

1 Introduction
Central African vegetation history is characterized by important changes in forest cover and composition. Global climatic changes throughout the Pleistocene and the Holocene had a major impact on flora distribution in the tropics. During cold and dry periods, such as the Pleistocene glaciations, savannah vegetation types were predominant in the Central African landscape (Colyn et al., 1991; Maley, 1996). Climate-induced vegetation dynamics also had an important effect on prehistoric human settlement, migration and subsistence. Central Africa is a key region for one of the major demographic events in African prehistory (Pakendorf et al., 2011), i.e. the so-called ‘Bantu Expansion’, which started ca. 4000 BP from a homeland in the Nigerian–Cameroonian borderland (Blench, 2006: 134, 136; Nurse and Philippson, 2003: 164; Vansina, 1995: 52).
Bantu speech communities occupy at present the greatest part of the equatorial rainforest as well as most of eastern and southern Africa as far as the Cape. The Bantu language family is by far Africa's largest one, both in terms of number of speakers and tongues as well as in terms of geographical spread, even if linguists consider it as only a fairly recent offshoot of the larger Niger–Congo language phylum. The most recent common ancestor of the Niger–Congo languages, spoken from Senegal to the Cape, is believed to have a time depth of 10,000 to 12,000 years, at least twice as old as the most recent common ancestor of the Bantu languages. The oldest ceramic traditions in South Africa, commonly associated with the first Bantu speakers there, date back to the first half of the first millennium AD (Phillipson, 2005). Taking this into account, the spread of Bantu speakers from their homeland to the southern extreme of the continent took less than 3000 years, much less than what their Niger–Congo ancestors needed to settle throughout western Africa.
This rapid expansion of Bantu speech communities over large parts of sub-Saharan Africa has fascinated generations of scholars from a wide variety of disciplines. Linguists, archaeologists, palynologists, molecular anthropologists and many more have debated on the driving forces behind the Bantu Expansion. Two interconnected factors that have received relatively little attention in this debate – certainly compared to agriculture and iron production – are climate change and vegetation dynamics. It has nevertheless been postulated that different stages in the Bantu Expansion were enabled, or at least facilitated, by climate-induced vegetation shifts.
Lavachery (2001), for instance, suggests that the ancestors of the Bantu speakers reached the Nigerian–Cameroonian borderland, from where the Bantu Expansion would start, as climate refugees. The serious climatic deterioration that occurred around 7100–6900 BP in the Sahara and the Sahel and the desiccation it induced would have forced populations to move from the Sahel southwards into the Guinean Gulf. The climate there was favourable to forest extension (Maley and Brenac, 1998). These newcomers introduced new languages, new technologies and a forest plant food complex, which underwent local adaptations and transformations before being diffused in West Central Africa (Bostoen, forthcoming). Neolithic technologies, such as macrolithic tools, polishing, and pottery, turn up in the archaeological record of the Grassfields region of Cameroon from around 7000 BP, but undergo a definitive breakthrough around 5000 BP, slightly anterior to the assumed beginning of the Bantu Expansion (Bostoen, 2007; Lavachery, 2001).
Between 4500 and 4000 BP, the Central African rainforest underwent an important climate-induced contraction at its periphery with considerable savannah development, especially along the coasts of Gabon and Congo (Ngomanda et al., 2009) and also with the extension of the Niari savannahs in Congo (Vincens et al., 1994, 1998). This contraction was contemporaneous with the opening of the Dahomey Gap further north (Salzmann and Hoelzmann, 2005). At that stage, however, the central evergreen forests were not affected (Maley, 2004).
The central rainforest was not disturbed before the middle of the first millennium BC when another major perturbation took place. It is particularly this second period of forest perturbation that has been linked with the early Bantu Expansion. As argued by Schwartz (1992), Maley (2001) and Oslisly (2001), the opening of the rainforest in the course of the first millennium BC may have given a boost to the migration of Bantu-speaking farming, pottery-making and iron-producing populations into regions that had been more difficult to access until then. Numerous palynological and geological data obtained over the last two decades testify indeed to a serious disturbance of the Central African rainforest during that period (Elenga et al., 2000; Maley and Brenac, 1998; Vincens et al., 1994, 1998). Pollen profiles from lake sediments in Cameroon and Congo manifest a distinct decrease of mature forest taxa, coupled with an increase in pioneers and/or grasses (Ngomanda et al., 2009). The appearance of the so-called ‘Sangha Interval’, a kind of large corridor composed of savannahs and open forests in the western part of the Congo basin linking the northern savannahs to those of the Batéké Plateau in the south is seen as one of the major impacts of this and previous forest perturbation phases. The present-day vegetation in this area still manifests traces of the past penetration of savannah plant species, such as refuge areas of Phoenix reclinata palm (Doumenge et al., 2012). Similarly, the savannah-forest mosaic zones of Gabon and Congo are home to certain fauna species, such as the Pygmy Mouse or Mus minutoides (Muridae) (Mboumba et al., 2011) or the bushbuck or Tragelaphus scriptus (Bovidae) (Moodley and Bruford, 2007), which have northern affinities. Their presence in the area can best be explained through migration from Cameroon, which was only possible thanks to past forest openings during the Pleistocene and Holocene.
The discovery of domesticated pearl millet (Pennisetum glaucum) in two southern Cameroonian archaeological sites, dated between 2400–2200 BP, gave further support to this hypothesis (Eggert et al., 2006). This savannah crop could only be cultivated due to the development of a distinct dry season. This increased seasonality was also beneficial to the availability of edible plant resources, such as yams, and made the clearing of secondary vegetation less arduous, since fast-growing pioneer trees produce softer woods (Kahlheber et al., 2009; Neumann et al., 2012). All these factors, linked with the emergence of forest-savannah mosaics, may indeed have favoured demographic growth and given an impetus to the expansion of early Bantu-speaking communities further south.
In this article, we deal with the question of knowing whether these Late Holocene palaeoclimatic changes in western Central Africa did indeed have an impact on the Bantu Expansion, by focusing on a body of evidence that is not commonly used to examine vegetation dynamics, i.e. language data. If the contraction of the Central African rainforest that emerged during the first millennium BC or earlier really played a role in the Bantu Expansion, these climate-induced vegetation changes are likely to have had their impact on the languages spoken by these immigrant groups and to have left their traces in the current-day Bantu languages that descended from these ancestral languages. In Section 3 of this article, we analyse common Bantu vocabulary for three pioneer species abundantly present in the Central African pollen record between 2500 and 2000 BP, i.e. Musanga cecropioides, Elaeis guineensis, and Canarium schweinfurthii. The geographical distribution patterns of the common Bantu vernacular names for these three pioneer species provide linguistic evidence adding impetus to the hypothesis that the significant rainforest contraction that emerged in the first millennium BC had an effect on the way Bantu languages expanded. Before discussing these specific linguistic data, we first explain, in Section 2, for readers not familiar with linguistics, how historical linguistic methods can be used to reconstruct environmental history.
