

Abridged English version
1 Introduction
During Late Oligocene–Miocene, an Avicennia mangrove developed around the Mediterranean realm in southern France and northern Spain [4,6,29]. It reached its maximum development during the Late Burdigalian-Langhian, coeval with that of coral reefs [7,11]. This mangrove constitutes a palaeoclimatic as well as stratigraphic indicator for Middle Miocene times.
Here we present a palynological investigation of the upper part of the Middle Miocene series of the Châteauredon area (southeastern France).
2 Geological setting
The Châteauredon series (∼ 44°N; 006°13′E) are located at the Digne thrust front (Fig. 1A and B). They are dominantly composed of deltaic sediments including some marine deposits and some continental episodes (Fig. 1C). Sampling has been carried out in the black levels present in the upper part of the series. Dating of this interval is based on micromammal fossil sites, palynological data from a previous study [9], and correlations of C values between marine and terrestrial material [8,9,24]. The stratigraphic interval extends from Upper Burdigalian to Serravallian.
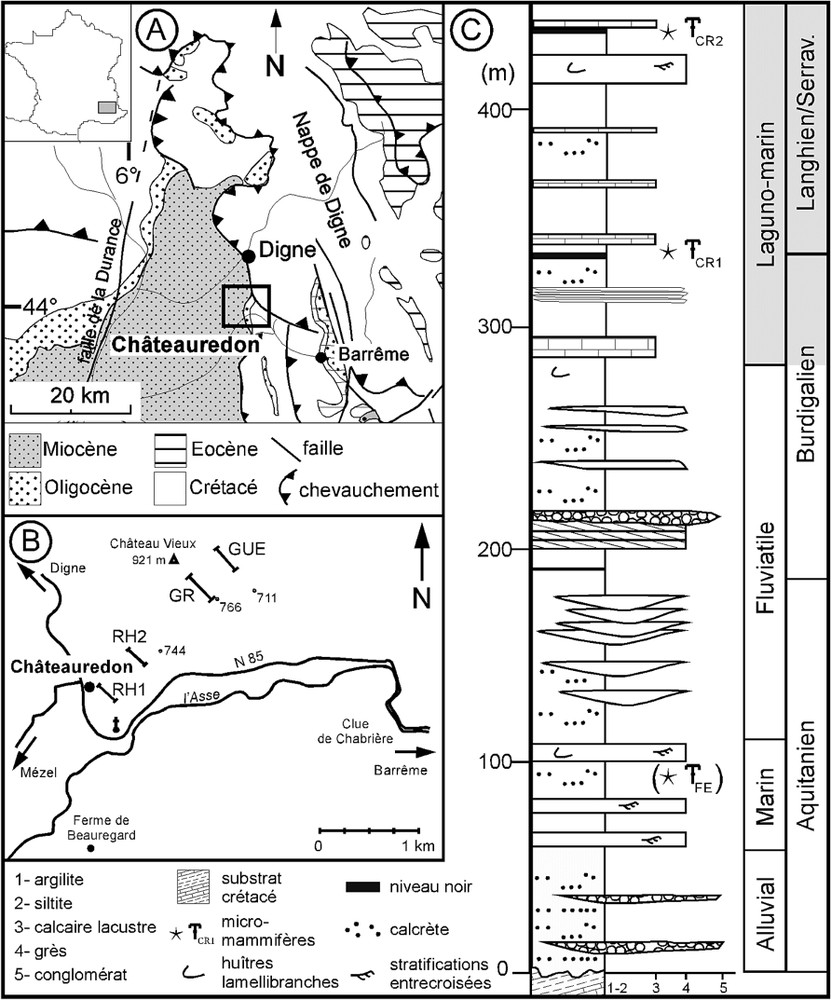
(A) Localisation de la zone d'étude dans le bassin de Digne–Valensole. (B) Localisation des coupes étudiées autour de Châteauredon. (C) Coupe schématique de la série miocène de Châteauredon [24].
(A) Location of the studied area in the Digne–Valensole Basin. (B) Location of the studied sections near Châteauredon. (C) Schematic log of the Miocene series of Châteauredon [24].
3 Palynological study
Three sections that present similar lithological successions were sampled (RH1, RH2 and GUE, Fig. 1B). They were correlated on the basis of the facies succession. Three samples out of the 15 collected contain Avicennia pollen grains. Two of them (RHCR1, GUE01) correspond to the level of the micromammal fossil site T̶CR1 [23], while the third one (RH01) was sampled 4 m below this level. All data presented in the following refer to these three samples.
