias [Comas-Rengifo et al. 2016], Iberian Ranges [Comas-Rengifo 1985; Braga et al. 1982; Comas-Rengifo et al. 1999], Southern Pyrenees [Fauré 2002], Corbières [Fauré 2006] and Grands-Causses [Meister 1989]. They are notably found in regions further north as Dorset [Howarth 1992] and Normandy [Dubar 1927].
This colonization movement of the South Armorican margin by Tethyan faunas goes on during the Paltus Subchronozone (Lower Toarcian). At that time, many Tethyan affinity ammonites (Protogrammoceras, Eodactylites, Neolioceratoides, …) will constitute, as in Portugal, most of the ammonite associations [Bécaud 2005].
5. Discussion
5.1. The expression of the lower Pliensbachian Lusitanian endemism
It should be remembered that, during the Early Pliensbachian, the Lusitanian Basin was exclusively populated with genera and species of North-West European type. The paleogeographical configuration of this region has, however, on several occasions, been favourable to the development of endemic taxa (species and genera). These phases of endemism begin in Portugal as early as the Upper Sinemurian (Obtusum Chronozone), a time interval in which it only affects the Asteroceratinae Family, with the genera Ptycharietites, Epophioceroides, and Pompeckioceras, whose representatives are only known in Portugal [Dommergues et al. 2010].
- During the Taylori Subchronozone (Jamesoni Chronozone), targeted endemism phenomena affects the Phricodoceratidae with the presence of the endemic genus Pseudophricodoceras [Mouterde et al. 1983]. This genus has however been reported in the “Sierra de la Demanda” (Iberian Cordillera) by Comas-Rengifo et al. [1988] and in North-West Germany by Hoffmann [1982].
- With the Brevispina Subchronozone, the apparent endemism phenomena described by Mouterde et al. [1983] must be put into perspective. Two micromorphic Polymorphitinae, Platypleuroceras muellensis [Mouterde 1951] and P. acanthobronni [Mouterde et al. 1983], said to be endemic to the Lusitanian Bassin, are respectively the microconch of two ammonites, Playpleuroceras gr. brevispina [J. de C. Sowerby, 1823–1846] and P. submuticum group [Oppel 1853], whose paleogeographic range is much wider [Fauré and Téodori 2019].
- With the Valdani Subchronozone (Ibex Chronozone), the endemism is the most pronounced (Figure 3). Indeed, while on the North-West European platform, the Polymorphitidae (genus Uptonia) are completely replaced at the top of the Jamesoni Subchronozone by the Tropidoceratidae (genera Tropidoceras and Acanthopleuroceras), in the Lusitanian Basin, the evolutionary sequence of the Polymorphitidae continues within the Ibex Chronozone with several taxa, such as Uptonia atlantica Fauré and Bohain 2017 within the Masseanum Subchronozone and the genus Dayiceras within the Valdani Subchronozone [Mouterde et al. 1983; Dommergues 1987].
In the lower part of the Valdani Subchronozone [Renzi Lusitanian subzone of Mouterde et al. 1983], there have been six successive species of Dayiceras as a result of processes of gradual evolution [Mouterde et al. 1983; Dommergues 1987] (Figure 5): D. dayiceroides [Mouterde 1951], Dayiceras renzi [Meister 1913] [ = D. quiaosensis Mouterde 1967], D. polymorphoides Spath 1920, D. splendens Mouterde 1967, and D. amaltheiforme Mouterde 1967 with D. nanum Mouterde 1967.
The Dayiceras lineage disappears at a level equivalent to the upper part of the Actaeon Zonule of the North-West European zonal standard (Figure 5). Then, the Eoderoceratidae Metaderoceras beirense Mouterde 1967, remains the only representative of Lusitanian endemism which definitively disappears at the top of the Actaeon Zonule.
5.2. Worldwide distribution of the genus Dayiceras
The Dayiceras genus is the emblematic ammonite of the Lusitanian endemism. It has nevertheless been reported, before us, by many authors outside Portugal:
5.2.1. In the North-West European Province
In the Dorset Coast (Great Britain), where the genus Dayiceras has been defined by Spath [1920], Dayiceras polymorphoides Spath 1920 is precisely positioned stratigraphically by Phelps [1985] in the “Centaurus Zonule”. One can draw a parallel between this level and the Actaeon Zonule of the standard (Valdani Subchronozone, Ibex Chronozone). We also point out to the unique specimen of Dayiceras [D. cf. langi Spath 1920] reported by Hoffmann [1982] in the Masseanum Subchronozone of North-West Germany (Hannover) (Figure 8).