2 Words as history
A language is an intangible cultural heritage. As long as it does not die out, it is transmitted from one generation to the next. As a system of communication, it is sensitive to the changing needs of its speakers. Transformations in the outside world may trigger change in a language too, especially in the lexicon. New words are coined or borrowed from neighbouring languages or old words get different meanings. Each language is the product of its speakers’ history and can therefore be used as a body of historical evidence. In this way, language can also be relied on to study vegetation history. Changes in the natural habitat tend to be reflected in today's languages, even if these changes happened a long time ago. This becomes particularly clear if one examines languages from a comparative point of view. Bantu languages are very rewarding in this respect, because they cover an immense portion of the African continent, but are still closely related due to their relatively young age. They still share a lot of vocabulary, which facilitates language comparison. Many western Bantu languages share, for instance, the generic term *-téndé for oil palm (E. guineensis).1 They inherited this word from Proto-Bantu, the ancestral language of all Bantu languages, which inherited it itself from a still older ancestor. This indicates that the ancestors of Bantu speakers were already familiar with this plant long before they started to wander in Central Africa. Most eastern Bantu languages lost this term, simply because the oil palm does not thrive in the savannahs of eastern and southern Africa. In some eastern Bantu languages, however, this word was retained, but refers now to other palm tree species, among others the date palm (Phoenix dactylifera) (Bostoen, 2005). Similarly, the Proto-Bantu word *-gàdí, which commonly designates oil palm nuts or palm oil – reddish in colour – in western Bantu languages was conserved in eastern Bantu languages through a metaphorically induced semantic shift. It became the common word for blood (Bostoen, 2005). These examples show nicely how changes in the natural habitat of communities become rooted in their languages and leave traces that are still retrievable today.
Historical linguists thus examine lexical and other resemblances between related and non-related languages. Apart from chance similarities, which can easily be excluded, shared vocabulary is indicative of shared history. Words that resemble one another in form and meaning are either inherited from a common ancestor and spread through the dispersal of its daughter languages or were adopted through contact and spread across languages as loanwords. Inherited vocabulary can only be shared between related languages. Loanwords can be transmitted between both related and non-related languages if these happen to be in contact. Once borrowed, a word can become inherited, if it is handed on to new generations of speakers. Historical linguists make use of the Comparative Method to distinguish between inherited and borrowed vocabulary by relying on regular sound correspondences. These cannot be historically fortuitous, since they are recurrent, systematic and without unexplainable exceptions. While synchronically widespread inherited terms can be reconstructed into a putative proto-language via these regular sound changes, loanwords cannot. The geographical distribution of loanwords may show us cultural diffusion paths across languages. The distribution of inherited vocabulary reflects spread coinciding with the dispersal of languages. Moreover, historical linguists interpret time as a function of space. A word's distribution among sub-groups of a language family gives an idea of its relative time depth with respect to the internal classification of this family. As regards Bantu, an inherited term can be reconstructed in Proto-Bantu, if it occurs in enough different Bantu sub-groups, or a more recent proto-language, such as Proto-East Bantu or Proto-South-West Bantu, if it only occurs in languages belonging to one of these specific sub-groups. The above-mentioned word *-téndé for oil palm, for example, occurs not only in western and eastern Bantu languages, but also in related non-Bantu languages from the Niger–Congo phylum. This provides us with evidence, both internal and external to Bantu, that this specific word can be reconstructed in Proto-Bantu, which inherited this term itself from an older pre-Bantu language stage. Absolute dating of language evolution is a rather tricky issue. Among the methods developed to calculate the absolute time elapsed since related languages split from a common ancestral language, glottochronology is the best known (Swadesh, 1955), but also the most contested one amongst linguists (Blench, 2006: 40–42). Vocabulary is not always replaced at a constant rate over time, as Swadesh (1955) assumed when he developed an arithmetic formula to convert the rate of lexical similarity between two languages in the absolute amount of time since they separated. Absolute dating of language evolution is only possible when certain nodes in a language family tree can be convincingly calibrated with historical or archaeological dates. This is often difficult in Central Africa due to a lack of good archaeological data for several regions of Central Africa (Clist, 2012).
Another problem when studying vernacular names for wild plants species across languages is the lack of good lexical data. When using comparative Bantu linguistics for the reconstruction of Central African history, as described above, it is crucial to have enough data from sufficiently diverse languages. Botanical sources, such as flora catalogues from African countries or ethnobotanical studies, provide vernacular names, but these are often limited to few languages. Moreover, the transcription of the sounds and tones of these language data are usually not transcribed according to the rules, which hampers a rigorous historical linguistic interpretation. Conversely, lexicographical sources on Bantu languages often provide better data in terms of phonological transcription, but the information they contain on vernacular plant names is often restricted to common crops. If they provide Bantu names for wild species, these are often not properly identified botanically. Obtaining good comparative ethnobotanical language data thus requires dedicated interdisciplinary fieldwork (Koni Muluwa, 2010). Hence, the choice to focus in this article on the common Bantu names for M. cecropioides, E. guineensis, and C. schweinfurthii is not only inspired by the fact that these pioneer species are well represented in the Central African pollen record between 2500 and 2000 BP, but also by the fact that they are amongst those wild tree species for which most lexical data are available in current-day Bantu languages.
3 Linguistic evidence for pioneer tree species in Central Africa
3.1 Musanga cecropioides
The M. cecropioides species, also known as umbrella tree in English or parasolier in French, is a short-lived, light-demanding pioneer colonizer that naturally thrives in large clearings in the tropical forest zone. It participates in the first stage of rainforest rebuilding. It also occurs in swamp forests and along rivers or lakesides (Burkill, 1985; Hutchinson and Dalziel, 1954; Orwa et al., 2009). The species is absent from most of eastern and southern Africa, where eastern Bantu languages are spoken. Hence, *-céngà and *-gùmbù, the two common Bantu names of this species, chiefly occur in western Bantu languages, where they have a geographic distribution that is more or less complementary. This is significant from a historical point of view.
The first term can be reconstructed as *-céngà. There is no doubt at the origin of the tree's scientific name Musanga, although this very term has not been found in a Bantu language spoken today. The tree's name in Kikongo, the language from which other species also drew their scientific name, is musenga, which might have been erroneously transcribed as Musanga. Attestations of this term have been found in about 40 Bantu languages. As shown in Figs. 4–6, those languages are spoken in six different countries: Cameroon, Equatorial Guinea, Gabon, Congo, Democratic Republic of the Congo, and Angola. Regarding the internal classification of Bantu languages (Bastin and Piron, 1999; Bastin et al., 1999; Grollemund, 2012; Vansina, 1995), the term is distributed over two major historical sub-groups, i.e. North-West Bantu (Table 1) and West-Coastal Bantu (Table 2). From a historical point of view, it is important to note that the phonetic shape of this vernacular name for M. cecropioides is diverse across languages. This is due to the fact that most forms underwent the respective sound changes to which their language was subject through time. This indicates that we are dealing with inherited vocabulary transmitted through time from an ancestor language to its descendants as opposed to loanwords transmitted through space between neighbouring languages. The current-day geographic distribution of this vernacular name can therefore be explained as the result of language expansion rather than as the outcome of diffusion through language contact, although early contact may have played a role, as discussed in Section 4. Finally, historically speaking, it is not insignificant, as can be observed on Fig. 1, that this geographic spreading is fairly contiguous and continuous.