3.1 Description of the Avicennia pollen genus
The pollen grains attributed to the genus Avicennia are tricolporate, isopolar in shape, and circular to lobate in polar view, elliptic in equatorial view. Their size varies from 18 to 25 μm. The aperture is elongate and pores are often rounded. The exine with two distinct layers consists of a regular endexine (0.5- to 1-μm thick) and a reticulate ectexine (1- to 1.5-μm thick) with regular network, psilate tectum and distinct columella. These grains are close to those of the modern species A. marina and alba (Fig. 2).
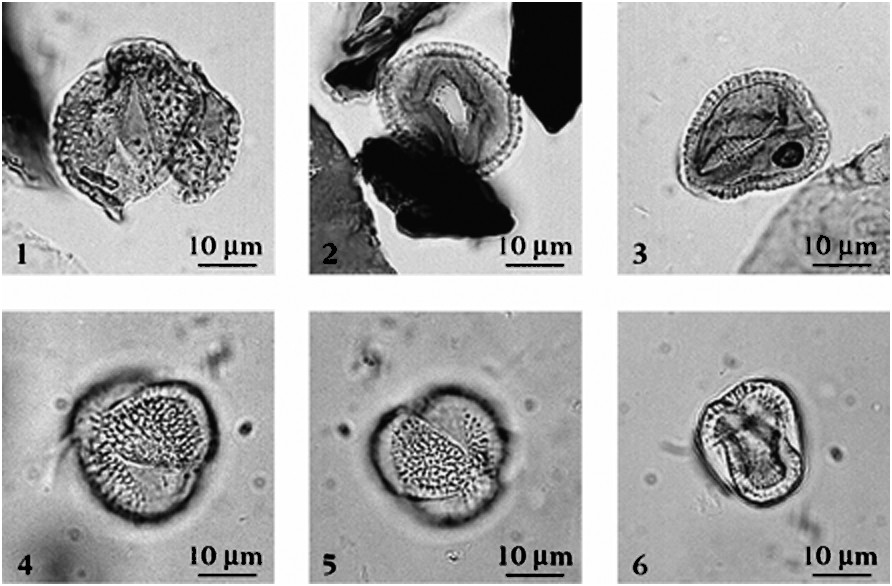
Grains de pollen d'Avicennia. 1–3 : Échantillon GUE01 ; 4–6 : Avicennia alba actuel.
Avicennia pollen grains. 1–3: Sample GUE01; 4–6: modern Avicennia alba.
3.2 Spores and pollens associations
The relative proportions of elements show that dinocysts represent 30 to 50% of the identified grains and that the Avicennia pollen grains correspond to 4 to 12% of the total pollen grains (except for RH01 where both are <1%) (Fig. 3).
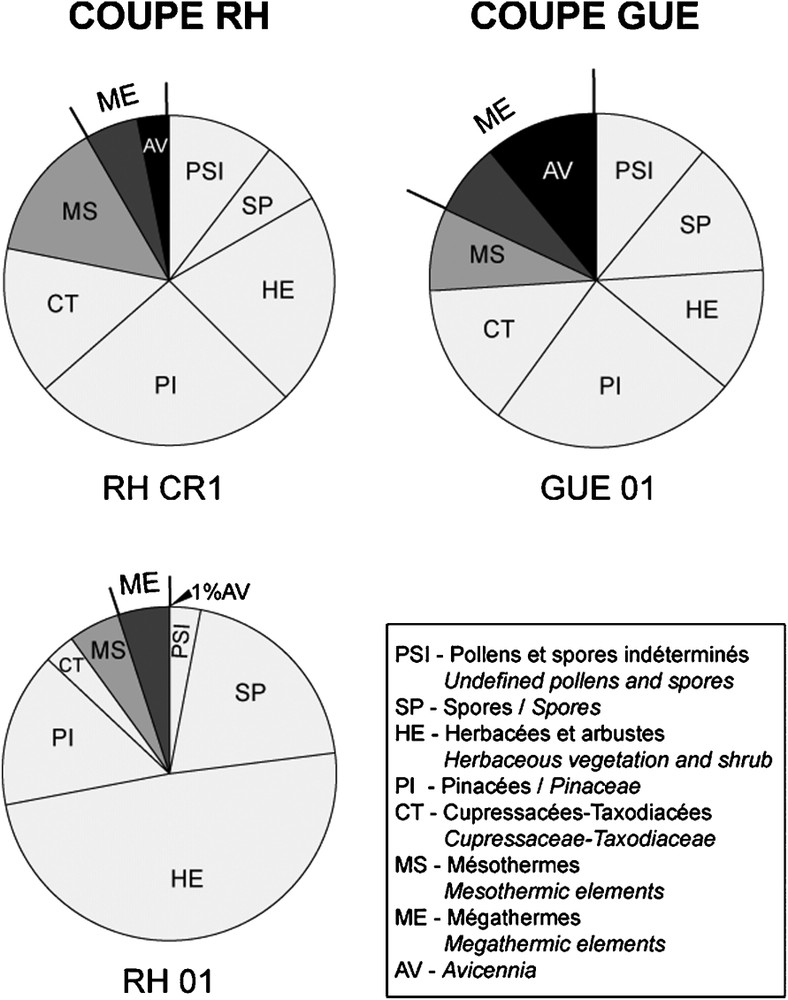
Associations sporo-polliniques des trois échantillons contenant des grains de pollen d'Avicennia.
