

1 Introduction
As the Eurasian beaver Castor fiber L. 1758 has declined over large regions in Europe, wild caught animals have been reintroduced on several occasions to restore lost populations [1–4]. In particular, after the disappearance of the Eurasian beaver Castor fiber galliae Geoffroy 1803 from the Loire valley (France) at the beginning of the 19th century, individuals from the Rhône River were released in the Loire valley near Blois, between 1974 and 1976 [5,6]. The released animals propagated on the Loire up- and downstream from Blois, and on most tributaries [7]. However, the colonization dynamic of the Loire River appears slower than that observed in other areas, such as Sweden [1]. The importance of intact riparian woods for conservation of healthy populations has also been underlined [2]. On the Loire River, downstream from Blois, a length of Salicaceae woods dominated by Salix alba L. and Populus nigra L. appears correlated to home range size [7]. Willow woods were often mentioned as the habitat where beavers attain a higher fecundity [2] but there is no evidence that this kind of relationship is due only to trophic needs. For their propagation, beavers must also find proper sites for lodge establishment.
Beavers are known to select lodge places based on water depth and bank characteristics [1,8] but, as yet, the influence of habitat characteristics on lodge establishment remains poorly documented. For C. fiber, as for the American beaver Castor canadensis Kuhl, habitat characteristics have been mainly investigated in relation to dam establishment [9], tree cutting, or beaver density [1,7,10–13]. Salicaceae are often mentioned as the bulk of beaver diet [2]; nevertheless beavers also cut branches of Salicaceae in order to build huts and dams, and to cover hut-borrows [8,9,14]. Salicaceae abundance may therefore influence lodge location, even though beavers are able to use numerous other woody plants for construction [15–17]. Barnes and Mallik [18] stressed the importance of plant structure, particularly stem diameters, for site selection when dams are built. Similarly, the size of trees or bushes on the shoreline may influence lodge location. On the other hand, Richard's description of burrows indicated that beavers usually dig galleries under a tree [8], and thus use vegetation even when lodges are not covered with branches.
We therefore assume that Salicaceae, combined with plant structure in the woods along the shoreline, influence selection of sites for lodge building. In order to find predictors for possible locations of new lodges, we searched for relationships between lodge occurrence and vegetation structure, Salicaceae canopy cover, and bank characteristics in an area already colonized by beavers. Then, we compared the site characteristics between this area and a downstream strech where beavers are not yet established. Finally, we estimated the number of proper sites for future lodges.
2 Study area and methods
2.1 Study area
The Loire valley, in France, is enclosed by the old Hercynian folds of the Armorican peneplain in the northwest, the Massif Central in the south and the Morvan in the east. From the secondary era, successive sea deposits of sand and clay interspersed with limestone and loess, leading to the formation of a vast alluvial plain. The Loire carries sand and gravel that form changing sandbanks and islands. Water level varies greatly; receiving little rainwater supply in summer, the river reaches its lowest level in September (160–), while shoreline woods are flooded in the winter (1320– February in Saumur-Montjean; [19]).
Riparian woods are characterised by three main plant communities. The small Willow Grove (less that 5 to 6 m high) includes pioneer species (Osier Salix purpurea L., Salix triandra L. and Salix viminalis L.). The tall Willow Grove (15–20 m high) is dominated by white Willow S. alba L., crack Willow Salix fragilis L. and black Poplar P. nigra L. often combined with the american box Elder Acer negundo L. [19,20]. On the top of the riverbank, narrow-leaved Ash (Fraxinus angustifolia Vahl) and smooth-leaved Elm (Ulmus minor Mill.) dominate accompanied by common Oak (Quercus robur L.). Banks and islands are sometimes protected from erosion by ripraps.
2.2 Field study
The studied part of the Loire River began 127 km downstream from the city of Blois. It was divided in two areas, one located east of Angers city (U-area) and the other located west of Angers (D-area). The U-area began at Montsoreau and ended at Bouchemaine, a 80-km long strech, where the first lodge was found in 1985. In 1996, five sites with several active lodges were recorded on this area [16]. D-area, located between Chalonnes-sur-Loire and Ancenis (36 km), was a part of the river where beavers have not established yet.