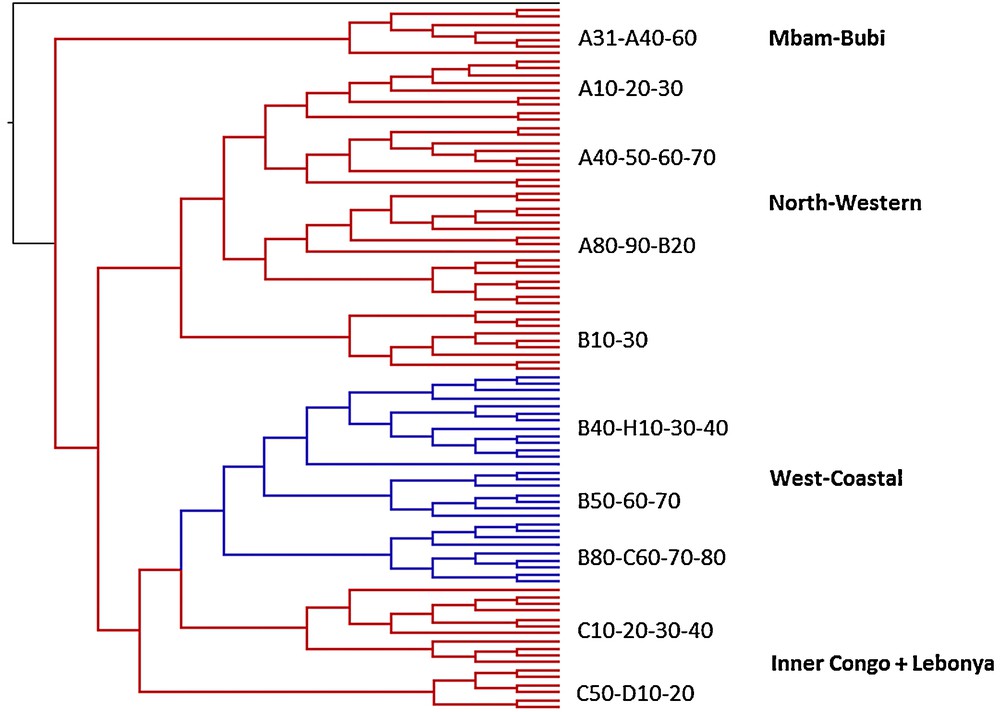
Mapping of common Bantu names for pioneer species on the phylogenetic tree of Bantu languages (Grollemund, 2012). Mapping of vernacular Elaeis guineensis names on the phylogenetic tree of the Bantu languages: *-bá (blue) and *-bídà (red).
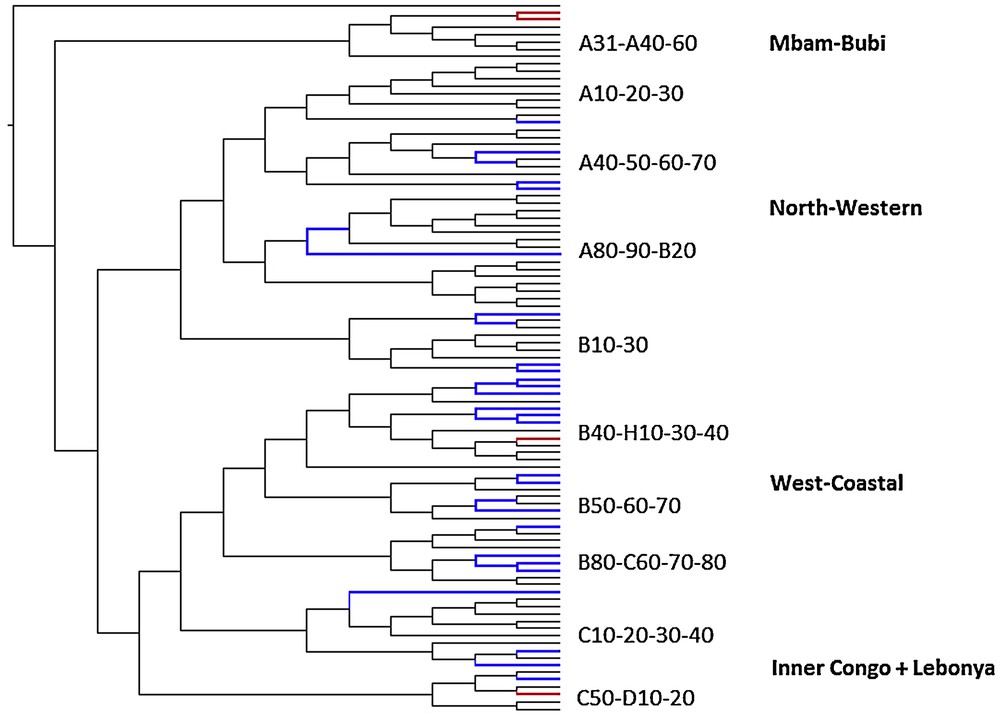
Mapping of common Bantu names for pioneer species on the phylogenetic tree of Bantu languages (Grollemund, 2012). Mapping of vernacular Canarium schweinfurthii names on the phylogenetic tree of the Bantu languages: *-bídí (blue) and *-pátù (red).
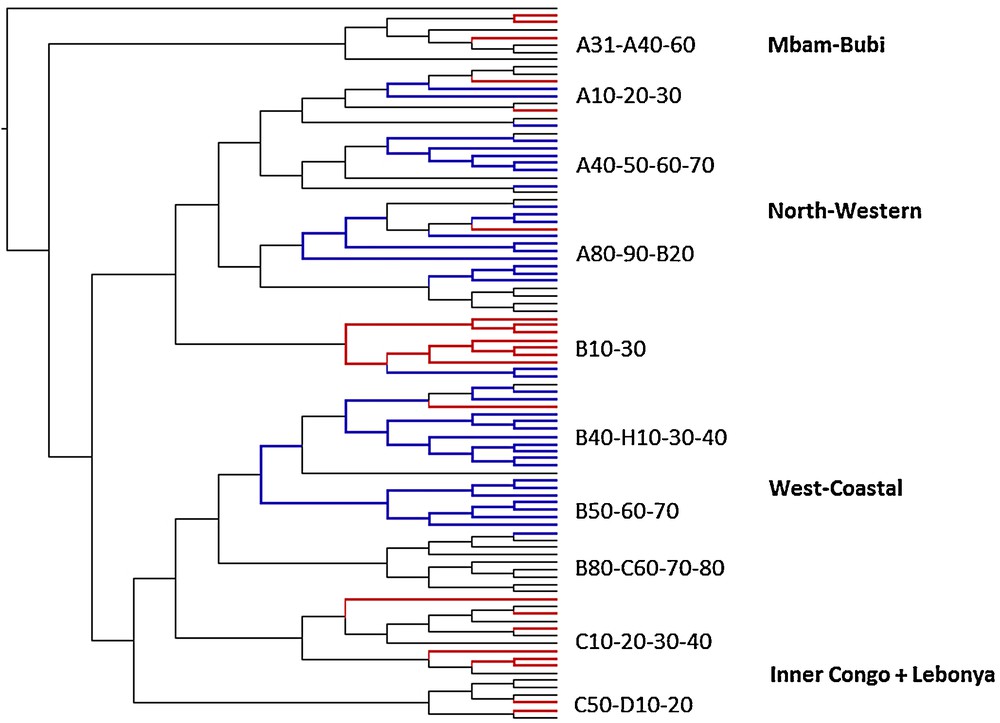
Mapping of common Bantu names for pioneer species on the phylogenetic tree of Bantu languages (Grollemund, 2012). Mapping of vernacular Musanga cecropioides names on the phylogenetic tree of the Bantu languages: *-céngà (blue) and *-gùmbù (red) reflexes.