Spore and pollen distribution in the three samples containing Avicennia pollen grains.
The spore and pollen assemblages cover a wide range of vegetation types from the low vegetation strata (mangrove, littoral fringe and coastal swamps) with quite open landscapes (herbaceous communities), to the mixed evergreen–deciduous forest and mid-altitude conifer forest, the later with very low proportion of microthermic elements such as Abies and Picea. Spores and pollen grains can be grouped in seven classes that are helpful for palaeoenvironmental and climatic interpretations (Fig. 3): (1) ferns and bryophytes spores, (2) non-identified spores and pollen grains, (3) herbaceous and bush vegetation (HE), (4) mangrove and coastal swamp forests (CT, AV). The forest vegetation is composed of three assemblages: (5) coniferous forest (PI), (6) mesothermic taxa (MS), and (7) megathermic taxa, also including AV.
Elements are well distributed in the pollen diagrams of the GUE01 et RHCR1 samples while herbaceous pollen grains, Pinaceae and spores dominate the RH01 sample. Based on the pollen diagram and the low percentages of dinocysts and Avicennia in RH01, we interpret this site as a back-mangrove with more fluvial input.
4 Discussion
4.1 Dating the ‘Avicennia peak’
The stratigraphic scheme of the Châteauredon series is based on an integrated stratigraphy combining micromammal fossil sites, palynology and chemostratigraphy (Fig. 4A) [8,9]. All of these independent stratigraphic methods lead us to propose a numerical age between 15.8 and 16.5 Ma for the ‘Avicennia peak’, according to the Berggren et al. timescale [3] (Fig. 4A and B).
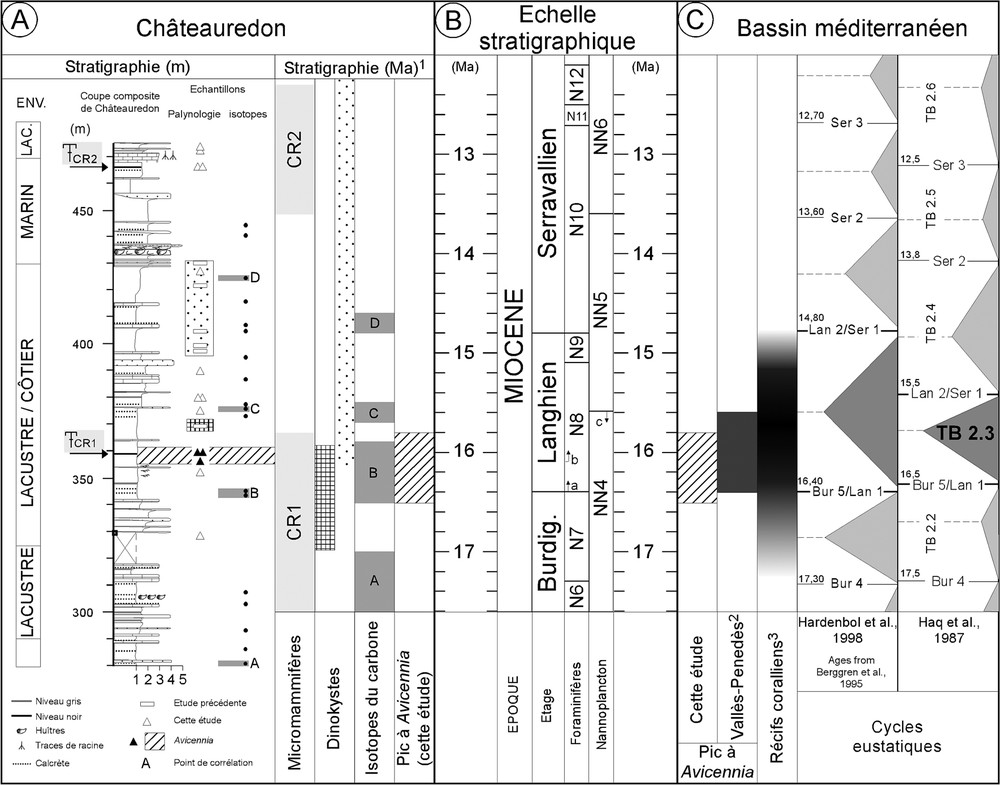
Le « pic d'Avicennia ». (A) Coupe lithologique et stratigraphie intégrée de la partie supérieure de la coupe de Châteauredon (cf. Fig. 1). Les échantillons ayant livré Avicennia sont indiqués par des triangles noirs ; les deux échantillons supérieurs sont RHCR1 et GUE01, surmontant RH01. 1 Stratigraphie de Châteauredon [8,9]. (B) Échelle stratigraphique [3] : (a) FAD de G. sicanus, (b) FAD de P. glomerosa, (c) LAD de H. ampliaperta. (C) Données stratigraphiques sur le bassin méditerranéen. 2 [7], 3 synthèse in [16].
