


 CC-BY 4.0
CC-BY 4.0
1. Introduction
The Early Cretaceous (Berriasian–Albian; 145.5–99.6 Ma) belemnite faunas of the Tethys have been the subject of numerous studies since the pioneering publications of the 19th century [e.g. Ducrotay de Blainville 1827; Raspail 1829; Duval-Jouve 1841; d’Orbigny 1847]. Since then, the rich faunas of the Vocontian Basin (VB; southeast of France) have been described in detail [e.g., Combémorel 1972, 1973; Gayte and Combémorel 1981; Janssen 2007, 2009, 2018].
For defining taxa, the concepts used in these studies have, however, several set-backs. They suffer either from a limited number of specimens, or are only explained using a qualitative descriptive approach. Efforts for a more objective approach to define belemnite species using quantitative, biometric data go back to the second half of the twentieth century. Biometric data of rostra and their statistical evaluation have been successfully used for defining species and accomplishing phylogenies, particularly for Late Jurassic and Cretaceous species [e.g., Spaeth 1971; Christensen 1975, 2000; Challinor 1979, 1999]. Till date, this biometric approach has not yet been applied to the study of the VB belemnite faunas.
A pre-requisite for such biometric studies is the availability of large belemnite assemblages (>30 specimens) of the conventionally defined species or genera, collected bed-by-bed from stratigraphically well-defined levels. A second step requires a time-consuming biometric analysis of specific features, and a subsequent statistical analysis of data in a third step.
The current study focuses, in the first instance, on a biometric and statistical analysis of 1762 rostra of the belemnite genus Castellanibelus (family Duvaliidae). These specimens have been collected following the standard ammonite zonation of the Valanginian–Hauterivian interval of the Early Cretaceous in the VB [Reboulet et al. 2014]. The data resulted in a taxonomic revision of the genus Castellanibelus. The individual species were then interpreted in their biostratigraphic context. Finally, the abundance patterns of Castellanibelus are compared to those of other co-occurring belemnite genera, in order to obtain quantitative distribution patterns for each ammonite zone.
2. Material
The specimens were collected from the upper surface of beds, following the standard ammonite zones of the standard Kilian group zonation [Reboulet et al. 2014], in the following Az stands for ammonite zones and Asz for ammonite subzones. Most of the specimens come from the departments of Alpes de Haute-Provence, Drôme and Hautes-Alpes. The reference specimens have been deposited in collections of the National Geological Reserve of Haute-Provence.
The genus Castellanibelus, which first appeared in the Tithonian [Combémorel 1972], reached its acme in the Early Valanginian and disappeared subsequently in the latest Valanginian. The biometric analysis performed in this study was carried out on 1762 rostra collected in 112 outcrops exposing Valanginian strata. The investigated exposed surface area in the VB is estimated to be around 4500 km2 based on data from the Geoportail website of Institut national de l’information géographique et forestière [IGN 2022].
The statistical study required the grouping of outcrops into study areas around a reference section. The five selected areas are as follows (Figure 1):
- AGL: Angles-Castellane and Barrème-Moriez area (n = 237); n = number of Castellanibelus studied
- BRT: Barret/Méouge area and Jabron valley (n = 235);
- LBR: Laborel-Rosans and Haute-Ouvèze area (n = 545)
- VGL: Montbrun-Vergol area (n = 593)
- VLX: Villeperdrix-Col Lazarier area (n = 136).
Five isolated outcrops on the margins of the VB yielded 16 additional rostra. These were used in the biometric study but excluded from the biostratigraphic study.
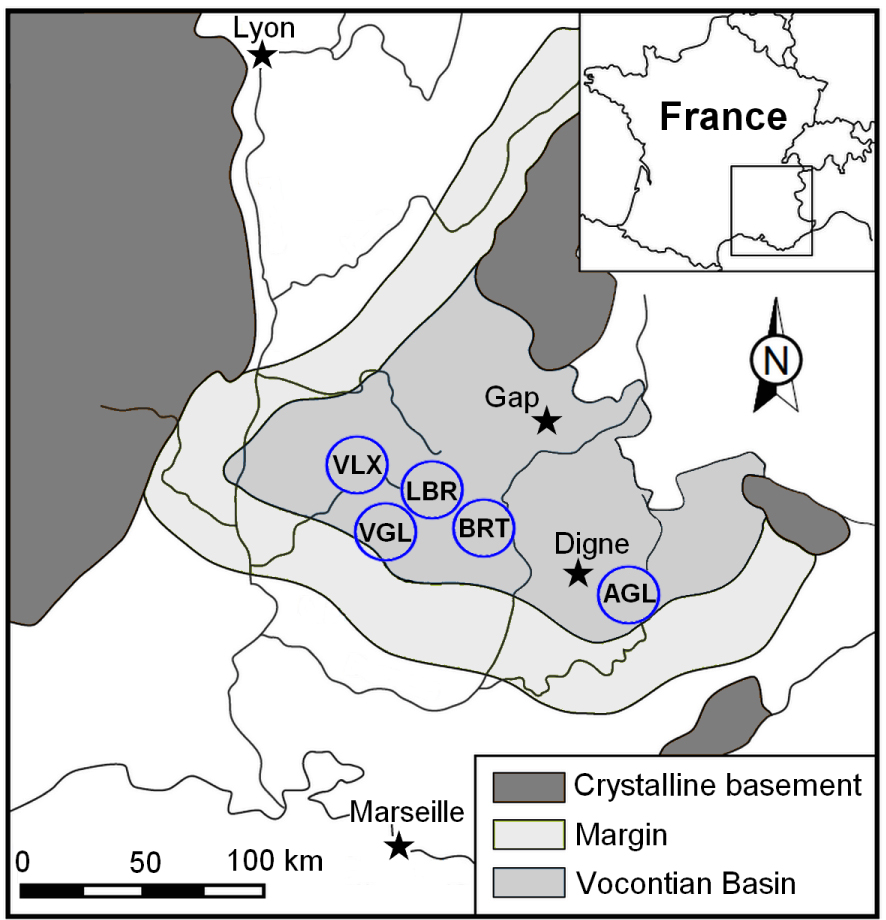
Distribution of study areas in the Vocontian Basin: AGL: Angles-Castellane; BRT: Barret/Méouge and Jabron valley; LBR: Laborel-Rosans; VGL: Montbrun-Vergol, VLX: Villeperdrix; [from Martinez 2013, modified].
Specimens of Castellanibelus are highly abundant throughout the Lower Valanginian Thurmanniceras pertransiens Az up to the basal Upper Valanginian Saynoceras verrucosum Az (Figure 2).
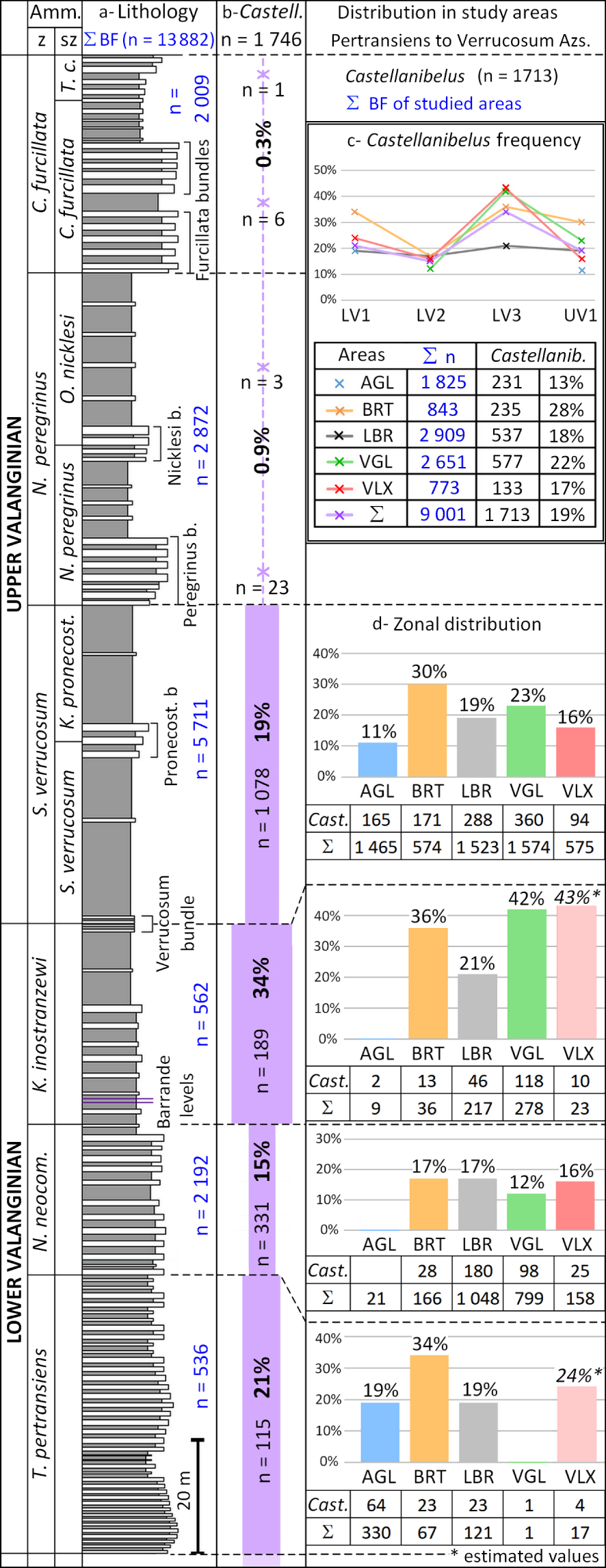
Biostratigraphic distribution of Castellanibelus spp. during the Valanginian by ammonite zone for the entire Vocontian Basin.(a) In blue: total number of rostra of BF per Az; (b) absolute numbers (=n) and relative numbers (%) of Castellanibelus spp. per Az; (c,d) Abundances of Castellanibelus spp. in the studied areas (Pertransiens Az—Verrucosum Az interval); codes for study areas as in Figure 1, [from the synthetic Valanginian lithographic section of Martinez 2013].