Vernacular Bantu names for Musanga cecropioides. Attestations of *-céngà in North-West Bantu languages.
| Language | Country | Term | Source |
| Mokpwe (A22) | Cameroon | lisengi | (Burkill, 1985) |
| Duala (A24) | Cameroon | bosénge | (Helmlinger, 1972; Burkill, 1985) |
| Duala (A24) | Cameroon | asseng | (Vivien and Faure, 1985) |
| Basaa (A43a) | Cameroon | lisseng | (Vivien and Faure, 1985) |
| Bafia (A53) | Cameroon | (Guarisma, 1969) | |
| Eton (A71) | Cameroon | esen | (Swartz, 1989) |
| Bulu (A74) | Cameroon | assan | (Vivien and Faure, 1985) |
| Gyele (A81) | Cameroon | léséng | (Vivien and Faure, 1985) |
| Makaa (A83) | Cameroon | (Heath, 1989) | |
| Njem (A84) | Cameroon | lèsíà | (Beavon, 2003) |
| Bajue (A841) | Cameroon | esséa | (Nguenang et al., 2010) |
| Koonzime (A842) | Cameroon | esía | (Beavon and Beavon, 1996) |
| Bekwil (A85b) | Cameroon | (Phillips, 2009) | |
| Kwakum (A91) | Cameroon | (Belliard, 2005) | |
| Kako (A93) | Cameroon | (Ernst, 1989) | |
| Fang (A75) | E. Guinea | aseng | (Akendengué, 1992) |
| Benga (A34) | Gabon | ihèngè | (Raponda-Walker and Sillans, 1995) |
| Fang (A75) | Gabon | asèng | (Raponda-Walker and Sillans, 1995) |
| Seki (B21) | Gabon | disyèngóko | (Raponda-Walker and Sillans, 1995) |
| Kele (B22) | Gabon | disyènghè | (Raponda-Walker and Sillans, 1995) |
| Koya (B22F) | Gabon | dísyɛŋa | (Medjo Mvé, 2011) |
| Kota (B25) | Gabon | ihèndjè | (Raponda-Walker and Sillans, 1995) |
| Geviya (B301) | Gabon | mosɛnga | (Van der Veen and Bodinga-bwa-Bodinga, 2002) |
| Vove (B305) | Gabon | mosènga | (Raponda-Walker and Sillans, 1995) |
| Tsogo (B31) | Gabon | mosènga | (Raponda-Walker and Sillans, 1995) |
Vernacular Bantu names for Musanga cecropioides. Attestations of *-céngà in West-Coastal Bantu languages.
| Language | Country | Term | Source |
| Vungu (B403) | Gabon | musènga | (Raponda-Walker and Sillans, 1995) |
| Sangu (B42) | Gabon | musènga | (Raponda-Walker and Sillans, 1995) |
| Punu (B43) | Gabon | musènga | (Raponda-Walker and Sillans, 1995) |
| Lumbu (B44) | Gabon | musèngè | (Raponda-Walker and Sillans, 1995) |
| Duma (B51) | Gabon | musèngè | (Raponda-Walker and Sillans, 1995) |
| Nzebi (B52) | Gabon | musèngè | (Raponda-Walker and Sillans, 1995) |
| Ambede (B61) | Gabon | osyè | (Raponda-Walker and Sillans, 1995) |
| Mbaama (B62) | Gabon | osyè | (Raponda-Walker and Sillans, 1995) |
| Ndumu (B63) | Gabon | osyè | (Raponda-Walker and Sillans, 1995) |
| Vili (H12) | Gabon | nsènga | (Raponda-Walker and Sillans, 1995) |
| Teke (B70) | Congo | onssié | (Adjanohoun et al., 1988) |
| Laari (H16f) | Congo | nsenga | (Adjanohoun et al., 1988) |
| Nsong (B85b) | RDC | (Koni Muluwa, 2010) | |
| Mpiin (B863) | RDC | musé | (Koni Muluwa, 2010) |
| Ngong (B864) | RDC | (Koni Muluwa, 2010) | |
| Mbuun (B87) | RDC | (Koni Muluwa, 2010) | |
| Yombe (H16c) | RDC | sééngá | (De Grauwe, 2009) |
| Ntandu (H16g) | RDC | nseenga | (Daeleman and Pauwels, 1983) |
| Hungan (H42) | RDC | músengi | (Koni Muluwa, 2010) |
| Kongo (H16a) | Angola | musenga | (Gossweiler, 1953) |
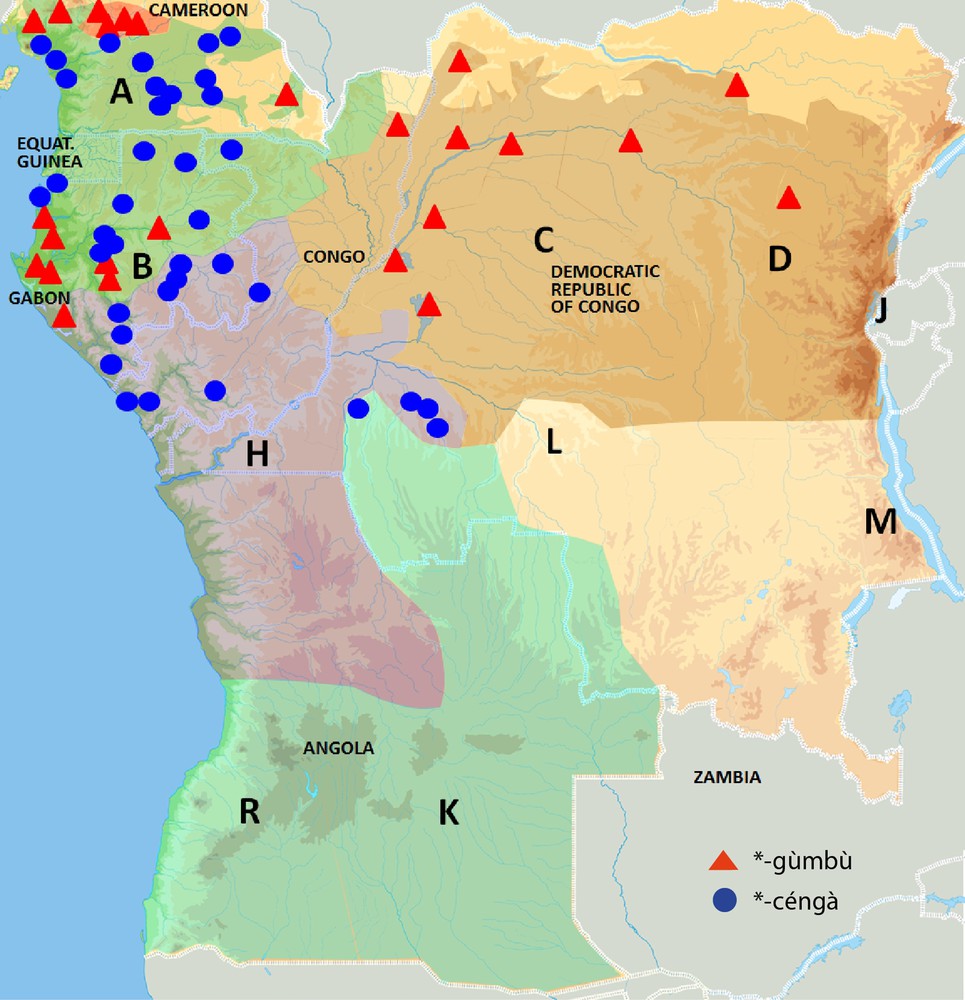
Geographical distribution of common Bantu names for Musanga cecropioides.