The age of these specimens is identical to that of the Portuguese Dayiceras, show the dispersal of isolated individuals at a distance from the Lusitanian Basin.
5.2.2. In the Tethyan Realm
Dayiceras has been so often reported in the Tethyan Realm that many authors have believed the ubiquitous nature of this genus [Taylor et al. 1984]. We propose here a critical review of all Dayiceras’s occurrences outside of the North-West European Province.
5.2.2.1. Occurrences in the Mediterranean Province
- In the “Tunisian Dorsale”, by Rakús [1972], who describes the new species “Dayiceras” balzeri Rakús 1972 in the Demonense Chronozone of Jebel Staa. But this generic attribution is questioned by Wiedenmayer [1977], who attributes this species, a homeomorph of Dayiceras, to an Eoderoceratidae. It will then be recombined by Rakús and Guex [2002] in the new genus Balzerites and placed, according to the suture lines, within the Ectocentritinae Subfamily. The taxon will then be placed in the Dubariceratinae Subfamily by Venturi et al. [2007].
- In the Italian Calcareous Alps, by Wiedenmayer [1977, pl. 15, figures 3–4], under the taxon “Dayiceras sp. nov. aff. polymorphoides”, and placed in the Polymorphitinae. The species is reassigned to “Dayiceras” bettonii [Parona 1897, pl. X, figure 1a–b] by Wiedenmayer [1980, p. 177], and placed in the Eoderoceratidae.
- In the Apennines (Italy), by Faraoni et al. [1996, pl. 7, figure 3; pl. 10, figures 7, 8], under the names “Dayiceras sp.” and “Dayiceras sp. aff. D. dayiceroides”, from the lower part of the “Gemmellaroi Zone” (Tethyan equivalent of the Valdani Subchronozone). A “Dayiceras sp.” will later be figured by Venturi and Ferri [2001]. All these samples are reassigned to Balzerites balzeri [Rakús 1972] by Venturi et al. [2010] and placed by these authors in the Dubariceratinae Subfamily. The genus Balzerites is now placed into the Ectocentritinae Subfamily by Dommergues and Meister [2017, p. 204].
5.2.2.2. East Pacific Domain (South America)
In Chile and Argentina, Hillebrandt [1987, pl. 2, Figure 11–12] describes “tight-costulated specimens similar to Dayiceras, but without the crenulated keel” in the Meridianus Zone, associated with a closely related species, “Polymorphites (?) sp.” [Hillebrandt 1987, pl. 2, figures 6–10]. All these species will later be attributed to the taxon Eoamaltheus multicostatus [Hillebrandt 2006, p. 143] whose age, specified to the Multicostatus Subzone, may be correlated with the lower part of the North-West European Davoei Chronozone.
The same is true for the “Dayiceras” described by Leanza and Blasco [1990], Dayiceras pseudophylliticum and D. pleuriforme, which Hillebrandt [2006, p. 174] compares, for the first one, to Andidiscus multiforme Hillebrandt 2006, and for the second one, to Andidiscus behrendseni [Jaworski 1926]. These two taxa originate from the Behrendseni Zone, which can be correlated with the upper part of the North-West European Davoei Chronozone (Figulinum Subchronozone).
All these genera and species are endemic to the Andean East Pacific Realm.
5.2.2.3. East Pacific Domain (North America)
The first mention of “Uptonia cf. U. dayiceroides” is given in the Princess Charlotte Island Pliensbachian by Frebold [1970, p. 438, pl. 1, figure 9a–b] but the author specifies that “the absence of crenulation differentiates it from specimens from Mouterde, 1951”. Smith [1983] gave the first detailed and well-argued description of numerous specimens of “Dayiceras dayiceroides” from the Outer Rocky Mountains (Nevada, Oregon, British Columbia, Alberta, Southern Alaska) and Princess Charlotte Island. Smith [1983], Taylor et al. [1984], then Thompson and Smith [1992], deduced that the genus “Dayiceras” was very common in the Tethyan Domain and would be a good marker of Tethyan fauna. According to Smith et al. [1988], all these specimens came from the base of the Freboldi Zone which can be correlated with the top of the Ibex Chronozone of the lower part of the Davoei Chronozone. “Dayiceras sp. [Thompson and Smith 1992, pl. 4, Figures 3–5] from British Columbia is later clearly related to Eoamaltheus multicostatus by Hillebrandt [2006, p. 154]. Like the Andean Cordilleras “Dayiceras”, the north American “D. dayiceroides” are more recent than the Lusitanian Dayiceras.