Out of a total of 9001 belemnites collected in this interval, in the study area, 1746 rostra were attributed to Castellanibelus (Figure 2a,b). Near the base of the Neocomites peregrinus Az, Castellanibelus suddenly became very scarce. A few specimens were still found in the Peregrinus bundle, above this level the genus becomes extremely rare (0.2% of the rostra). Out of the 4700 rostra collected in the Hauterivian not a single specimen has been attributed to Castellanibelus.
In the Valanginian, in its range of abundance from the Pertransiens Az to the Verrucosum Az, Castellanibelus is abundant and represents about one fifth (∼19%) of the belemnite fauna (BF). It peaks at one third of the belemnite fauna in the Inostranzewi Az.
The spatial and stratigraphic distribution of the 1746 Castellanibelus rostra studied across the VB is congruent with our knowledge of the rest of the belemnite fauna (13,882 rostra in total) long collected in the study area under the same conditions (Figure 2c,d), despite outcrops sometimes being 150 km apart. Only a slight under-representation of the genus in the Laborel sector (LBR), Inostranzewi Az, must be mentioned.
3. Measurements and morphometric analysis
3.1. Measurements
Biometric data have been obtained from the 1762 specimens collected from the Valanginian of the VB. The measurements were made using a digital caliper to an accuracy of 0.01 mm. Each value was measured three times successively for assessment of measurement error.
In order to obtain statistically useful data, measurements of specific features must be linked to tie points. The position of the protoconch is one such tie point used in many studies [e.g. Christensen 1975, 2000]. The position of the protoconch in Castellanibelus is easy to identify because of the regularity of its alveolar cavity. Its median position, relatively close to the apex but anterior to the maximum diameter of the posterior part, does not allow an efficient use of the protoconch-apex length (Figure 3a). The mucro, often broken or incomplete, was excluded from the longitudinal measurements.
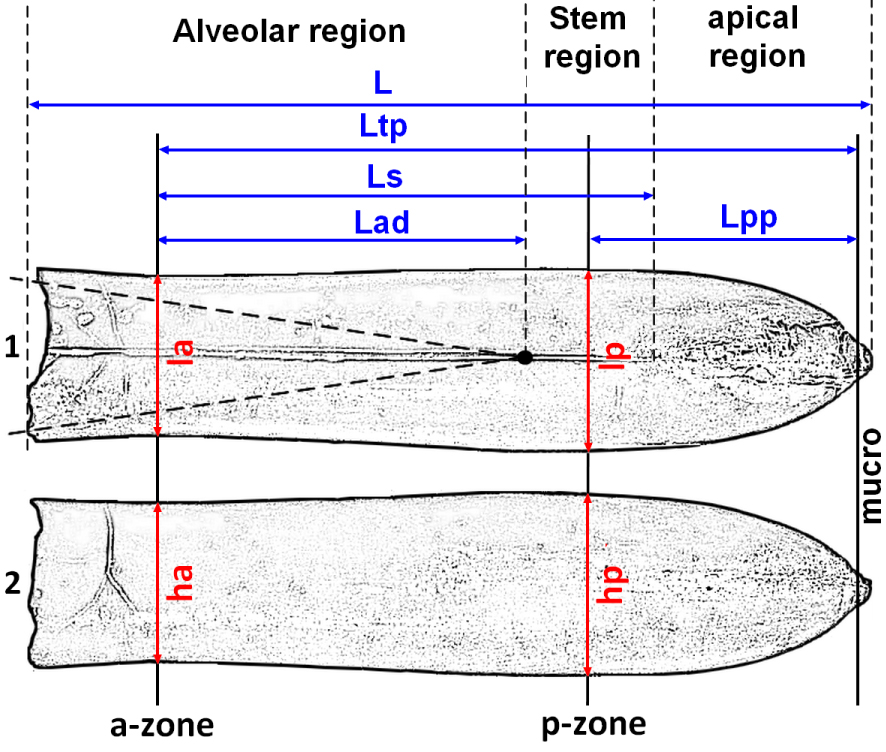
Dorsal (1) and lateral views (2) of a Castellanibelus rostrum, showing the measurements taken for the present study.
The slightly hastate or club-shaped rostra of Castellanibelus show two sectors of minimum and maximum diameters, both in dorsal and lateral views, which are here used as a reference for further measurements. The smallest lateral and dorsoventral diameter ∅, measured in the anterior or alveolar zone (a-zone), marks the first tie point (Figure 3). This definition allows to measure the length of the rostrum (a-zone–apex, =Ltp), excluding the mucro. The posterior zone (p-zone) corresponds to the maximum lateral and dorsoventral diameter ∅ of the rostrum. The resulting second tie point facilitates a second length measurement (p-zone–apex, =Lpp) excluding the mucro. A few rostra show a slight shift between the positions of posterior diameters. In this case, the p-zone retained is the one defined in dorsal view.
The following measurements were used in the current study:
Longitudinal measurements (in mm):
- L: Total length of rostrum.
- Ltp: Reference length (a-zone to apex excluding mucro).
- Lpp: Posterior part length (p-zone to apex excluding mucro).
- Ls: Groove length (a-zone to groove end).
Transverse measurements (in mm)
- ha: Anterior dorsoventral diameter
- la: Anterior lateral diameter
- hp: Posterior dorsoventral diameter
- lp: Posterior lateral diameter
Alveolar measurements
- Lad: Alveolar depth (a-zone to protoconch, in mm).
- Angle of the alveolar cavity (in degrees).
In lateral view, the position of the mucro was assessed as follows: centred (c), dorsal (d) and dorso-centred (dc), (Figure 4).
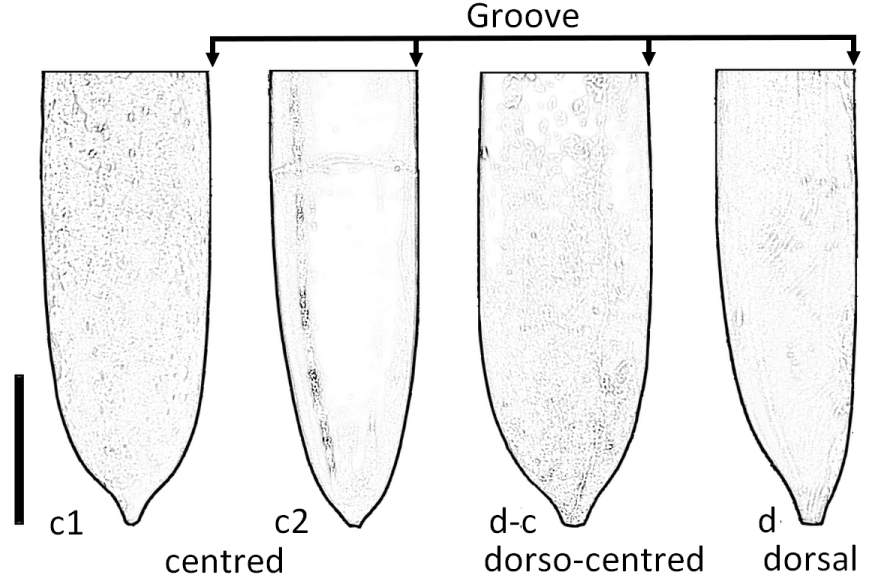
Position of the mucro in lateral view. (c1) centred, C. orbignyanus, no. 92234; (c2) centred, C. suborbignyanus, no. 3025; (d-c) dorso-centred, C. vaubellensis, no. 90872; (d) dorsal, C. toucasi, no. 48583. Scale bar = 1 cm.