The second term can be reconstructed as *-gùmbù. It is realized as the noun stem -kombo in most current-day Bantu languages, as can be observed in Tables 3 and 4. Attestations of this term have been found in about 30 Bantu languages from four different countries: Cameroon, Gabon, Congo, and Democratic Republic of the Congo. This largely corresponds to the countries where *-céngà is attested. Nevertheless, both terms have a geographic distribution, which is almost perfectly complementary. The only language where both terms co-occur according to the data of Raponda-Walker and Sillans (1961) is Tsogo in Gabon, but the *-gùmbù attestation is phonologically irregular there and thus almost surely a loanword, possibly from the Myene languages spoken in the Ogooué-Maritime and Middle Ogooué provinces, and the coastal Lambaréné area in Gabon where *-gùmbù is regularly attested. Unlike in Tsogo, *-gùmbù attestations do manifest regular sound correspondences in most other languages. This indicates again that we are dealing, as in the case of *-céngà, with regularly inherited words whose geographic distribution is the result of language expansion. However, with respect to the internal classification of Bantu languages, *-gùmbù is distributed over more major historical sub-groups than *-céngà, i.e. North-West Bantu and Mbam-Bubi Bantu in western Central Africa (Table 3) and Congo River Bantu and Boan-Lebonya Bantu north of the Congo River (Table 4). Historically speaking, this is very significant, because *-gùmbù is mainly represented in languages belonging to different primary offshoots of the Bantu family tree (Bastin et al., 1999; Grollemund, 2012; Vansina, 1995) and is thus probably older than *-céngà. Its geographical distribution actually points in the same direction. As can be seen in Fig. 1, *-gùmbù attestations do not have a continuous block distribution like their *-céngà equivalents. Their spreading is much more scattered over the Bantu domain. They rather occur in three geographically isolated zones. Apart from the Myene languages in Gabon, and some neighbouring languages, such as Apindji and Ngowe, it is attested in two other clusters. The first cluster is situated much further north in Cameroon, and includes both North-West and Mbam-Bubi Bantu languages. The second cluster is situated much more easterly, mainly north of the Congo River, but also in some languages spoken in the proximity of its left bank. It includes languages belonging to the Congo River and Boan/Lebonya Bantu sub-groups. Unlike its synonym *-céngà, the noun stem *-gùmbù thus has a quite discontinuous geographic distribution. In historical-comparative linguistics, language features manifesting a scattered distribution are commonly interpreted as being old or certainly older than those having a more continuous distribution. This is certainly so if the block distribution of one lexical item, in this case *-céngà, crosscuts the more fragmented spreading of another with same meaning, here *-gùmbù. This indicates that *-céngà is a later innovation with regard to *-gùmbù. This greater antiquity of *-gùmbù seems to be borne out by the fact that possible cognates of this noun stem occur in related non-Bantu languages belonging to different Benue-Congo branches, such as bok(u)obe in Bokyi, a Cross-River language from Nigeria, and egum-amfuk in Ejagham, an Ekoid language from Nigeria (Burkill, 1985). The internal Bantu distribution of *-gùmbù over several primary Bantu branches combined with these probable non-Bantu Benue-Congo cognates allows reconstructing *-gùmbù into Proto-Bantu. No cognates of *-céngà have been found beyond Bantu. This, together with its geographically continuous distribution over only two historical Bantu sub-groups, confirms that it is a post-Proto-Bantu innovation. The fact that the term *-gùmbù can be reconstructed for M. cecropioides in Proto-Bantu indicates that the linguistic ancestors of current-day Bantu speakers were familiar with this species before the start of the Bantu Expansion. Why then did early Bantu speakers coin a new term for this pioneer colonizer once the Bantu Expansion had started? This question becomes all the more intriguing, if one reckons that Bantu vocabulary for other pioneer species also has two or more common Bantu terms, whose geographic distribution patterns display striking similarities with those of the two M. cecropioides terms. This is especially the case for vernacular Bantu names referring to E. guineensis and C. schweinfurthii, which were studied in more detail elsewhere (Bostoen, 2005; Bostoen, forthcoming).
Vernacular Bantu names for Musanga cecropioides. Attestations of *-gùmbù in North-West (3a) and Mbam-Bubi (3b) Bantu languages.
| Language | Country | Term | Source | |
| a | Oroko (A101) | Cameroon | ekombo | (Thomas et al., 1989) |
| Kundu (A12) | Cameroon | bokombo | (Burkill, 1985) | |
| Akoose (A15) | Cameroon | ekombo | (Burkill, 1985) | |
| Babinga (A80) | Cameroon | kombo | (Vivien and Faure, 1985) | |
| Mpiemo (A86) | Cameroon | kombo | (Thornell, 2004) | |
| Mpongwe (B11a) | Gabon | nkombo-gombo | (Raponda-Walker and Sillans, 1995) | |
| Orungu (B11b) | Gabon | nkombo-gombo | (Raponda-Walker and Sillans, 1995) | |
| Galwa (B11c) | Gabon | nkombo-gombo | (Raponda-Walker and Sillans, 1995) | |
| Nkomi (B11e) | Gabon | nkombo-gombo | (Raponda-Walker and Sillans, 1995) | |
| Apindji (B304) | Gabon | mogombo | (Raponda-Walker and Sillans, 1995) | |
| Tsogo (B31) | Gabon | mogombo | (Raponda-Walker and Sillans, 1995) | |
| Ngowe (B404) | Gabon | kombo-gombo | (Raponda-Walker and Sillans, 1995) | |
| Okande (B32) | Gabon | mókòmbò | (Grollemund, 2006) | |
| b | Tunen (A44) | Cameroon | bòkomba; kòmbombaŋ | (Dugast, 1967; Burkill, 1985) |
Attestations of *-gùmbù in Congo River (4a) and Lebonya/Boan (4b) Bantu languages.
| Language | Country | Term | Source | |
| a | Aka (C104) | CAR | èkòmbò | (Motte, 1982) |
| Akwa (C22) | Congo | ikombo | (Adjanohoun et al., 1988) | |
| Bobangi (C32) | RDC | mókòmbò | (Whitehead, 1899) | |
| Ntomba (C35a) | RDC | bokombo | (Mamet, 1955) | |
| Bolia (C35b) | RDC | bokómbo | (Ngila, 2000) | |
| Lingala (C36d) | RDC | kombókombó | (Dzokanga, 1979) | |
| Boloki (C36e) | RDC | bonkombo | (Hulstaert, 1966) | |
| Ngombe (C41) | RDC | gombo | (Rood, 1958) | |
| Binza (C41D) | RDC | ngombo | (Jean-Pierre Donzo, pers. comm.) | |
| Bwela (C42) | RDC | ikombo | (Jean-Pierre Donzo, pers. comm.) | |
| Turumbu (C54) | RDC | bokombo | (Termote et al., 2010) | |
| b | Pagibete (C401) | RDC | gombo | (Gillardin, 1959) |
| Baati (C43) | RDC | kombo | (Gillardin, 1959) | |
| Boa (C44) | RDC | kombo | (Jean-Pierre Donzo, pers. comm.) | |
| Mbole (D11) | RDC | bokombo | (Gillardin, 1959) | |
| Mbuti (D32) | RDC | kombo | (Terashima, 2003) |
3.2 Elaeis guineensis
As discussed above, one of the oil palm terms reconstructed in Proto-Bantu is *-téndé. It originally referred to a young immature oil palm. Two other common Bantu names for oil palm are *-bá and *-bídà, whose geographic distribution is represented in Fig. 2. Being a species whose economic exploitation is much more important than that of M. cecropioides, vernacular oil palm names are much easier to find in the literature, which explains to a great extent the higher number of attestations. Furthermore, the distribution of this species itself is wider than that of the umbrella tree. Although the geographic distribution of *-bá and *-bídà is not exactly the same as for *-céngà and *-gùmbù, there are striking correspondences. First of all, *-bá and *-bídà are also in complementary distribution. Secondly, the geographical spreading of *-bá is, just like that of *-céngà, much more clustered than that of *-bídà and crosscuts somehow that of *-bídà. The spreading of *-bídà is, just like that of *-gùmbù, much more fragmented and it occurs in the same historical Bantu sub-groups as *-gùmbù, i.e. Mbam-Bubi Bantu and North-West Bantu, including the peripheral Gabonese Myene languages in the west, as well as Congo River Bantu and Boan/Lebonya Bantu, further to the east, predominantly spoken north of and along the Congo River. Attestations of *-bá and *-céngà do not occur in exactly the same historical Bantu sub-groups. Both occur in West-Coastal Bantu languages, but *-bá does not occur in North-West Bantu, where *-bídà is predominant. It does occur, however, in the Inner Congo Basin Bantu languages, where we could not retrieve vernacular M. cecropioides names, due to the species’ absence in the dense forest. The third and probably most important correspondence is that *-bídà, just like *-gùmbù, can be reconstructed in Proto-Bantu and even beyond. It is not only representatively distributed among historical Bantu sub-groups, it also occurs in many related non-Bantu languages, spoken in Cameroon and Nigeria. On the contrary, *-bá does not occur beyond Bantu and is restricted to only two historical sub-groups. Just like *-céngà, it is to be considered a post-Proto-Bantu innovation. So, also in this case, early Bantu speakers coined a new term for the oil palm once the Bantu Expansion had started, while their ancestors where already familiar with this pioneer species before the start of the Bantu Expansion. Palynological studies indicate that the oil palm's presence in the natural vegetation of some parts of Africa is extremely ancient (Maley, 1999; Maley and Chepstow-Lusty, 2001; Sowunmi, 1999). Archaeological evidence confirms that its exploitation by Central Africans must indeed predate the beginning of the Bantu Expansion (D’Andrea et al., 2006; de Maret, 1994–1995; Lavachery, 2001; Neumann et al., 2012).