The ‘Avicennia peak’. (A) Integrated stratigraphy of the upper part of the Châteauredon series (cf. Fig. 1). Samples containing Avicennia are in black triangles; RHCR1 and GUE01 overlay RH01. 1 Châteauredon stratigraphy [8,9]. (B) Middle Miocene timescale [3], (a) FAD of G. sicanus, (b) FAD of P. glomerosa, (c) LAD of H. ampliaperta). (C) Stratigraphic data for the Mediterranean Basin. 2 [7], 3 synthesis in [16].
The ‘Avicennia peak’ is associated with a high sea level, as indicated by the large number of dinocysts and the underlying tidal facies, in lagoonal marls with pedogenesis traces. This coeval high sea level could be attributed to the TB2.3 cycle [17], i.e. to the sequence limits Bur5/Lan1 and Lan2/Ser1 on the Berggren timescale [18] (Fig. 4C).
In the western Mediterranean realm, the Avicennia mangrove is known from Chattian to Upper Langhian [4,6], with some relics during the Messinian in Sicily [28] and Algeria [12]. Its maximum development, so-called ‘Avicennia peak’, corresponds to the Upper Burdigalian–Langhian (Rhone Valley to Catalonia) [6] and is restricted to N8–NN4 pro parte (Vallès–Penedès Basin, Spain) [7] (Fig. 4B and C). Our dating of the peak based on other stratigraphic tools confirms these results, indicating that the mangrove extension lasted less than 700 kyr (Fig. 4C). A coeval large extension of the corals is also well known all over the Mediterranean realm during Middle Miocene from N7 to N9, that is classically related to the Langhian high sea level [14,15] (synthesis in [16]) or to the cycle TB2.3 [10] of Haq et al. [17]. The occurrence of the Avicennia peak just above marine facies in the Châteauredon succession also supports a coeval maximum mangrove development with the Langhian high sea level.
4.2 Palaeoclimatic reconstruction
The ‘Avicennia peak’ and the large development of the coral reefs corresponded to a period of climatic optimum during Middle Miocene. This relatively warm period is characterized, in addition to the Avicennia pollen grains, by elements from megathermic tropical floras, now principally restricted to southern Asia, and that were present at that time in Spain [2,31], Portugal [1], Hungary [19,22], and Germany [30].
Estimations of average temperatures range from 20–23 °C in Portugal [1] to 18 °C for northwestern Mediterranean areas [5], 20 °C in Germany [30] and in the Pannonian Basin [22]. Temperature of the warmest months reached in average 28 °C, while those of the coldest months did not go below the 10 °C isotherm. Annual rainfall is estimated between 1300 [30] and 1400 mm [22]. These palaeo-temperatures and precipitations are consistent with present-day impoverished mangrove distribution [26].
Proportion of Avicennia pollen grains in the Châteauredon area (4 to 12% of the pollen amount) is higher than in Languedoc (<1%, except for the Montpeyroux section, 6 to 8%), but lower than those from Catalonia (15 to 20%) [6]. Discovery of a mangrove in the Digne area corresponds to the northernmost extension of such a vegetation, since previously study only placed the mangrove up to the southern Provence area, some 100 km to the south [6]. Palaeogeographic reconstructions place the Châteauredon area by 42°N latitude during Middle Miocene [13]. During the same period (biozone N8), a mangrove had also been identified at similar latitude in Japan [20,32] and in Hungary [25,27]. This extension of the Avicennia mangrove, nearly 20° to the north of modern systems [26], is a consequence of the global climatic dynamics: during the Middle Miocene, the absence of an Arctic ice cap [27] favoured lower climatic gradients, which enabled development of mangrove at more northerly latitudes.