3.2. Morphometry and statistics
Biometric data provide a quantitative means of comparing and statistically testing differences among individual rostra of a belemnite assemblage. Because belemnite rostra are subject to allometric growth, biometric ratios rather than raw measurement values are commonly used in statistical analyses of belemnite assemblages comprising individuals of different sizes and ages. Ratios of values taken at the same point on rostra are therefore widely used in belemnite biometry suggesting that they are reliable tools [e.g. Spaeth 1971; Mutterlose et al. 1983].
The following ratios have been computed for the present study:
- ica (ha/la): anterior compression index
- icp (hp/lp): posterior compression index
- iddv (lp/la): dorsoventral dilation index
- idlat (hp/ha): lateral dilation index
- Psr (Lpp/lp): posterior shape ratio
- apical angle (INVTAN[(lmax∕2)∕Lpp]) × 2))
- alveolar relative depth (Lad∕Ltp)
Calculations and graphs were accomplished with an Excel database. The arithmetic means and standard deviations were calculated to the nearest hundredth. Many graphics show a polynomial curve of degree 2, used to analyse variations in a large data set. The full data and calculations are available in the Supplementary Information.
3.3. Belemnite populations and ontogeny
In biology, the term “population” is defined as a group of organisms of a species co-existing in an area, i.e. organisms which reproduce sexually with each other. The usage of the term “population” is thus problematic when analysing a belemnite genus which may well consist of several species. Another problem is the duration of 0.6–0.8 Ma of individual Valanginian ammonite zones, and even 1.4 Ma in the case of the N. peregrinus Az. These long time intervals, which reflect our belemnite assemblages, are clearly excluding the use of the term “population”. We therefore prefer the term “assemblage” when discussing the biometric data of specific, stratigraphically bounded rostra.
For the statistical analysis, the rostra have been grouped as assemblages in the following way. For each of the five studied areas, the available belemnites have been assigned to one of the 6 standard ammonite zones of the Valanginian zones [standard Kilian group zonation—Reboulet et al. 2014]. In this manner, five assemblages have been evaluated for each ammonite zone.
Daily increments of rostrum growth [Hoffmann, R., Stevens, K. 2019] can be observed in rostrum longitudinal sections (Supplementary Information, Plate S5, Figure S1). They show a continuous growth of the rostra which is not contradictory with potential allometric growth. For the ontogenetic study, the 1746 rostra assemblage was partitioned into five growth stages. The criteria used were rostra length (L) and/or cavity depth (Lal).
The shortest rostra correspond to shapes observed on the first growth increments. They are therefore comparable to the most juvenile individuals (mj) (mj: L ≈ 21 mm, Lal ≈ 4.5 mm). In contrast, the longest rostra are here interpreted as adult forms (a) of Castellanibelus (a: L ≈ 58 mm, up to 70 mm [Combémorel 1972], Lal ≈ 20.5 mm). Three other categories present intermediate forms between these two extreme stages. They are named here very juvenile (vj) (vj: L ≈ 28 mm, Lal ≈ 6.5 mm), juvenile (j) (j: L ≈ 41 mm, Lal ≈ 11.5 mm) and subadult (sa) (sa: L ≈ 50 mm, Lal ≈ 16 mm). These last designations have no absolute biological meaning but only correspond to distinct growth stages in terms of size and shape.
4. Biometric results
4.1. Ontogeny
4.1.1. Rostrum length and alveolar depth
Measuring the length of rostra between the protoconch and the anterior extremity, and between the protoconch and the apex requires that rostra be complete, a requirement fulfilled by only 251 out of the 1746 studied rostra (=14%).
The shortest measured length of a complete specimen is 15.6 mm long (no. 98790, Verrucosum Az), the largest complete one 67.6 mm (no. 79022, Inostranzewi Az). Adults longer than 60 mm were found from the Pertransiens Az to the Verrucosum Az.
Alveolar depth changes during growth, from 37% on average for juveniles to over 50% for adults (extreme values are comprised between 17 and 67%, n = 875). The increase in alveolar depth is continuous in juvenile stages (mj–j), and it slows down strongly in subadult and adult stages, when the width of rostra is maximum.
4.1.2. Compression indices
The compression index (ic) describes the lateral compression in the anterior (ica = ha/la) or posterior part of the rostrum (icp = hp/lp). Three conditions can be met: laterally compressed (ic > 1), circular (ic ≈ 1), or dorsoventrally depressed (ic < 1). Overall, Castellanibelus rostra show a slight depression (mean ic = 0.92, n = 861). The degree of depression varies according to the region of the rostrum, the taxonomy (Supplementary Information 1.A) and the growth stage, with most juvenile rostra showing a sub-circular posterior cross-section.
Most juvenile rostra (mj) are sub-circular in the apical region (mean icp = 1) and slightly depressed in the alveolar region (mean ica = 0.92). Later ontogenetic stages (vj–a) show a steady shift to more depressed apical regions, due to an allometric increase of width with growth (Figure 6). The adult phase (a) is depressed in the apical region (icp = 0.89) and less so in the alveolar region (ica = 0.94). Throughout ontogeny, the area of maximum depression is shifting from its alveolar position in juvenile specimens to a more apical one in adults. These data document an allometric growth of Castellanibelus in width, the lateral growth rate obviously exceeding the dorsoventral one (Figure 5).

Transverse section through the p-zone of one of the oldest adult rostrum showing growth increments and the progressive increase in width of the rostrum (icp = 0.84, no. 79759).
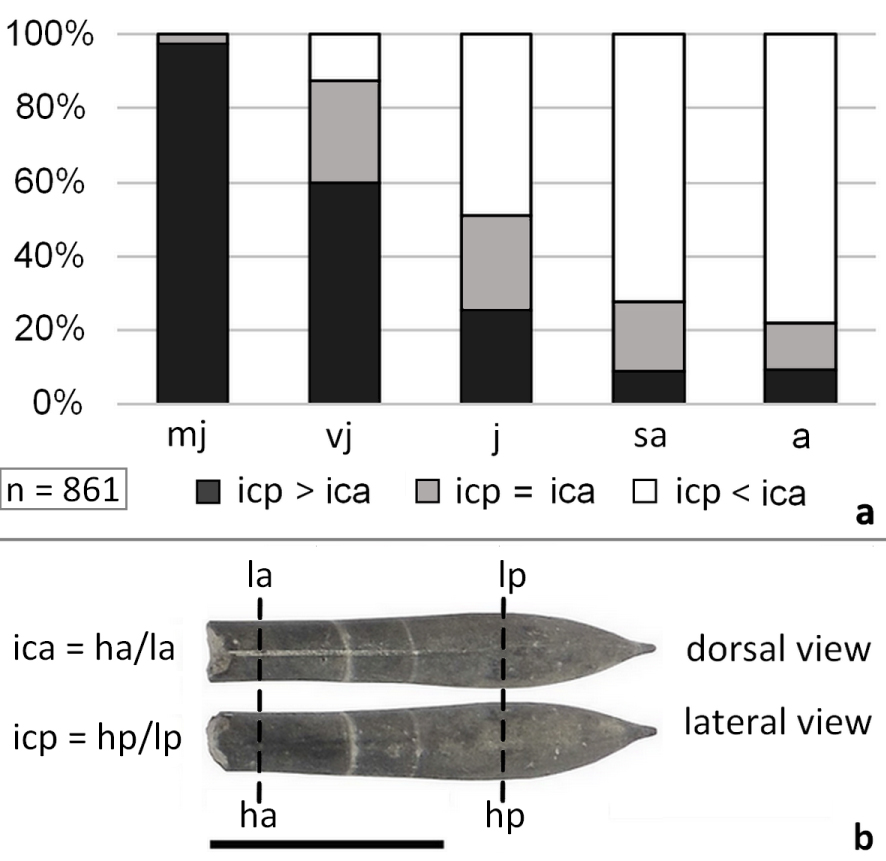
(a) Ontogenetic stages and compression index for Castellanibelus: icp > ica: rostrum more depressed anteriorly; icp = ica: rostrum depressed anteriorly as posteriorly; icp < ica → rostrum more depressed posteriorly. (b) One of the most juvenile rostrum, depressed anteriorly and sub circular posteriorly (ica = 0.97, icp = 1.03, no. 4924, scale bar = 1 cm).
The degree of depression of the posterior part of the rostra decreased during the Valanginian. For adult specimens, the icp values increased from 0.85 in the Pertransiens Az (icp = 0.85) to 0.91 in the Verrucosum Az (icp = 0.91; n = 157).