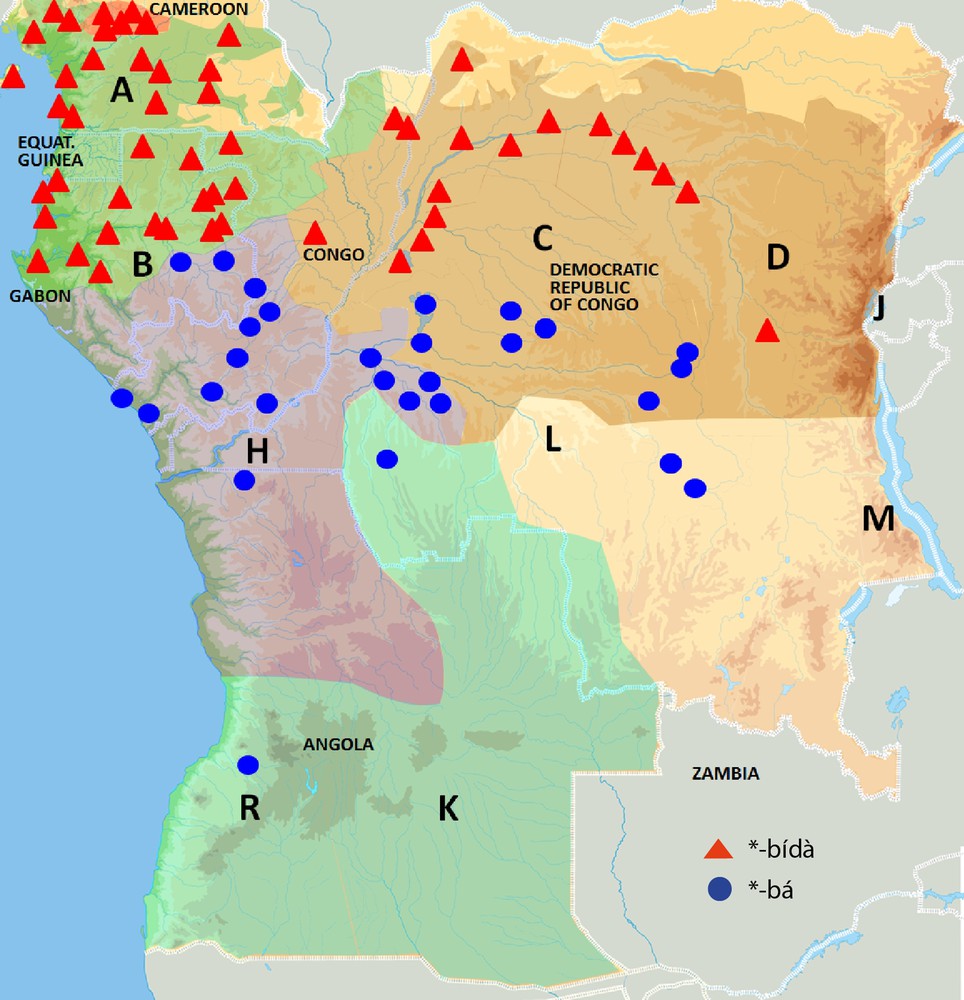
Geographical distribution of common Bantu names for Elaeis guineensis.
3.3 Canarium schweinfurthii
C. schweinfurthii is another light-demanding useful tree thriving in pioneer forests. Contrary to M. cecropioides, it is not a short-lived pioneer and it also has a wider ecological spectrum. It thrives in both humid and dryer dense forest types and can grow as high as 1600 m, while both other pioneer species discussed here are limited to 1200 m. C. schweinfurthii is found throughout tropical Africa in rainforest, gallery forest and transitional forest from Senegal to Cameroon and extending to Ethiopia, Tanzania and Angola (Burkill, 1985; Hutchinson and Dalziel, 1954; Orwa et al., 2009). It has a wider distribution than the umbrella tree and the oil palm, but its economic importance is lower that that of the oil palm. As a consequence, its Bantu names are less well documented. Similarly to the Bantu terms for the oil palm and umbrella tree, there are also two recurrent nouns for C. schweinfurthii across Bantu languages, suggesting that the economic importance of the tree is old. Once more, both nouns have an almost entirely complementary distribution and one term is considerably older than the other. The most archaic term is reconstructed as *-pátù in Proto-Bantu (Bostoen, forthcoming). As can be seen in Fig. 3, it has the most scattered distribution among current-day Bantu languages, as is the case for the most archaic umbrella tree and oil palm terms. Present-day forms of this noun occur in North-West Bantu, in East Bantu languages further east and in two different West-Bantu groups further south, i.e. West-Coastal and South-West Bantu. Attestations of this Proto-Bantu name for C. schweinfurthii were also found outside Bantu. Related names were reported in a number of Central Nigerian languages, belonging to the Plateau sub-group of the East-Benue-Congo group (Blench, 2006: 117). Just like the most archaic Bantu names for the other two pioneer species, *-pátù thus has a time depth that is considerably greater than Proto-Bantu. The second, more recent C. schweinfurthii name can be reconstructed as *-bídí and has no attestations beyond Bantu. Moreover, its present-day forms have a much more contiguous distribution within Bantu. They are limited to North-West-Bantu and two distinct West-Bantu groups, i.e. West-Coastal Bantu and Congo Basin Bantu. This corresponds largely to the distribution of the more recent term for M. cecropioides, i.e. *-céngà, except that the latter species does not occur in the Inner Congo Basin.