It is very likely that all of the East Pacific species reported as genus Dayiceras by the authors correspond in fact to primitive Dubariceratidae or Fanninoceratidae [sensu Venturi et al. 2007] and that they are endemic to the East Pacific Domain.
5.3. Paleobiogeographic implications
The genus Dayiceras is absent in the Tethyan Realm, Mediterranean Province, and East Pacific Domain (Chile, Argentina, Canada, United States), in which all of its mentions have since been recombined within the Ectocentritidae, Dubariceratidae or Fanninoceratidae families. It thus appears that the genus Dayiceras depends only on the North-West European Province and that, with the rare exceptions corresponding to the dissemination of isolated individuals in Dorset and North-Western Germany, its paleogeographic distribution is restricted to the Lusitanian Basin which is its evolutionary source.
The discovery, in Vendée, of a population of Dayiceras dayiceroides makes it possible to widen the paleogeographic distribution of the species, because the Vendean populations of this late Polymorphitidae are abundant and their lineage appears well established there. In Vendée, as in the Lusitanian Basin, the species extends the Polymorphitinae clade in the Ibex Chronozone and succeeds to Uptonia atlantica, a species which was also, to date, only known in Portugal. It cannot be a dispersion of isolated individuals, by an Atlantic route, at a distance from a Lusitanian pool, but rather a permanent installation, although of short duration, which makes it possible to significantly extend the paleogeographical distribution of Masseanum and Early Valdani Subchronozones Polymorphitinae. To these two emblematic species of Lusitanian endemism, we can add Acanthopleuroceras carinatum atlanticum.
These ammonites suggest integrating the Lusitanian Basin and Southern Vendée, which paleogeographic reconstructions place close to each other (Figures 2, 8), in the same Atlantic paleobiogeographic area and demonstrate, despite their complexity, that there were connections between the pre-drift basins of the “Iberia–Armorica–Newfoundland” conjugate margin.
As in Vendée, the Lusitanian faunas are clearly dominated by North-Western European influences [Dommergues 1987]. The North-West European polarity of Lusitanian faunas remains unexplained because all the paleogeographic reconstructions of the proto-Atlantic region currently available [Thierry et al. 2000] suggest a large structural opening of the western Iberian Basins towards the Mediterranean Basin of the western Tethys, and the absence of an obvious structural barrier (Figures 2, 8). The demonstration of the Tethyan (Mediterranean) affinities of the Algarve Basin faunas, the southernmost basins of the western Iberian margin (Figures 2, 8), could geographically precise the transition, but does not provide additional explanation for the structural and/or ecological nature of the barrier that separates the two faunal realms [Dommergues et al. 2011]. This barrier, whatever its nature, may have been particularly sealed during the Lower Pliensbachian, a time interval during which the provincialism of the ammonite faunas was the most pronounced [Dommergues 1987]. It suggests a particular tectonic context which could also be the cause of the large proportion of endemic taxa in the Lusitanian Basin. Its attenuation in the Upper Pliensbachian allowed Tethyan taxa, which are numerous in the western Mediterranean Basins (Rif, Middle Atlas, High Atlas) to migrate north toward the “Iberia–Armorica–Newfoundland” conjugate margin. This is especially true for the Late Pliensbachian (Hawskerense Subchronozone), when several waves of Harpoceratinae (Argutarpites, Leioceratoides) and Arieticeratinae (Arieticeras, Emaciaticeras, Canavaria, Tauromeniceras), which are very common in the western Tethyan realm, reached northward the western Iberian Basins. Note that this interval also allowed various typically West European ammonites (Amaltheidae) such as Pleuroceras gr. solare, to reach the western Tethyan Basins and, beyond, part of the Mediterranean Province. The presence of Tethysian ammonites in the Upper Pliensbachian of Southern Vendée is related to this northward migration movement of Mediterranean ammonites. Despite the complexity of the “Iberia–Armorica–Newfoundland”, no obstacle could limit the dispersion of the ammonites in an Atlantic marine area (Figure 8).