This evolution towards more circular cross-sections is linked to a change in taxonomic composition: the dominant taxon at the base of the Valanginian, which is very depressed, tends to disappear progressively in favour of other, less depressed taxa of Castellanibelus.
4.1.3. Dilation indices
The dilation index (id) describes the shape of the rostrum in dorsoventral (dv) or lateral (lat) view. Three conditions are possible: hastate, or club shaped rostra (id > 1); cylindrical rostra (id ≈ 1); conical rostra (id < 1).
The mean dilation indices of Castellanibelus document a slightly hastate shape in general (iddv = 1.12; idlat = 1.11, n = 861).
The mean dilation indices change with growth stages. They show a greater dilation in the early stages of growth (iddv = 1.15, idlat = 1.22). This dilation then evolves towards less dilated forms in the following growth stages (iddv = 1.11, idlat = 1.10). Finally, there is a further increase in the dilatation index in dorsolateral view in the longest rostra (iddv = 1.13, idlat = 1.08; Figure 7, Supplementary Information 1.A).
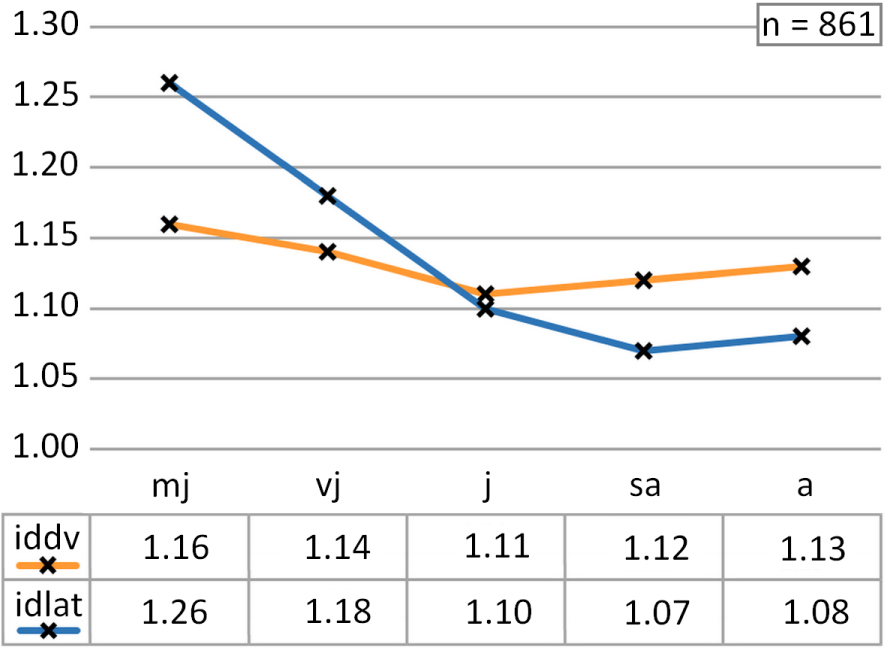
Distribution of mean values of dorsoventral (iddv) and lateral (idlat) dilation indices for the different ontogenetic stages.
The dilation of the rostra of Castellanibelus is therefore varying according to ontogenetic stages. Three phases can be distinguished. The rostra of very juvenile specimens are slightly hastate (mj/vj). This dilation phase is followed by an elongation of the rostra which corresponds to the juvenile stages (j/sa). The rostra then show a sub-cylindrical shape. Then, with the thickening of the posterior part of the rostra, the mature rostra again show a slightly hastate outline in dorsolateral view.
No significant changes of shape have been observed throughout the Valanginian.
4.1.4. Alveolar groove and position of mucro
The alveolar groove, positioned dorsally, is narrow and long (mean Ls = 74% of L, n = 864). It never reaches the apex (maximum 92% of L). Its length increases with age from 65% in most juvenile forms to about 76% in adults.
The growth of the rostrum is asymmetrical in the apical region. The mucro gradually shifts with rostrum length to a more centred position: only 11% of the juveniles show a centred mucro compared to 42% of the adults (n = 1423, Supplementary Information 1.B).
4.1.5. Growth key
Using morphological features (e.g. shape, mucro, sulcus, lateral lines), previous authors concluded that there were no significant ontogenetic changes in Castellanibelus [e.g. Duval-Jouve 1841; d’Orbigny 1847; Combémorel 1972; Janssen 2018], an assessment based solely on qualitative observations.
Present biometric results document significant morphological changes of the rostrum with growth. These changes are summarised in Figure 8.
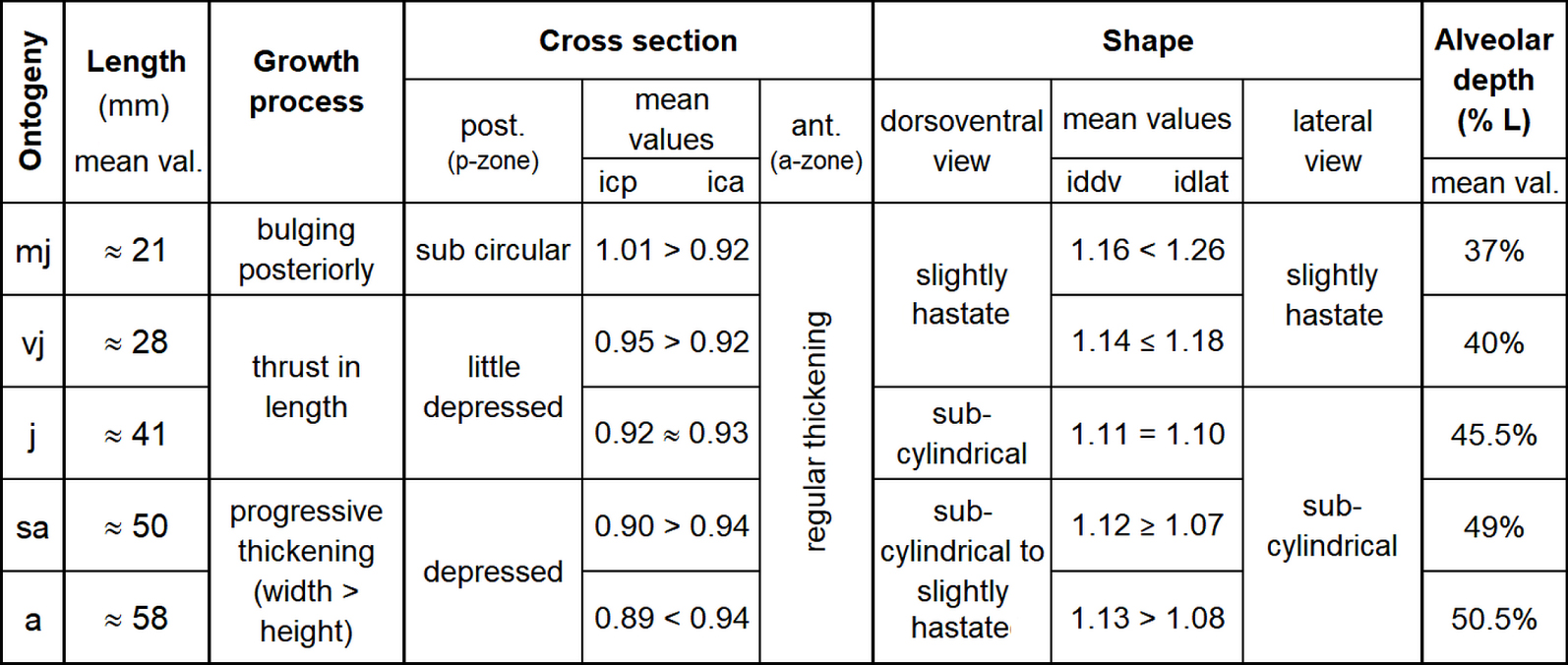
Measurements of the five ontogenetic stages of Castellanibelus; icp, n = 1416; ica, iddv, idlat, n = 861; length and alveolar depth, n = 875.
4.2. Morphotypes and stratigraphy
The following biometric indices of Castellanibelus show a significant variation across the Valanginian: the Posterior shape ratio (Psr), the apical angle and the Posterior compression index (icp) (Figure 9 and Supplementary Information 1.A, 1.B, 1.C).