The field study was conducted by canoe and by foot from July to September 2000 and 2001. Low water level allowed an easy access to riparian woods, and a better detection of beaver signs and of sandbanks. Both sides of the Loire were surveyed, as were the numerous islands. In U-area, 90 kilometers of contiguous shoreline were surveyed in 2000, with approximately 20 km of diked bank. In D-area, 88 km of shoreline were surveyed in 2001. In streches, the whole bankline was divided into 10-m wide contiguous plots (or sites), parallel to the stream. Plots varied from 0.6 to 1.5 km in length and corresponded to zones where vegetation (structure and composition) appeared similar. On each plot, plants were identified and categorized based on their morphology: (TT) 10–15-m tall trees with large trunks and no branch touching the water at flood time, (ST) 5–10-m small trees, (B) 5–12 m tall bushes with basal branches accessible to beavers even at low water level, (H) herbs and forbs. Within each class of woody plants, Salicaceae were respectively called TTS, STS and BS. At each plot, canopy cover by each plant category was estimated using an Abundance-Dominance scale (AD), ranging from 1 to 5: (1)<5%, (2) 5–25%, (3) 25–50%, (4) 50–75%, and (5) 75–100% [21]. The number of lodges was recorded, as the signs of beaver activity (cut trees, refectories, bark stripping, trunk gnawing, tracks...). The bank slope was classified as: (1) <20%, (2) 20–50% or (3) >50%. The percentage of bank length protected from erosion by ripraps, the characteristics of these ripraps and the presence of a contiguous sandbank in front of the site were also recorded. Human frequentation (mainly fishers and strollers) was assessed using a qualitative scale ranging from 1 to 5.
2.3 Data analysis
Data collected in U-area were analysed to compare plots with lodges, plots with other beaver signs and plots without beavers. The 20-km diked bank was excluded. As the number of plots within each group was small, we performed a Kruskall–Wallis nonparametric ANOVA. Multiple comparison Dunn's post test was used for means separation. We then conducted multivariate analysis (Automatic Interaction Detection, AID) with lodge occurrence as dependant variable, in order to identify factors influencing lodge location (minimum split value 0.05, Phi-square fitting method, Instat software). A first AID was conducted with all U-plots. A second was conducted after removal of the plots without beavers. Student's t-tests were performed to compare habitat variables between up- and downstream sites. Welch t-test was used when variances were unequal. U- and D-plots were examined regarding their suitability for lodge construction according to the AID results. Plots that fitted the model within both datasets were compared using a Mann–Whitney nonparametric test.
3 Results
3.1 Beaver activity in upstream area
The surveyed U-area shoreline was divided into 86 plots. One to four lodges were recorded at 19 plots and other beaver signs were observed at 36 plots. All these signs of activity were mainly recorded on island edges: 70% of the lodges, 65% of cut trees, refectories, bark stripping or tracks.
Analysis of vegetation structure (Table 1) indicated that the canopy cover by tall trees was higher in the plots with lodges (35.1%, SD 19.3%, KW 9.62, P=0.008, df=2). Plots with lodges have significantly higher tall Salicaceae canopy cover than other plots (11.8%, SD 10.3%, KW 9.5, P=0.009, df=2). Beavers settled down in rather quiet places (mean human frequentation 1.2 in sites with lodges, SD 0.4, KW 12.5, P=0.002, df=2). Even though ripraps were most found in sites without beavers, the animals did not always avoid bank protection to build lodges. Most of ripraps were made of non-cemented stones, which length ranged from 20 to 40 cm and width from 15 to 30 cm (n=23). Stones never covered more than 40% of the plot bank, and lodges were observed in the stoneless areas. At two sites, beavers managed to dig between adjacent stones: in both cases, 100% of the bank was covered with bigger stones (50–100 cm long, 30–35 cm wide, n=23) that were not cemented to each other and interstices were large enough (mean 27.3 cm, SD 9.7, n=13) to allow an access to the bank substrate. No beaver signs were observed in two other sites, continuously covered with a flagstone pavement.