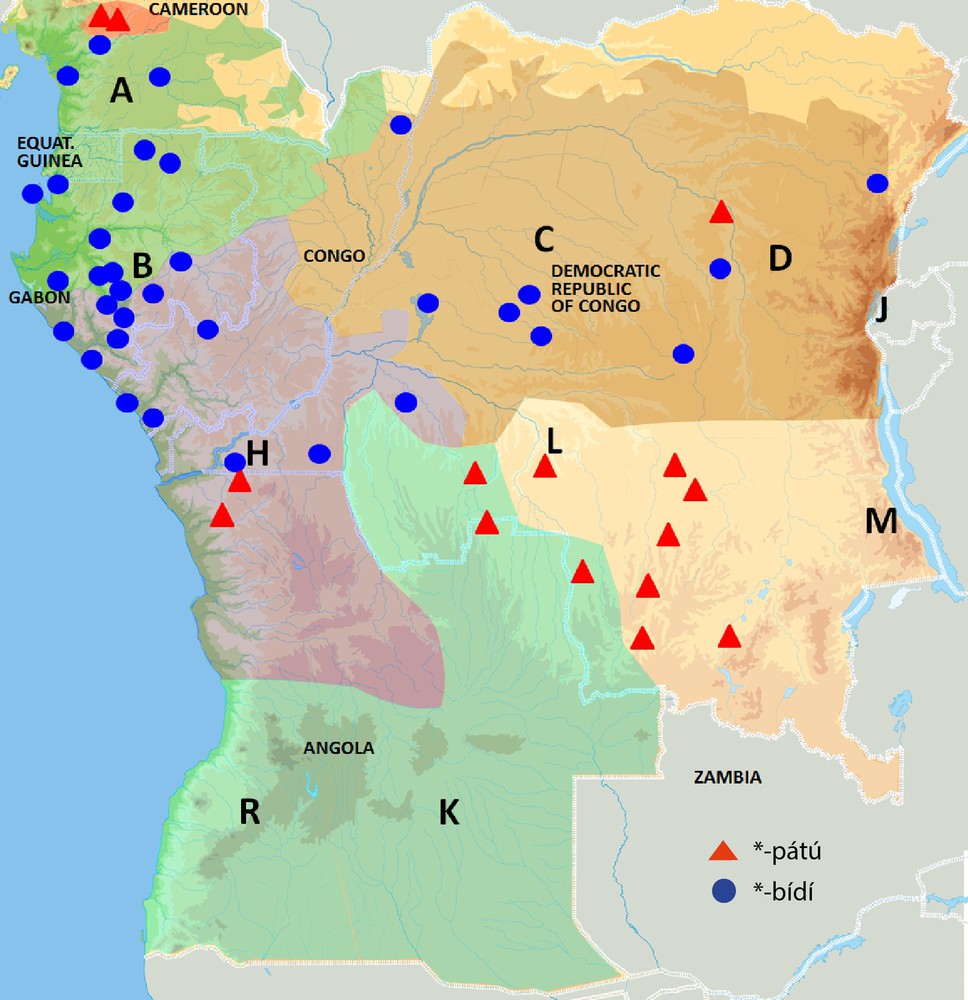
Geographical distribution of common Bantu names for Canarium schweinfurthii.
3.4 Mapping on the phylogenetic Bantu tree
The pioneer species M. cecropioides, E. guineensis and C. schweinfurthii all have two common Bantu names of which one has a more scattered distribution among present-day Bantu languages and can be reconstructed to Proto-Bantu and far beyond and another that has a much more contiguous geographic distribution and a much more shallow time depth, i.e. posterior to Proto-Bantu. In spite of these overall correspondences, they also differ in one important respect, namely their mapping on the phylogenetic tree of the Bantu languages. According to the most recent phylogenetic classification of north-western Bantu languages (Grollemund, 2012), as shown in Figs. 4–6, the Cameroonian Mbam-Bubi languages constitute a primary branch followed by the branching of two parallel clades. The first one is the so-called North-West Bantu branch (Vansina, 1995), consisting of the Cameroonian and Gabonese languages of Guthrie's zone A and groups B10–30.2 The second one is the so-called Narrow West Bantu branch (Vansina, 1995), consisting of the languages of Guthrie's groups B40–80 and zones C and H, spoken further south and east in Gabon and the two Congos. In the phylogenetic classification of Grollemund (2012), the latter branch also includes the so-called ‘Boan/Lebonya’ languages of Guthrie's groups D10–20 (Vansina, 1995).
As shown in Fig. 4, the introduction of the innovative *-bá term for E. guineensis happened at a well-defined node of the phylogenetic Bantu tree, i.e. at the level of the most recent common ancestor of the so-called ‘West-Coastal’ languages of Guthrie's zone H and groups B40–80 and the so-called ‘Inner Congo Basin’ languages of Guthrie's groups C60–80 (Vansina, 1995). This means that the innovative term *-bá for oil palm was coined by the speakers of this specific intermediate ancestor language to the detriment of the more archaic term *-bídà, which they had lost by that time. The introduction of *-bá thus corresponds to a clearly identifiable stage in the Bantu Expansion. It happened before the break-up of the most recent common ancestor of the ‘West-Coastal’ and ‘Inner Congo Basin’ Bantu languages. Taking into account the current-day geographic distribution of these languages and Ockham's razor principle of the least moves, the cradle of this branch can be placed, following Vansina (1995: 187), ‘somewhere in northern Congo’.
As shown in Fig. 5, the introduction of the innovative *-bídí term for C. schweinfurthii also happened at a well-defined node of the phylogenetic Bantu tree, though at a node superior to the one where *-bá for E. guineensis was introduced. It turns up at the stage of the most recent common ancestor of the so-called North-West and Narrow West Bantu branches after its split from the Mbam-Bubi branch. The new term must have replaced the archaic term *-pátù, but not entirely, since it still occurs in some peripheral Narrow West-Bantu languages of the West-Coastal and South-West sub-branches. Just like *-bá, *-bídí was introduced at a clearly identifiable stage in the Bantu Expansion, but earlier and further north. Applying again the least moves principle, the separation of the most common recent ancestor of the North-West and Narrow West Bantu languages from the Mbam-Bubi branch must have happened somewhere in the wider vicinity of current-day Yaoundé.
The complementary distribution of the M. cecropioides terms *-céngà and *-gùmbù does not fit so well with the phylogenetic Bantu tree, as shown in Fig. 6. Just like the archaic terms for E. guineensis and C. schweinfurthii, *-gùmbù is also represented in the primary Mbam-Bubi branch. Just like the oldest oil palm term, it was also maintained in the sub-group of B10–B30 languages spoken in northern Gabon, and in the sub-group of C10–30, C50–D10–20 languages predominantly spoken north of the Congo River. However, in contrast to the innovative terms for the other two pioneer species, *-céngà does not seem to have been introduced at a clearly identifiable node of the phylogenetic Bantu tree. It certainly happened earlier than the introduction of the oil palm term *-bá, since *-céngà is also present in certain North-West Bantu languages. It must have happened after the separation of the Mbam-Bubi languages, but later than the introduction of the C. schweinfurthii term *-bídí, since not all North-West and Narrow West Bantu sub-branches were affected by this lexical innovation. It is historically significant that the most archaic term for M. cecropioides mainly survives in the primary sub-branches of both North-West Bantu and Narrow West Bantu. These are also geographically outliers, i.e. the B10–30 languages of lower Ogooué area in northern Gabon and the C10–50-D10–20 Bantu languages spoken north of the Congo River in the northern part of both Congos. It is likely that these languages were geographically more rapidly separated from their closest relatives. The remainder of both North-West and Narrow West Bantu languages may have remained in communication with each other much longer (Vansina, 1995: 186). This explains how the new term *-céngà could make its way in sub-groups of both branches. The early contact zone between North-West and Narrow West Bantu languages must have been somewhere in southern Cameroon, ‘in an area closer to the North-West cluster and the Atlantic Ocean’ (Vansina, 1995: 186). This is in between the wider Yaoundé area, where Mbam-Bubi languages broke away from the rest, and northern Congo, where the cradle of Narrow West Bantu languages is situated.