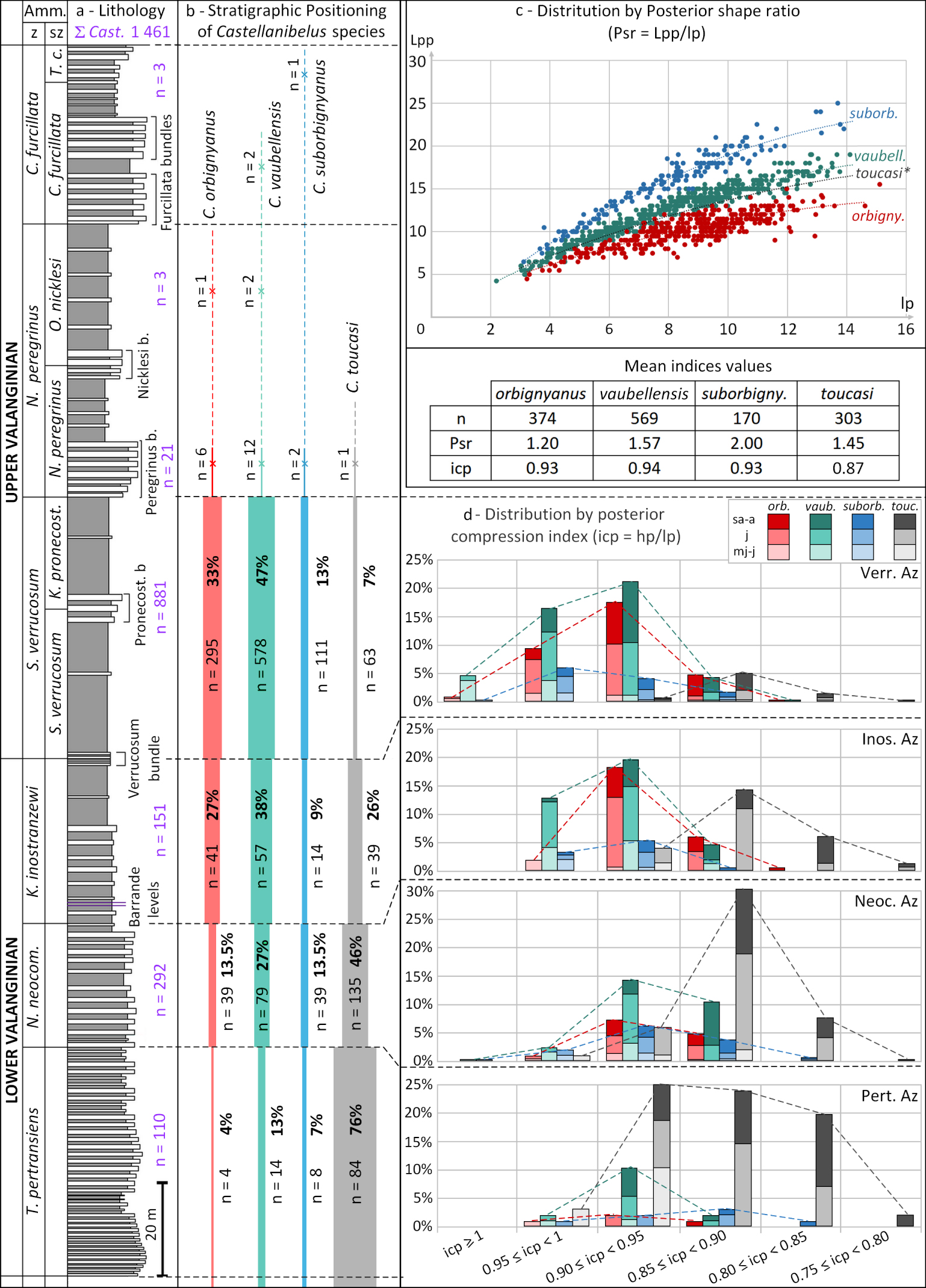
Stratigraphic distribution (a–b) and biometric indices (c–d) of Castellanibelus species during the Valanginian [from Martinez 2013]; (a) number of Castellanibelus specimens collected per Az; (b) absolute number (n) and percentage (%) of specimens for each Castellanibelus species: red = C. orbignyanus, green = C. suborbignyanus, blue = C. vaubellensis and grey = C. toucasi; (c) distribution of posterior shape ratio for each species, polynomial curves give the general trend for each species (*C. toucasi is only represented by its polynomial curve for a better readability); (d) distribution of the posterior compression index values by ammonite zone in the Castellanibelus abundance interval (Pertransiens Az – Verrucosum Az); for each species, colour intensity refers to the different ontogenetic stages: light (mj–vj), medium (j), dark (sa–a). Masquer
Stratigraphic distribution (a–b) and biometric indices (c–d) of Castellanibelus species during the Valanginian [from Martinez 2013]; (a) number of Castellanibelus specimens collected per Az; (b) absolute number (n) and percentage (%) of specimens for each Lire la suite
4.2.1. Posterior shape ratio and apical angle
The Psr values are also controlled by ontogeny, the earliest ontogenetic stages (mj) having a higher relative posterior length than adults. The Psr ratio varies considerably between specimens and indicate for <1 a very short posterior length (<1), for a median posterior length (±1.50) and a very long posterior length (for >2). Extreme values of Psr range between 0.73 and 2.76.
Based on the examination of 1416 specimens, Psr values document three distinct morphotypes (A, B, C; Figure 9c; Supplementary Information 1.B). A fourth morphotype, D, differs significantly in its posterior compression index (icp), described in Section 4.2.2. The Psr values are closely related to the size of the apical angle, where a short posterior length is mirrored by a large apical angle.
- A: median length posterior, mean Psr = 1.57; extreme values between 1.99 (most juvenile) to 1.13 (adult), mean apical angle = 36°, n = 569;
- B: short posterior length, mean Psr = 1.20, extreme values between 1.66 (most juvenile) to 0.73 (adult), mean apical angle = 46°, n = 374;
- C: long posterior length, mean Psr = 2.00, extreme values between 2.76 (most juvenile) to 1.58 (adult), mean apical angle = 28°, n = 170;
- D: median posterior length, mean Psr = 1.45, extreme values between 1.94 (most juvenile) to 0.97 (adult), mean apical angle = 39°, n = 303.
4.2.2. Compression indices
The anterior (ica) and posterior (icp) compression indices show a steady decrease in depression during the Valanginian:
- Pertransian Az: mean ica = 0.89; mean icp = 0.89
- Verrucosum Az: mean ica = 0.93; mean icp = 0.93.
This decrease in depression is due to morphotype D that is more depressed than other morphotypes, especially posteriorly: mean icp D = 0.87; icp A, B and C = 0.93, Figure 9d).
Morphotype D is clearly dominant at the base of the Valanginian: from 76% of all Castellanibelus in the Pertransian Az, it decreases in abundance steadily and represents only 7% of Castellanibelus in the Verrucosum Az. This decreasing abundance of morphotype D explains the overall increase in compression indices observed due to morphotypes A, B and C.
The strong depression of D, both anteriorly and posteriorly, is a characteristic dorso-ventral flattening, which results in a sub-quadrangular to oval shape in cross-section. A trend towards dorso-ventral flat rostra was also observed for mature rostra of morphotypes A, B, C, where it is less distinctive and only developed in the posterior part.
4.2.3. Mucro position
The position of the mucro in lateral view has been quantified semi-quantitatively (mucro centred, dorso-centred, dorsal; Figure 4), based on 1423 rostra. The position also varies with ontogeny (Supplementary Information 1.B).
Two of the four morphotypes show a mucro in a centred position: C (77%, n = 172), B (67%, n = 385). The rostra of morphotype A have a dorso-centred mucro (85%, n = 567). Group D has a mucro between dorso-centred and dorsal, depending on the ontogenic stage (dorsal: 55%, dorso-centred 43% and 2% centred, n = 299).
4.3. Biostratigraphy
Specimens of Castellanibelus represent about one fifth of the belemnite fauna in the Pertransiens Az to Verrucosum Az interval (Figure 2). The four morphotypes (A, B, C, D) are not evenly distributed across this stratigraphic interval (Figure 9).
In the Lower Valanginian Pertransiens Az, morphotype D is very frequent (76% of all Castellanibelus specimens). Morphotypes A, B and C are present, but rather rare. The abundance of morphotype D gradually decreases in the Neocomiensiformis Az (46%) and Inostranzewi Az (26%), while morphotypes A and B become more common. The Verrucosum Az sees a peak of A and B (47%, 33%), while D is rare (7%). Morphotype C is stable throughout the entire Valanginian ( ≈12%).
Castellanibelus becomes very rare at the transition with the Peregrinus Az. A few rare resilient specimens of all four morphotypes are still present in the Peregrinus bundle (Figure 9). A single specimen of morphotype C was found at the top of the Valanginian stage in the Teschenites callidiscus Ammonite subzone.