Comparison of habitat variables among sites with lodges, sites with other beaver signs, and sites without beavers, in the U-area. Mean values (SD), Kruskall–Wallis test
| No beavers | Other beaver signs | Lodge | P a | |
| n=32 | n=36 | n=19 | ||
| Total plant cover (%) | 86.1 (13.4) | 80.5 (13.4) | 85.3 (7.9) | 0.134 |
| Tall trees (%) | ||||
| All species (TT) | 20.0 (16.6) | 19.2 (19.7) | 35.1 (19.3)∗∗ | 0.008 |
| Salicaceae (TTS) | 5.5 (7.8) | 5.5 (7.2) | 11.8 (10.3)∗ | 0.009 |
| Small trees (%) | ||||
| All species (ST) | 9.7 (12.1) | 12.6 (16.5) | 3.7 (5.5) | 0.161 |
| Salicaceae (STS) | 1.6 (3.1) | 2.8 (6.8) | 0.8 (1.2) | 0.592 |
| Bushy plants (%) | ||||
| All species (B) | 13.9 (19.9) | 17.1 (18.4) | 10.9 (8.1) | 0.174 |
| Salicaceae (BS) | 5.8 (9.6) | 6.0 (7.4) | 4.9 (4.7) | 0.269 |
| Herbs and forbs (%) | 2.7 (1.1) | 2.5 (0.8) | 2.2 (1.1) | 0.125 |
| Bank slope | 2.5 (0.6) | 2.8 (0.5)∗ | 3.0 (0.0)∗∗ | 0.002 |
| Human frequentation | 2.0 (0.8) | 1.6 (0.7) | 1.2 (0.4)∗∗∗ | 0.027 |
| % plots with sandbank | 30.5 | 28.0 | 0 | – |
| % plots with ripraps | 34.3 | 19.4 | 10.5 | – |
a Assuming Chi-square distribution with 2 df.
∗ P<0.05,
∗∗ P<0.01,
∗∗∗ P<0.001: Dunn's multiple comparison test.
Among the 34 lodges, 12 were burrows, 20 hut-burrows, and two were temporary huts built on the river edge. Beavers always dug their lodges into abrupt banks (slope>50%), with the absence of sandbank (Table 1). Rooms and galleries were always burrowed between roots of a 10–15-m tall tree. A lot of these trees were poplars (about 44%, n=32); only one willow was used (S. alba, 3%). However, most trees were non-Salicaceae (53%): F. angustifolia (31%), Q. robur (13%), U. minor (6%), and A. negundo (3%).
3.2 Factors influencing lodge location
The AID carried out with all U-plots (R2=0.468; Fig. 1a) clearly related plots with lodges with a canopy cover by tall trees higher than 37.5% (AD⩾3), sandbank absence, and canopy cover by tall Salicaceae higher than 12.5% (AD⩾2). The second AID with only sites with beaver signs (R2=0.484) added as factor bushy Salicaceae higher than 2.5% (AD⩾1; Fig. 1b). Among the 19 plots with lodges, only six of them did not fit this model. Regarding the plots without beavers, only one fitted the model, but at that site, the bank was protected by a continuous small-stoned riprap.
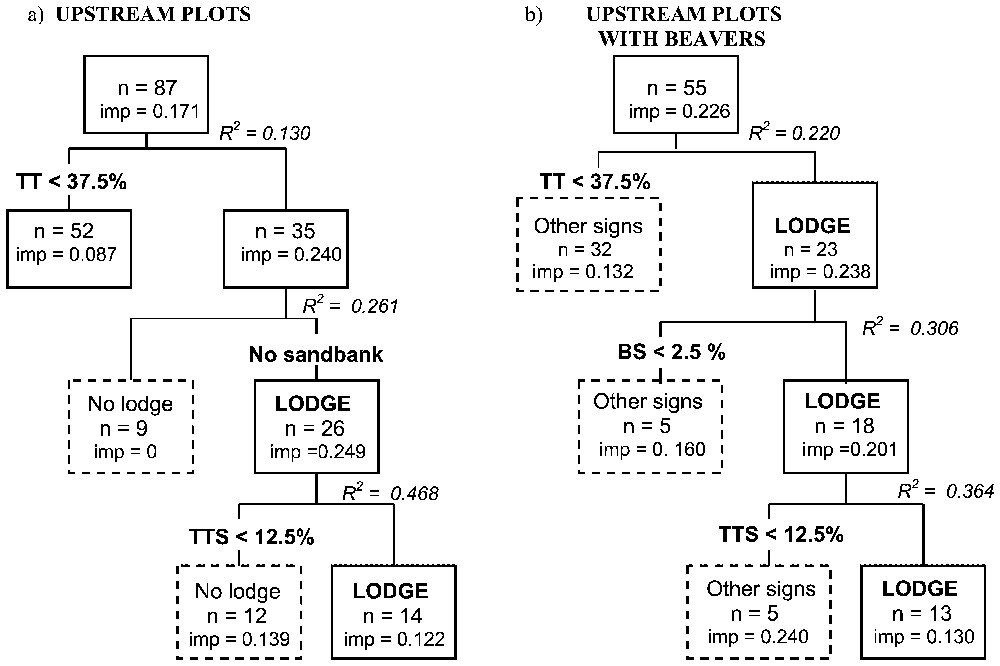
AID revealing factors influencing lodge occurrence. (a) Mobile obtained with all U-plots, (b) Mobile obtained after removal of U-plots without beaver sign. TT=tall trees, TTS=tall Salicaceae trees, BS=bushy Salicaceae, n=number of plots, imp=impurity.