4 Discussion
The possibility of reconstructing a vernacular name for M. cecropioides in Proto-Bantu indicates that the linguistic ancestors of current-day Bantu speakers were familiar with this pioneer species before the beginning of the Bantu Expansion and that they must have had access to it in their homeland. Consequently, they must have lived in a tropical forest zone with enough large clearings for this short-lived, light-demanding pioneer to thrive. The situation of the Bantu homeland in a forest-savannah mosaic area is in line with earlier Proto-Bantu reconstructions for tree species such as E. guineensis, C. schweinfurthii, Cola spp., and Parinari curatellifolia (Bostoen, 2005; Bostoen, forthcoming) as well as with archaeological evidence (Lavachery, 2001). The reconstruction of Proto-Bantu terms for both C. schweinfurthii and E. guineensis is in line with findings at Shum Laka in the Cameroonian Grassfields region, the most important archaeological site associated with the Bantu homeland (Lavachery, 2001) and later Ceramic Late Stone Age sites further south in West Central Africa, generally associated with the Bantu Expansion, where C. schweinfurthii remains are often found in association with those of E. guineensis (D’Andrea et al., 2006; de Maret, 1994–1995; Neumann et al., 2012). These Proto-Bantu tree names also suggest that the economic exploitation of these useful trees in their natural environment is old and at least as important as the cultivation of domesticated plants in the mixed subsistence economies of early Bantu speech communities.
If Bantu speakers were already familiar with M. cecropioides, E. guineensis, and C. schweinfurthii before they started to immigrate into Central Africa, it is at least remarkable that they coined new terms for each of these pioneer species in the course of their expansion. Given that language change is often the outcome of changes in the outside world, these lexical innovations may reflect changes in the natural environment in which early Bantu speakers lived. It could imply that at some point, these pioneer species became less abundant or even disappeared completely from their habitat and that Bantu speakers had to reinvent words when those became more plentiful again or reappeared at some later stage. This is difficult to imagine, except if one assumes that Bantu speakers started to live in forest environments where these trees no longer thrived. Certain early Bantu speech communities, once they started to move south, probably had to adapt to living in a dense forest environment where they no longer had access to certain tree species typical of their ancestral forest-savannah mosaic habitat, such as M. cecropioides, E. guineensis and C. schweinfurthii. The coining of new names may have happened then when these pioneer species re-emerged in the environment of Bantu speakers. This occurred at clearly distinct stages for the three species, since the innovative term for each of them turns up at a successive position in the phylogenetic Bantu tree: firstly C. schweinfurthii, subsequently M. cecropioides, and lastly E. guineensis. It could be assumed that these pioneer species reappeared in the habitat of Bantu speakers, because the latter migrated to regions where those trees thrived again, i.e. south or east of the rainforest. However, this is rather unlikely, if one reckons that the respective ancestral Bantu languages, in which the new terms appeared, were spoken somewhere in between the area south of Yaoundé and Douala and northern Congo and Gabon. It seems more plausible to assume that these pioneer species re-emerged or became abundant again in the life of Bantu speakers, because their forest environment was seriously disturbed, for instance due to climate change. In this regard, it is quite alluring to link the appearance of innovative Bantu vocabulary for pioneer colonizers to the climate-induced development of forest-savannah mosaics and young forests that replaced major parts of the rainforest in the course of the first millennium BC. The three pioneer species discussed here are represented in Central African pollen record between 2500 and 2000 BP (Elenga et al., 2000; Maley and Brenac, 1998; Ngomanda et al., 2009; Vincens et al., 1994, 1998). Moreover, as mentioned above, E. guineensis and C. schweinfurthii are well attested in archaeological sites from that same era (de Maret, 1994–1995; Neumann et al., 2012). Moreover, the area where the ancestral languages, in which the new Bantu names for these pioneer species emerged, were spoken is within the radius of the so-called ‘Sangha Interval’, the savannah and open forest mosaic corridor in the western part of the Congo basin that emerged around 2500–2000 BP as a result of the climate-induced disturbance of the Central African rainforest (Doumenge et al., 2012). The fact that the vocabulary for the three pioneer species did not innovate at the same pace could indicate that changing climate conditions affected them differently. In this respect, one also needs to take into account, as discussed above, that their ecological spectrum is not the same. The environment types where C. schweinfurthii thrives are much wider than those of E. guineensis and M. cecropioides, which often grow together in the same disturbed environments. Moreover, the three species are not equally important from an economic point of view. The oil palm was no doubt the most useful in the subsistence economy of early Bantu speech communities. This may explain why its terminology was longer conserved in Bantu languages, even when the species became less abundant in the habitat of their speakers. C. schweinfurthii may have suffered competition from other fruit yielding species, especially Dacryodes edulis. In popular taxonomy, C. schweinfurthii is commonly considered to be a wild variety of D. edulis, which provides similar, but bigger fruits. Linguistic evidence suggests that the latter tree was also exploited by early Bantu speech communities (Bostoen, forthcoming).
Finally, it is historically meaningful that some Bantu languages simply retained the Proto-Bantu vocabulary for the pioneer species. This could indicate that those trees never disappeared from their ancestor's habitat and retained their economical importance. This is especially the case for the Mbam-Bubi Bantu languages of Central Cameroon, which retained the Proto-Bantu terms of all three species. It is also remarkable that the B10–30 Bantu languages from the lower Ogooué area in northern Gabon and the C10–50–D10–20 Bantu languages spoken north of the Congo River innovated the term for C. schweinfurthii, but conserved the Proto-Bantu names for oil palm and umbrella tree. These language groups constitute both primary sub-branches of respectively North-West and Narrow West Bantu, two parallel branches. Their ancestors may thus have lived at some point in a forest environment where they no longer had access to C. schweinfurthii or no longer use it, but where they continued to exploit M. cecropioides and E. guineensis. In contrast to their closest relatives from the North-West and Narrow West Bantu, they kept in touch with the last two species while migrating respectively south to the lower Ogooué area and east to the area north of the Congo River. Consequently, the ancestors of these two sub-branches must have broken away from their closest relatives before those started to settle in a forest environment where the oil palm and umbrella tree disappeared or lost economic importance. They must have migrated themselves, respectively southwards and eastwards, through forest-savannah mosaics where both M. cecropioides and E. guineensis still thrived.
In conclusion, the linguistic data discussed in this article provide circumstantial evidence adding to the hypothesis that migrating early Bantu speech communities confronted climate-induced changes of the natural environments in which they lived and that these vegetation dynamics influenced their migration routes. It is obvious that the wider historical implications of these linguistic data can only be fully assessed if they are considered in a wider interdisciplinary perspective. They need to be confronted with evidence from other disciplines, especially archaeology, palynology, and biogeography.
Acknowledgements
The authors’ thanks go to Jean Maley and Charles Doumenge for their very useful comments on an earlier version of this article. The usual disclaimers apply. Koen Bostoen's research is funded by the Special Research Fund of Ghent University and his research group by a Starting Grant of the European Research Council (KongoKing, Grant No. 284126). Rebecca Grollemund's research is also supported by the European Research Council (Mother Tongue, Grant No. 268744) and the ANR ‘CLHASS’. Joseph Koni Muluwa's research was financed by the Belgian Science Policy, while he was employed at the Linguistics Service of the Royal Museum for Central Africa in Tervuren, Belgium (2011–2012).
1 The starred forms in this article are lexical reconstructions. These are etymons or earlier forms of a given word in an ancestor language, in this case Proto-Bantu or a subsequent proto-language, reconstructed on the basis of a comparative series of attestations of this word in current-day daughter languages.
2 Guthrie (1971) used a number of typological and geographical criteria to classify the Bantu languages in a number of zones indicated by a capital letter (zones A–S) further subdivided in groups indicated by a decimal cipher (10–90). This referential, ahistorical classification is still used by comparative Bantu linguists today because it facilitates the geo-location of the Bantu languages. Codes, such as A22, A24, etc., following language names in Tables 1–4, refer to Guthrie's classification.