5. Taxonomy
5.1. Species concept in the genus Castellanibelus
Toucas [1890] proposed two varieties of the species created by Duval-Jouve [1841]: Belemnites orbignyanus var. jouvei and Belemnites orbignyanus var. suborbignyi. These creations were not taken up subsequently. When the genus Castellanibelus was created, Combémorel [1972] considered it to be a mono-specific genus. All rostra in this genus were attributed to the species Castellanibelus orbignyanus until the subsequent revisions and creations by Janssen [1997, 2003, 2007, 2018] and Janssen and Clément [2002].
In the present study, biometric data demonstrate the existence of four distinct species within the genus Castellanibelus, three were attributed to earlier creations (morphotypes A, B and C), the fourth (morphotype D) requiring the erection of a new species.
5.1.1. Type species
The short, obtuse apical part, the centred or slightly eccentric mucro and the slight depression of the rostra assigned to morphotype B are consistent with the diagnosis of the species Castellanibelus orbignyanus [Duval-Jouve 1841]. The type figure is a juvenile and not an adult specimen as thought by Duval-Jouve [1841] (L = 46 mm). The juvenile rostrum no. RHP.04236-026.AA.516 (Figure 10B2), figured here, is very close.

Castellanibelus species from the Valanginian of the Vocontian Basin. Scale bar 1 cm. (A) C. vaubellensis [Janssen 2018], (B) C. orbignyanus [Duval-Jouve 1841], (C) C. suborbignyanus [Toucas 1890], (D) C. toucasi sp. nov.. Very juvenile (1), juvenile (2) and adult (3) forms for each of the 4 species. References: A1 (no. RHP.04236-026.AA518), A2 (no. RHP.04204-024.AA522), A3 (no. RHP.26018-001.AA514), B1 (no. RHP.04236-026.AA517), B2 (no. RHP.04236-026.AA516), B3 (no. RHP.26018-001.AA513), C3 (no. RHP.30173-001.AA515)—Upper Val., Verrucosum Asz.; C2 (no. RHP.05135-002.AA521), D1 (no. RHP.26153-001.AA523)—Lower Val., Neocomiensiformis Az.; C1 (no. RHP.04210-001.AA519), D2 (no. RHP.04210-001.AA520), D3 (no. RHP.04179-026.AA524)—Lower Val., Pertransiens Az. Masquer
Castellanibelus species from the Valanginian of the Vocontian Basin. Scale bar 1 cm. (A) C. vaubellensis [Janssen 2018], (B) C. orbignyanus [Duval-Jouve 1841], (C) C. suborbignyanus [Toucas 1890], (D) C. toucasi sp. nov.. Very juvenile ... Lire la suite
Castellanibelus (Belemnites) picteti [Mayer 1866] shows the same characters and corresponds to the type species orbignyanus.
5.1.2. Variety suborbignyi Toucas 1890
Morphotype C is characterized by a minor depression, an elongation of the apical part which ends in a mostly centred mucro (Figures 10C1–C3). These criteria are similar to those of the Castellanibelus variety suborbignyi [Toucas 1890], which is thus raised to the species status under the name Castellanibelus suborbignyanus [Toucas 1890].
5.1.3. Morphotypes A and D
Morphotype A has an apical part of medium length, a mucro in intermediate position associated with a weak posterior depression (mean icp = 0.94). The intermediate positioning of the mucro is correlated with a more flattened dorsal surface in dilated adults and a characteristic posterior bulge in lateral view (Figure 10A3).
While these criteria match with those of the holotype of Castellanibelus vaubellensis Janssen 2018, some elements of the diagnosis of this species require revision (sub-hastate form and dorso-ventral flatness). The dilation of the rostra of Castellanibelus is variable and cannot be used as a determination criterion. This variability is due to ontogeny (see Section 4.1.3) and to a more or less strong growth in width of the adult rostrum according to the individuals, inducing shapes going from nearly sub-cylindrical to sub-hastate in all morphotypes (Supplementary Information 1.A, 1.C).
Janssen [2018] reports a dorso-ventral flattening in C. vaubellensis. Adult rostra of all morphotypes tend towards a posterior dorso-ventral flattening due to the increase in width with age. When this flattening occurs all along the rostrum and appears in juvenile stages, it induces the significant depression characteristic of morphotype D. The juvenile paratype of C. vaubellensis [Janssen 2018] (Jans. Figures 7.3–4) shows such a depression, accompanied by the dorso-ventral flattening attributable to D.
Morphotype D is characterised by its strong depression, its dorso-ventral flatness inducing an oval to sub-quadrangular cross-section, its predominantly dorsal mucro, and its stratigraphic peak in the Lower Valanginian (Figures 10D1–D3). The distinctive depression of this morph group is mentioned by Pictet and de Loriol [1858, ic = 0.81] and Combémorel [1972, ic ≈ 0.85]. A sub-quadrangular section or a flattening of the sides is also indicated by Janssen and Clément [2002, Castellanibelus sp. B] and Janssen [2003, Castellanibelus sp. A]. No valid species exhibiting the characters of morphotype D, it is assigned to a new species, Castellanibelus toucasi sp. nov.
5.1.4. Comparison with other species
Janssen [2003] proposed the classification of Conobelus triquetus Weiss 1991, in the genus Castellanibelus. If the depression (0.73) of the rostrum of Weiss would tend to bring it closer to Castellanibelus, the shallow depth of the alveolus strongly eccentric, the groove reaching the apex, the absence of lateral lines, the absence of a mucro are incompatible with a positioning within the genus Castellanibelus. C. triquetus should therefore be returned to its original genus, Conobelus.
Castellanibelus? bonti Janssen 2007, was created from a single specimen showing characters contradictory with the diagnosis of the genus (very shallow alveolar cavity, very dilated asymmetric shape, prominent alveolar groove…). Its leaf-like shape allows a very probable attribution to a malformed specimen of Duvalia emerici [Raspail 1829]. The organ responsible for the presence of the alveolar groove has slipped laterally, inducing a flattening of one face and a bulging of the other [forma aegra dissulcata Keupp 2012].
5.2. Revision of the genus Castellanibelus
Order Belemnitida Zittel, 1895
Suborder Belemnopseina Jeletzky, 1966
Family Duvaliidae Pavlow, 1914
Genus Castellanibelus [Combémorel 1972]
Type species: Belemnites orbignyanus Duval-Jouve 1841, Plate 8, Figures 4–9.
Diagnosis: Rostrum slender, up to 68 mm long; dorsoventrally depressed, except for very juvenile stages, which are sub cylindrical; anteriorly less depressed than posteriorly; slightly hastate in outline and profile; dorsal alveolar groove narrow, not reaching mucronate apex; alveolus circular, reaching half of the rostrum in adult stage, ∼18°; double lateral lines faint, sometimes reduced to simple flat surfaces.
Discussion: Combémorel [1972] established the genus Castellanibelus and assigned it to the family Duvaliidae. This classification was based on 3 rostra showing a dorsal position of the groove, opposite to the siphuncle. This observation is here confirmed for 62 studied specimens (Supplementary Information 3, Plate S5; Figures 1–6).
Castellanibelus differs from the other genera of Valanginian Duvaliidae (Duvalia, Berriasibelus, Pseudobelus) by its depression, which can be observed on the whole rostrum, more or less important depending on the area of the rostrum, the species and the ontogenic stage. Other characteristic features of the genus are the mucronate apex, the alveolar depth (18°, centred, 1∕2 of the adult rostrum), the long narrow groove not reaching the apex and the presence of lateral lines.
Stratigraphic distribution: Castellanibelus is reported as soon as the Tithonian [Combémorel 1972], it is common throughout the Lower Valanginian and basal Upper Valanginian (Verrucosum Az) where it represents about 20% of the total belemnite fauna (BF). Abundance decreases dramatically near the boundary of the Verrucosum Az/Peregrinus Az, only very rare specimens have been observed in the Peregrinus Az and Furcillata Az (0.4% of BF).
Geographical distribution: Southeast of France, the type species coming from the Castellane region which gave its name to the genus. Common in the Vocontian Basin (Alpes-de-Haute-Provence, Drôme, Hautes-Alpes, Ardèche, Gard). Reported from Switzerland [Ooster 1857; Pictet and de Loriol 1858; Mayer 1866], Czech Republic [Vaňková 2015] and Spain [Janssen, e.g. 1997, 2003].
Castellanibelus orbignyanus [Duval-Jouve 1841]
Figures 10.B1–B3; Appendix 3, Plate S1, Figures 1–22
1841 Belemnites orbignyanus Duval-Jouve, p. 64–65, Plate 8, Figures 4–9.
non 1847 Belemnites orbignyanus Duval: Orbigny, p. 8–9, Plate 4, Figures 10–16.
1857 Belemnites orbignyanus Duval-Jouve: Ooster, p. 23, Plate 1, Figures 9–10.
non 1858 Belemnites orbignyanus Duval-Jouve: Pictet and Loriol, p. 8–9, Plate 1, Figures 6–7.