5 Conclusion
The presence of Avicennia pollen grains in Châteauredon area is of both stratigraphic and palaeoclimatic interest. Using independent stratigraphic data, the ‘Avicennia peak’ at Châteauredon is given an age of 15.8–16.5 Ma, consistent with the biostratigraphic data of the Mediterranean Basin (N7/N9) and more particularly with the Vallès–Penedès Basin (N7/8–NN4 pro parte). Associated marine facies also coincides with the Langhian highstand (cycle TB2.3), which is also known to be coeval with Avicennia in the Vallès–Penedès Basin. Finally, this mangrove implies a (sub)tropical climate at a 40°N palaeolatitude. This could represent the northernmost extension of the mangrove in Neogene subalpine and Provence Basins.
1 Introduction
L'existence d'une mangrove à Avicennia au Miocène inférieur et moyen est connue sur le pourtour méditerranéen du Sud de la France et du Nord de l'Espagne [4,6,29]. Celle-ci semble avoir atteint son maximum d'extension au Burdigalien terminal–Langhien, en concomitance avec le développement de récifs coralliens [7,11]. Outre une information climatique précieuse, l'âge du maximum d'extension de cette mangrove et de la flore qui lui est associée constitue un apport stratigraphique intéressant.
Dans cette optique à la fois stratigraphique et paléoenvironnementale, une étude palynologique de la partie supérieure de la série miocène de Beynes–Châteauredon (Alpes-de-Haute-Provence) a été entreprise sur les niveaux riches en matière organique présents dans les dépôts laguno-marins.
2 Contexte géologique
La série de Beynes–Châteauredon (env. 44°N ; 006°13′E) est localisée dans le bassin de Digne–Valensole, au front de la nappe de Digne (Fig. 1A et B). Elle est composée de dépôts deltaïques avec quelques incursions marines marquées (faciès à tidalites, débris de balanes) et des épisodes plus continentaux (faciès de plaine d'inondation et de cônes alluviaux) (Fig. 1C). La partie supérieure de la série, dans laquelle l'échantillonnage a été effectué, correspond à des dépôts lagunaires et marins qui présentent quelques épisodes de pédogenèse. Cette unité, datée à l'aide de deux gisements de micromammifères, de données palynologiques précédemment acquises [9] et de corrélations entre les signaux du C marin (foraminifères) et continental (paléosols), s'étend du Burdigalien supérieur au Serravallien, toutes incertitudes retenues (Fig. 1C) [8,9,24].
3 Analyse palynologique
L'analyse palynologique s'appuie sur l'échantillonnage de trois coupes (RH1, RH2 et GUE, Fig. 1B). Ces coupes, qui présentent une succession lithologique très semblable, ont été corrélées sur le terrain grâce à des faciès repères. Les quinze échantillons prélevés sur ces coupes s'ajoutent aux six échantillons étudiés précédemment (coupe GR) [8,9].
Trois échantillons ont livré des grains de pollen du genre Avicennia, associés à une riche microflore pollinique ainsi qu'à des assemblages diversifiés de kystes de dinoflagellés. Ils appartiennent à deux coupes différentes (Fig. 1B) : coupe RH1 (échantillons RH01 et RHCR1) et coupe GUE (GUE01). Deux échantillons (RHCR1, GUE01) correspondent à un même niveau noir, ayant livré des restes de rongeurs (gisement « Châteauredon 1 » [23], T̶CR1, Fig. 1). Le troisième (RH01) a été prélevé 4 m en dessous de RHCR1, dans un autre horizon noir.
3.1 Description du pollen du genre Avicennia
Ces grains tricolporés de forme isopolaire, longiaxe et elliptique en vue méridienne, ont un contour circulaire à sub-triangulaire en vue polaire (Fig. 2). Leurs dimensions varient de 18 à 25 μm. Ils possèdent une ectoaperture large et granuleuse et une endoaperture avec pore de forme ronde à elliptique, à bords irréguliers. L'exine à deux couches distinctes est composée d'une endexine régulière, dont l'épaisseur varie de 0,5 à 1 μm et d'une ectexine réticulée à mailles irrégulières, de 1 à 1,5 μm, avec tectum lisse et columelles nettes.
Bien qu'il existe une littérature descriptive accessible [4], les formes rencontrées ont été comparées au pollen actuel d'Avicennia marina, alba et africana provenant de l'océan Indien et de l'océan Atlantique. Elles sont proches des espèces de la mangrove actuelle indo-ouest-pacifique à A. marina et alba.