The reality of an Atlantic diffusion area is also attested in the Lower Pliensbachian, with an ammonite, Prodactylioceras rectiradiatum [Wingrave 1916], common to the Lusitanian Basin, Southern Vendée, and Dorset [Dommergues et al. 1984]. Argutarpites argutum [Buckman 1909–1930], although not listed in Portugal, seemed to show the same Atlantic distribution area.
This marine area, where we demonstrate the presence, in the Lower Pliensbachian (Lower Ibex Chronozone) and in the Late Pliensbachian (Hawskerense Subchronozone), is from the Early Mesozoic, initiated by an important phase of rifting. It is established on a mosaic of basins, grabens or hemigrabens, linked to the “Iberia–Armorica–Newfoundland” passive conjugate margin, which also includes the Rift of Biscay (Figures 2, 8). Although located between stable western Europe and the Iberian micro-plate, the Rift of Biscay is at the origin of the proto-Gulf of Biscay, a narrow basin which is largely opened on this Atlantic domain (Figure 8). The crustal thinning that affects its margins is attested from the Hettangian in Southern Vendée [Montenat et al. 2003]. It makes the proto-Bay of Biscay and its margins a favored route for the diffusion of Atlantic fauna towards the western borders of the West European Domain (Figure 8).
6. Conclusions. Towards the individualization of an area of Atlantic paleobiogeographic influence
The Pliensbachian Ammonitina under study are clearly related to the North-West European paleobiogeographic Bioprovince (Figure 2). The presence, in the Lower Pliensbachian, of taxa representative of the endemism that trends in the Lusitanian Basin, then in the Upper Pliensbachian, of numerous Tethyan taxa, confers clear originality to the ammonite faunas of Southern Vendée.
The most outstanding result of this study is the demonstration of the persistence, in the Vendean Ibex Chronozone, of several Polymorphitidae whose distribution seemed until now restricted to the Lusitanian region. As in Portugal, these taxa extend the linear evolutionary sequence of Polymorphitidae into the Ibex Chronozone, while it died out everywhere else in the summital Jamesoni Chronozone. It is possible in Vendée, as in Portugal, to study the complete evolutionary sequence between the genera Uptonia and Dayiceras (Figure 5):
- Uptonia atlantica, in the Masseanum Subchronozone, gradually succeeds to U. gr. jamesoni and U. regnardi.
- Dayiceras dayiceroides, first species of the genus, extend the Polymorphitidae lineage in the Valdani Subchronozone.
These two taxa, whose paleogeographic distribution seemed restricted to the Lusitanian Basin, till date, are representative of the endemism that developed during the same time (Masseanum and Valdani Subchronozones) in the Lusitanian Basin. Their discovery in Vendée allows to put into perspective the importance of the Lusitanian endemism at the base of the Ibex Chronozone because the Vendean populations of these late Polymorphitidae are not isolated individuals, but an abundant population that appears to be well settled. This makes it possible to integrate the Lusitanian Basin and Vendée, which paleogeographic reconstructions place close to each other (Figures 2, 8), in the same Atlantic paleobiogeographic area.
It is probably through a diffusion within this Atlantic paleogeographic area that the “southern” affinity taxa such as the genera Matteiceras and Fieldingiceras, but also Tethyan genera (genera Reynesocoeloceras, Arieticeras, Emaciaticeras, Tauromeniceras, Leioceratoides …), which are very numerous in the Upper Pliensbachian of Portugal, would reach Vendée in successive waves and would be able to reach, by this way, the western borders of the North-West European Bioprovince such as, for example, the Basque-Cantabrian region, Asturias, Iberian Ranges, Pyrenees and, to a lesser extent, Dorset and Normandy, as many regions where the Upper Pliensbachian also shows important supplies of Tethyan ammonites.
Without however denying the possibility of Tethyan influences coming from the faunal transition zones, which are well documented on the distant Alpine margins of the European craton [Meister and Stampfli 2000], this Atlantic communication route seems to us to be privileged, at least until the Lower Toarcian.
Conflicts of interest
Authors have no conflicts of interest to declare.
Acknowledgements
We thank Ms. Aloé Hok-Schlagenhauf for checking the English translation. We sincerely thank the two anonymous reviewers and the editor Thomas Saucède for their valuable comments and their constructive suggestions on an earlier version of the article, which allowed a very significant improvement of the manuscript.