1866 Belemnites picteti Mayer, p. 366.
non 1867 Belemnites orbignyanus Duval-Jouve: Pictet, p. 54, Plate 8, Figure 2.
non 1868 Belemnites orbignyanus Duval-Jouve: Pictet, p. 217–218, Plate 36, Figure 3.
non 1873 Belemnites orbignyanus Duval: Gilliéron, p. 204–205, Plate 8, Figure 11.
1878 Hibolites orbignyi Duval: Bayle, atlas Plate 31, Figures 9–12.
non 1890 Belemnites orbignyi Duval: Toucas, p. 587–588, Plate 15, Figures 1–3.
? 1920 Conobelus orbignyanus Duval-Jouve: Bulow-Trummer, p. 177–178.
? 1963 Curtohibolites orbignyanus (Duval-Jouve): Stoyanova-Vergilova, p. 214–215, Plate 1, Figure 1.
pars 1972 Castellanibelus orbignyanus (Duval-Jouve, 1841): Combemorel, p. 75–77, Plate A, non Figures 12–14.
non 1988 Curtohibolites orbignyanus (Duval-Jouve): Ali-Zade, p. 394–395, Plate 1, Figure 4.
pars 2003 Castellanibelus orbignyanus (Duval-Jouve): Janssen, p. 142–143, Plate 2, Figures 11–12, non Figures 6–7, 13–14.
pars 2018 Castellanibelus orbignyanus (Duval-Jouve): Janssen, p. 175, Figures 7: 7–9, ?18–21, non 10–11.
Type: Type-species of the genus, introduced by Duval-Jouve [1841]. One adult and two juvenile specimens are designated here as paratypes (Figures 10B1–B3).
Material: 386 rostra, Valanginian, Southeast France.
Measurements: adult mean values (n = 44)
ica = 0.95; icp = 0.90; iddv = 1.12; idlat = 1.06; apex angle = 52°.
Diagnosis: Rostra medium size, maximum observed length 60 mm; slightly depressed anteriorly and posteriorly, increasing posteriorly with age (icp mean: juveniles’ 0.97 to adults’ 0.90); dorsal side tending to flatten with age; moderate dilatation; long alveolar groove extending into apical region, not reaching the apex; apical region short to very short, becoming obtuse, apical angle increasing with age (juveniles’ mean 39°, adults’ mean 52°, up to 68°); mucro centred or slightly offset dorsally.
Differential diagnosis: C. orbignyanus differs from related species (C. suborbignyanus, C.vaubellensis) by its short apical region and its very obtuse apex and from C. toucasi by its lesser depression and its obtuse apex and its more centred mucro.
Stratigraphic distribution: Reported by authors in the Berriasian [Pictet 1867; Toucas 1890]; rare at the base of the Valanginian (Pertransian Az 14% of all Castellanibelus specimens), getting common in the Neocomiensiformis Az and Inostranzewi Az. C. orbignyanus reaches its acme in the Verrucosum Az (33%) and is very rare from the Peregrinus Az onwards. The last occurrence is found in the Nicklesi Asz.
Geographic distribution: Common in the Vocontian Basin (Southeast France), reported from Switzerland [Ooster 1857; Pictet and de Loriol 1858; Mayer 1866], Czech Republic [Vaňková 2015] and Spain [Janssen, e.g. 1997, 2003].
Castellanibelus suborbignyanus [Toucas 1890]
Figures 10.C1–C3; Appendix 3, Plate 2, Figures 1–23.
? 1847 Belemnites orbignyanus Duval: Orbigny, Plate 4, Figure 15.
1867 Belemnites orbignyanus Duval-Jouve: Pictet, p. 54, Plate 8, Figure 2.
1868 Belemnites orbignyanus Duval-Jouve: Pictet, p. 217–218, Plate 36, Figure 3.
? 1873 Belemnites orbignyanus Duval: Gillieron, p. 204–205, Plate 8, Figure 11.
1890 Belemnites orbignyi Duval-Jouve var. surborbignyi Toucas, p. 588, Plate 15, Figure 2.
1997 Castellanibelus sp. A Janssen, p. 6, 8, Plate 3, Figures 7–8.
pars 2003 Castellanibelus orbignyanus [Duval-Jouve 1841]: Janssen, Plate 2, Figures 13–14.
Type: Variety of Belemnites orbignyanus introduced by Toucas [1890], raised to the species status here. One adult and two juvenile specimens are designated here as paratypes (Figures 10C1–C3).
Material: 174 rostra, Valanginian, Southeast France.
Measurements: adult mean values (n = 17)
ica = 0.94; icp = 0.90; iddv = 1.09; idlat = 1.07; apex angle = 31°.
Diagnosis: Rostra maximum observed length 60 mm; slight depression anteriorly and posteriorly; increasing posteriorly with age (icp mean: juveniles’ 1.00 to adults’ 0.90); very moderate dilation (around 1.08 in dorsal and lateral views) shape tending towards sub-cylindrical; distinctive dorsal alveolar groove, slightly shorter than in other Castellanibelus; apical region elongate, with very acute (juveniles, mean 21°) to acute (adults, mean 31°) apical angle; mucro centred or slightly dorsally offset.
Differential diagnosis: C. suborbignyanus differs from other species of Castellanibelus by the greater length of its posterior part. This gives it an acute alveolar angle and an elongated outline/profile, more sub cylindrical, and by its slightly shorter dorsal groove.
Stratigraphic distribution: Reported by Pictet [1867] and Toucas [1890] in the Berriasian. Present from the base of the Valanginian [Janssen 1997, this study], in the Pertransiens Az (7% of all Castellanibelus specimens), its population remains around 12% up to the top of the Verrucosum Az., very rare above. One specimen found at the top of the Valanginian (Callidiscus Asz).
Geographic distribution: Southeast of France [Pictet 1867; Toucas 1890; this study], Switzerland [Pictet 1868] and southeast of Spain [Janssen 1997, 2003].
Castellanibelus vaubellensis Janssen 2018
Figures 10.A1–A3; Appendix 3, Plate 3, Figures 1–20.
1890 Belemnites orbignyi Duval-Jouve var. jouvei Toucas, p. 588, Plate 15, Figure 3.
1972 Castellanibelus orbignyanus (Duval-Jouve): Combemorel, Plate A, Figures 12–14.
pars 2018 Castellanibelus vaubellensis Janssen, p. 175–176, Figures 7.5–6, non 1–4.
Type: The holotype of Janssen 2018 is here supplemented by three paratypes: one adult and two juvenile specimens (Figures 10A1–A3).
Material: 579 rostra, Valanginian, Southeast France.
Measurements: Adult mean values (n = 77)
ica = 0.94; icp = 0.91; iddv = 1.14; idlat = 1.11; apex angle = 41°.
Diagnosis: Rostra as long as 68 mm and slightly depressed anteriorly and posteriorly; the depression increases with age (icp mean: juveniles 1.01, adult 0.91), the flattening of the dorsal side being compensated by a characteristic bulge of the ventral side; the dilation is moderate (mean 1.13) but can be more accentuated (up to 1.39) giving individuals a more hastate outline/profile, both dorsoventrally and laterally; dorsal furrow long, fading at the maximum width, apical part of median size (juv. 31°, adult 41°); mucro intermediate in position, between centred and dorsal.
Differential diagnosis: C. vaubellensis is distinguished by the median length of its posterior part, between the obtuse part of C. orbignyanus and the acute part of C. suborbignyanus. It differs from C. toucasi by its less pronounced depression and swollen ventral side in the posterior part.
Stratigraphic distribution: Reported by Toucas [1890] from the Berriasian and by Janssen [2018] from the Lower Valanginian (Neocomiensisformis Az), C. vaubellensis is common from the base of the Valanginian (Pertransiens Az, ∼13% of all Castellanibelus specimens) and becomes the dominant species from the Inostranzewi Az. Abundance of C. vaubellensis peak in the Verrucosum Az (∼47% of all Castellanibelus specimens) before declining abruptly at the transition with the Peregrinus Az. Very rare above the Peregrinus bundle, the last occurrences were found in the Furcillata Asz.
Geographic distribution: Southeast of France [Toucas 1890; Janssen 2018; this study].
Castellanibelus toucasi sp. nov.
Figures 10.D1–D3, Appendix 3, Plate 4, Figures 1–22
? 1858 Belemnites orbignyanus Duval-Jouve: Pictet and Loriol, p. 8–9, Plate 1, Figures 6–7.
? 1972 Castellanibelus orbignyanus (Duval-Jouve): Combemorel, p. 81, Plate 3.