3.2 Les associations sporo-polliniques
Les grains de pollen d'Avicennia sont associés, dans les échantillons de Beynes–Châteauredon, à un assemblage d'une trentaine d'espèces de kystes de dinoflagellés et à des associations sporo-polliniques variées, détaillées ci-après. Les comptages effectués sur les trois échantillons montrent que les kystes de dinoflagellés représentent 30 à 50% des grains identifiés, à l'exception de l'échantillon RH01, dans lequel ils ne dépassent pas 1%.
Les grains d'Avicennia, quant à eux, rapportés au total des grains de pollen, présentent des pourcentages de 4 à 12% pour les échantillons GUE01 et RHCR1, et moins de 1% pour l'échantillon RH01 (Fig. 3). Ces pourcentages, apparemment faibles, ne sont pas surprenants pour ce pollen, toujours sous-représenté.
Les associations sporo-polliniques vont des strates de végétation basses (mangrove, frange littorale et marécages côtiers) avec étendues herbacées plus ouvertes, à la forêt mixte sempervirente à éléments décidus, à la forêt de moyenne altitude à conifères, les genres « microthermes » telles qu'Abies ou Picea étant très peu représentés.
Les assemblages ont été groupés de façon à faciliter une interprétation climatique et environnementale. Les spores et les grains de pollen se répartissent en sept groupes (Fig. 3) :
- – les spores de fougères et de bryophytes (SP) ;
- – les spores et grains de pollen non identifiés (PSI) ;
- – la strate herbacée et buissonnante (HE), représentée par les Poaceae, Amarantaceae–Chenopodiaceae, Asteraceae, Sparganiaceae/Typhaceae, Liliaceae, Apiaceae et Plumbaginaceae ;
- – la mangrove et les marécages littoraux, représentés par les Verbenaceae (Avicennia, AV), Taxodiaceae-Cupressaceae (CT), Myricaceae, Cyrillaceae–Clethraceae, Nyssaceae, Symplocaceae et rares Sapotaceae.
La strate arbustive est composée de trois ensembles :
- – les Gymnospermes (PI), comprenant essentiellement des Pinaceae : Pinus, Cathaya, Tsuga, Abies, Picea, Cedrus, et de rares Podocarpaceae ;
- – les mésothermes (MS), avec les genres Fagus, Betula, Alnus, Tilia, Ilex, Ulmus, Carya, Pterocarya, Corylus, Carpinus, Juglans et quelques représentants des familles d'Eleagnaceae et Buxaceae ;
- – les mégathermes (ME), avec des Juglandaceae (Engelhardtia, Platycarya), Sapotaceae, Euphorbiaceae, Thymeleaceae, Arecaceae, Chloranthaceae, Malpighiaceae, Cycadaceae, Magnoliaceae, Hamamelidaceae (Corylopsis), Asclepiadaceae et Verbenaceae (Avicennia, AV). Les éléments de la flore méditerranéenne sont très rares (quelques grains de Vitaceae, Oleaceae et Berberidaceae).
Les diagrammes (Fig. 3) montrent un équilibre relatif de ces divers groupes pour les échantillons GUE01 et RHCR1, et une prédominance des herbacées, des pinacées et des spores pour l'échantillon RH01. La rareté des kystes de dinoflagellés et du pollen d'Avicennia dans ce dernier échantillon, nous conduit à interpréter l'environnement de dépôt comme une zone d'arrière-mangrove, soumise à une dessalure par les apports fluviatiles dominants, en raison des pourcentages importants de spores de fougères, de Bryophytes, d'herbacées, de pollens de Sparganium et de formes d'eau douce.
4 Discussion
4.1 Âge du « pic d'Avicennia »
Les différentes méthodes utilisées pour construire la stratigraphie de cette série deltaïque [8,9] nous permettent de contraindre l'intervalle stratigraphique des niveaux contenant le « pic d'Avicennia » (Fig. 4A) : l'étude palynologique suggère un âge Burdigalien supérieur–Langhien inférieur, en accord avec l'attribution biostratigraphique du gisement de micromammifères CR1 à la zone MN4. La chimiostratigraphie conduit à proposer un âge compris entre 15,8 et 16,5 Ma (âges de [3]), en accord avec les données biostratigraphiques (Fig. 4A et B).