2002 Castellanibelus sp. B sp. nov. Janssen and Clement, p. 520
2003 Castellanibelus sp. A Janssen, p. 144, Plate 2, Figures 3–7.
pars 2018 Castellanibelus orbignyanus [Duval-Jouve 1841]: Janssen; Figures 7.10–11.
pars 2018 Castellanibelus vaubellensis nov. sp. Janssen, Figures 7.3–4.
Locus typicus: 04200 Saint-Geniez, Alpes-de-Haute-Provence.
Stratum typicum: base of the Pertransiens Az.
Derivatio nominis: in homage to Aristide Toucas [1843–1912] who was the first to observe the variability within the genus Castellanibelus.
Type series: Adult holotype (Figure 10D3, no. 3028), Pertransiens Az, Lower Valanginian, Saint-Geniez locality. Paratypes: very juvenile (Figure 10D1, no. 43568), Neocomiensiformis Az, lower Valanginian, Laborel locality; juvenile (Figure 10D2, no. 88233), Pertransiens Az, Lower Valanginian, Soleilhas locality.
Material: 319 rostra, Valanginian, Southeast France.
Measurements: Adult mean values (n = 23)
ica = 0.91; icp = 0.83; iddv = 1.15; idlat = 1.04; apex angle = 45°.
Diagnosis: Rostra with a maximum length of 63 mm; markedly depressed anteriorly and more posteriorly (mean icp 0.87, up to 0.75 for adults); dorsal and ventral sides flattening, inducing an oval to sub quadrangular posterior cross-section; moderate dilation, especially in lateral view (mean idlat 1.09); long dorsal groove, extending apically without reaching apex; apical part of median size, apical angle increasing with age (juveniles’ mean: 32°, adults’ mean: 45°, up to 50°); mucro dorsally offset in juvenile stage, dorso-centred thereafter.
Differential diagnosis: C. toucasi differs from other species of Castellanibelus by its anteriorly and posteriorly more marked depression (C. toucasi mean icp = 0.87; other Castellanibelus mean icp ≈ 0.94), its dorso-ventral flatness and its mucro tending towards a more dorsal position. Showing an apical angle close to that of C. vaubellensis, it differs in its posterior sub-quadrangular cross-section and in the absence of the ventral bulge observed in C. vaubellensis.
Stratigraphic distribution: C. toucasi occurs from the Berriasian to the mid-Valanginian. The species is frequent at the base of the Valanginian, in the Pertransiens Az (∼76% of all Castellanibelus specimens). Its abundance gradually decreases and shows a clear decline from the Inostranzewi Az onwards. C. toucasi is rare in the Verrucosum Az (∼7% of all Cast. specimens) and disappears in the Peregrinus Az (last occurrence at the base of the Peregrinus bundle).
Geographic distribution: Southeast of France [Combémorel 1972; this study], Switzerland [Pictet and de Loriol 1858] and southeast of Spain [Janssen 2003].
Taxonomic key: The Figure 11 summarizes the different taxonomic variations.
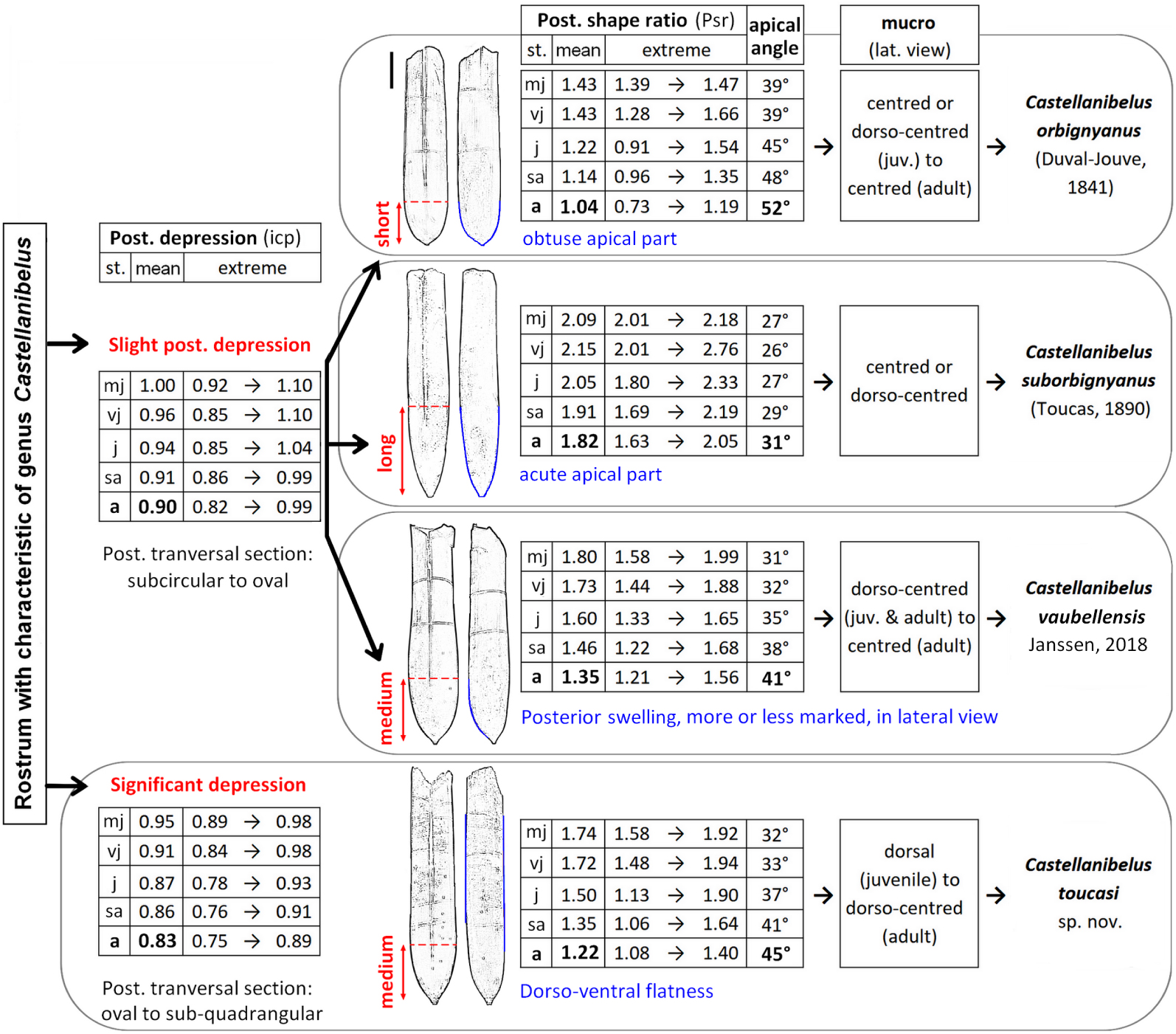
Taxonomic key of Castellanibelus species based on index values for the different ontogenetic stages.
6. Conclusions
A biometric study of 1762 rostra of the belemnite genus Castellanibelus, collected in the Valanginian marls of the Vocontian Basin (VB; Southeast France), has provided insights into the ontogeny, evolution and phylogeny of this group of cephalopods. In the VB, throughout the Valanginian, the genus Castellanibelus is common and evenly distributed in all five study areas and in stratigraphy, implying a cohesive distribution of belemnite faunas across the VB. In terms of abundance, Castellanibelus specimens represent about 1/5th of the entire Valanginian belemnite faunas in the Lower Valanginian-Upper Valanginian interval (from the Thurmaniceras pertransiens to the Saynoceras verrucosum ammonite zone). The genus becomes very rare from the middle Upper Valanginian onwards (base of the Neocomites peregrinus ammonite zone), with only a few occurrences in the Uppermost Valanginian. It disappears at the top of the stage.
Species of the genus Castellanibelus show an allometric growth. Juvenile forms have a sub-circular cross-section and an elongated shape. Adult forms show a depressed cross-section and a sub-cylindrical to slightly hastate shape.
Biometric data enable differentiation of four species of Castellanibelus varying by the degree of depression and shape of the apical part of the rostrum. The taxonomy of the species Castellanibelus orbignyanus [Duval-Jouve 1841] and Castellanibelus vaubellensis [Janssen 2018] has been revised. Castellanibelus surborbignyanus [Toucas 1890] has been raised to the species level and one new species, Castellanibelus toucasi sp. nov. is proposed.
Conflicts of interest
The author has no conflict of interest to declare.
Acknowledgements
I would like to thank my husband, Patrick, for his unfailing support and involvement, my father, Gérard Thomel, who collected half of the specimens, Jörg Mutterlose and Thomas Saucède for their precious help and encouragement, and Didier Bert, curator of the Haute-Provence Geological Reserve, for his kind permission to carry out research.