À Châteauredon, le niveau noir contenant la plus grande proportion de grains d'Avicennia (RHCR1 et GUE01) est riche en dinokystes. Il surmonte des faciès marins francs (tidalites, 350–356 m, Fig. 4), et est recouvert par des dépôts régressifs (faciès de lagune, paléosols, 364–378 m, Fig. 4). Le pic d'Avicennia est donc contemporain de la fin d'un haut niveau marin. Nos données stratigraphiques permettent de l'attribuer au haut niveau marin du début du Langhien, cycle TB 2.3 [17] ou entre les limites de séquences Bur5/Lan1 et Lan2/Ser1 [18] (Fig. 4C).
La présence d'une mangrove à Avicennia a déjà été signalée en Méditerranée occidentale par Bessedik dans des niveaux dont l'âge va du Chattien au Langhien supérieur [4,6]. La mangrove a persisté au Messinien en Sicile [28] et en Algérie [12]. Cependant, le développement maximal de cette formation végétale, dont témoignent les forts pourcentages en pollen du genre Avicennia dans une série de coupes allant de la Catalogne à la vallée du Rhône, se situe au Burdigalien supérieur–Langhien [6].
Ce maximum ou « pic d'Avicennia », est très réduit dans le temps, comme l'ont montré Bessedik et Cabrera [7]. Ils ont pu préciser son âge à l'aide d'associations de foraminifères et de nannofossiles dans le bassin de Vallès–Penedès (Espagne) ; cet âge est compris dans un intervalle comprenant Helicosphaera ampliaperta (LAD à la limite NN4/NN5) et Globigerinoides sicanus (équivalent de Praeorbulinoides sicana, FAD à la limite N7/N8, Fig. 4B). Par ailleurs, Praeorbulina glomerosa (FAD intra-N8) est observée dans des marnes situées latéralement à celles contenant le pic d'Avicennia (Fig. 4 in [7]). Toutefois, la position stratigraphique relative de leurs échantillons reste incertaine, et ne permet de restreindre l'intervalle stratigraphique qu'à une partie des zones N8–NN4 (Fig. 4C). Ces informations, une fois replacées sur l'échelle stratigraphique de Berggren et al. [3], conduisent à proposer pour le pic d'Avicennia un âge compris entre 15,6 et 16,4 Ma (Fig. 4C). Notre datation du pic à partir d'autres méthodes stratigraphiques conforte la durée maximale de cette extension, que nous estimons à 700 ka.
Par ailleurs, les mêmes auteurs ont souligné le contemporanéité du pic d'Avicennia et de l'extension maximale des coraux du Miocène moyen, bien que, dans le bassin de Vallès–Penedès, les échantillons contenant Avicennia soient situés stratigraphiquement légèrement au-dessus des deux niveaux récifaux. L'extension des récifs coralliens est observée, au Miocène moyen, dans tout le Bassin méditerranéen, dans un intervalle compris entre N7 et N9. Elle est ainsi rapportée au haut niveau marin du Langhien [14,15] (synthèse in [16]). Dans cette logique, Buchbinder et al. [10] font correspondre l'extension récifale au cycle TB 2.3 dans la charte de Haq et al. [17] et à la base du cycle suivant (TB 2.4). La présence du pic d'Avicennia au-dessus de niveaux marins francs dans la série de Châteauredon corrobore aussi la concomitance de la mangrove avec le haut niveau marin langhien.
4.2 Implications paléoclimatiques
Les niveaux à Avicennia et les récifs coralliens associés décrits au Miocène moyen en Méditerranée occidentale caractérisent une période d'optimum climatique dont l'âge a été discuté ci-dessus. Cette période chaude est confirmée par la présence d'éléments des flores tropicales mégathermes peuplant aujourd'hui, pour la plupart, l'Asie du Sud-Est. Ils sont par ailleurs connus au Miocène moyen, en Espagne (Aragonien de Burgos et de Teruel) [2,31], au Portugal (bassin du Tage) [1], en Hongrie–Bulgarie (Bassin pannonien) [19,22] et en Allemagne (formation de Ville) [30].
Dans les échantillons de Châteauredon, les pourcentages de grains de pollen d'Avicennia varient de 4 à 12% du matériel pollinique compté (Fig. 3). Ces valeurs sont supérieures à celles observées sur les coupes du Languedoc, où elles sont généralement inférieures à 1% dans l'intervalle Burdigalien–Langhien, à l'exception de la coupe de Montpeyroux, proche de Montpellier, où elles atteignent 6 à 8%. En revanche, ces valeurs sont plus faibles qu'en Catalogne, où des pourcentages de 15 à 20% ont été obtenus [6].
Les températures et les valeurs hygrométriques avancées pour cette période varient selon la nature continentale ou marine des espèces fossiles prises en compte. Les températures moyennes annuelles évoquées par différentes études vont de 20 à 23 °C au Portugal [1] à 18 °C pour le Nord-Ouest de la Méditerranée [5], à 20 °C en Allemagne [30] et dans le Bassin pannonien [22]. Les températures des mois les plus chauds auraient atteint une moyenne de 28 °C et celles des mois les plus froids seraient restées au-dessus de l'isotherme 10 °C. Ces températures sont compatibles avec celles suggérées par l'étude des coraux [11] pour les eaux marines. Elles seraient de 18 à 20 °C pour la saison la plus froide. Les précipitations annuelles oscillaient localement entre 1300 [30] et 1400 mm [22].
La répartition des mangroves actuelles appauvries, où ne subsistent que les espèces d'Avicennia, est conforme à ces valeurs, comme le suggèrent les environnements de mangrove d'Australie du Sud-Est se développant dans un climat subtropical [26].
La présence de la mangrove aux alentours de Digne positionne cet environnement plus au nord que ce qui était connu en France, puisque les grains d'Avicennia les plus septentrionaux trouvés en Provence appartiennent aux coupes proches de l'Étang de Berre (Pont-Gayé, Tamaris [6], Bayanne [21] et Besson, communication orale).
Les reconstitutions paléogéographiques placent la région de Digne à la paléolatitude d'environ 42°N [13]. Le développement d'une mangrove, même appauvrie, à une telle latitude a été également observé au Japon à la même époque (40–44°N, biozone N8) [20,32], et en Hongrie (42–44°N, Langhien) [25,27]. Ces données paléolatitudinales indiquent un décalage de 20° vers le nord de l'extension des mangroves par rapport à l'Actuel [26].
L'extension à des latitudes plus septentrionales est à mettre en relation avec la dynamique climatique globale du Miocène moyen. Si la présence d'une calotte antarctique est avérée au Miocène moyen, celle du pôle Nord n'est pas admise avant 7 Ma [33]. L'absence d'inlandsis au pôle Nord permettait un gradient climatique plus doux entre les pôles et l'équateur, autorisant aux mangroves une plus large bande latitudinale de développement [26]. Par conséquent, l'extension de la mangrove à Avicennia à des latitudes plus septentrionales est à mettre en relation bien plus avec ce faible gradient, qu'avec une augmentation significative des températures [26].
Cette mangrove pourrait ainsi appartenir à l'extension la plus septentrionale de ce peuplement végétal connu en basse Provence à la même époque, mais qui existait probablement aussi en Hongrie [25,27].
5 Conclusion
La présence de grains de pollen du genre Avicennia est documentée dans la série de Châteauredon (Alpes-de-Haute-Provence), bien contrainte stratigraphiquement. Elle constitue un marqueur stratigraphique et paléoenvironnemental intéressant pour le Miocène moyen :
- – l'attribution stratigraphique des échantillons ayant livré Avicennia est obtenue à partir d'une stratigraphie intégrée associant bio- et chimio-stratigraphies. L'âge obtenu sur l'échelle de Berggren et al. [3] est compris entre 15,8 et 16,5 Ma. Ce résultat est parfaitement en accord avec les données biostratigraphiques de gisements de paléo-mangroves à Avicennia du domaine méditerranéen (N7/N9) et plus particulièrement du bassin de Vallès–Penedès (N8–NN4 pro parte). La datation vient préciser la courte extension temporelle du pic, estimée à 700 ka maximum ;
- – ces grains de pollen sont associés, à Châteauredon, à des faciès transgressifs en contexte deltaïque qui sont rapportés, d'après l'attribution stratigraphique, au haut niveau marin du Langhien (cycle TB 2.3). Cette observation place la mangrove de Châteauredon dans un contexte comparable à celui de la mangrove de Vallès–Penedès, associée, elle, à des récifs dont l'extension maximale est reconnue en Méditerranée au cours du même cycle eustatique ;
- – la découverte d'une mangrove appauvrie (l'Avicennia étant le seul palétuvier identifié) et l'interprétation des données palynologiques des mêmes horizons dans la région de Digne impliquent (i) un contexte climatique de type subtropical plus ou moins humide à une paléolatitude de 42°N et (ii) une extension septentrionale de la mangrove connue plus au sud en Provence.
Remerciements
Les auteurs tiennent à remercier Anne-Marie Lézine (laboratoire LSCE du CEA) pour avoir fourni les lames d'Avicennia alba, marina et africana provenant de mangroves actuelles. Les remarques et conseils d'A.-M. Lézine, J.-C. Plaziat (laboratoire IDES de l'Université d'Orsay) et M. Durand-Delga ont nettement contribué à l'amélioration du manuscrit initial.