3.3 Habitat characteristics of the downstream area
In D-area, the surveyed part of shoreline was divided into 89 plots. The canopy cover by tall trees and bushy plants did not differ between up- and downstream riparian woods (Table 2). Regarding tall Salicaceae, values were higher in D-area compared to U-area (mean 13.5%, SD 16.2%, P=0.0009, t=3.42, df=128). With a mean cover percentage of 35.8% (SD 25.1%), D-plots were also significantly richer in small trees than U-plots (mean 9.7%, SD 13.7%, P<0.0001, t=8.54, df=132). This difference was also verified for small Salicaceae trees, whose mean cover reached 9.0% in D-area (SD 12.3%), and 2.1% in U-area (SD 4.9%, P<0.0001, t=4.86, df=112). Both areas did not differ in the abundance of bushy plants. Moreover, from U-plots to D-plots, comparison of canopy cover by woody species, showed a significant decrease of S. fragilis (P=0.0031, t=3.03, df=105) and an increase of S. alba (P<0.0001, t=4.028, df=136). P. nigra cover markedly rose up, reaching 9.42% in U-plots (SD 13.2%), against 23.3% in D-plots (SD 18.7%, P<0.0001, t=6.71, df=158). Regarding non-Salicaceae species, F. angustifolia was more dominant in D-area (P<0.0001, t=6.44, df=139), likewise U. minor (P=0.0027, t=3.045, df=168). No significant differences in bank slope, the presence of sandbank and human frequentation was found between both areas. Ripraps were however more abundant in D-plots (mean 44.8%, SD 32.7%, P<0.0001, t=5.9, df=173).
Comparison between U- and D-areas in canopy cover (%). Mean values (SD)
| U-plots | D-plots | P | |
| n=86 | n=89 | ||
| Total plant cover | 83.4 (12.5) | 90.1 (8.9) | ns |
| Main woody species | |||
| Salicaceae | |||
| Populus nigra | 9.4 (13.2) | 23.3 (18.7) | <0.0001 |
| Salix alba | 2.9 (7.1) | 9.0 (12.1) | <0.0001 |
| Salix fragilis | 2.4 (5.1) | 0.7 (1.7) | 0.0031 |
| Other species | |||
| Fraxinus angustifolia | 8.3 (10.9) | 23.1 (18.4) | <0.0001 |
| Ulmus minor | 3.0 (6.4) | 6.2 (7.2) | 0.0027 |
| Plant structure | |||
| Tall trees | |||
| All species (TT) | 22.4 (19.5) | 20.5 (21.0) | ns |
| Salicaceae (TTS) | 6.8 (8.4) | 13.5 (16.2) | 0.0009 |
| Small trees | |||
| All species (ST) | 9.7 (13.7) | 35.8 (25.1) | <0.0001 |
| Salicaceae (STS) | 2.1 (4.9) | 9.0 (12.3) | <0.0001 |
| Bushy plants | |||
| All species (B) | 14.9 (17.3) | 15.6 (15.7) | ns |
| Salicaceae (BS) | 5.7 (7.7) | 7.3 (7.8) | ns |
| Herbs and forbs | 27.7 (23.1) | 34.5 (23.6) | ns |
Twenty-eight D-plots fitted the model. Sixty four percent of them were located on islands. Compared with U-sites with lodges that fitted the model, these plots were markedly richer in tall Salicaceae (mean 31.2%, SD 15.2%, P=0.004, U=222; Table 3), in small trees (mean=25.4%, SD 21.9%, P<0.0001, U=258.5), small Salicaceae (mean=6.1%, SD 10.4%, P=0.001, U=232.5), and in herbs (mean=27.6%, SD 20.8%, P<0.0001, U=252). With a mean of 1.92 (SD 0.76), human disturbance at these plots was higher than at U-plots with lodges (P=0.016, U=210).
Comparison between plots located in U- and D-area and fitting the model. Mean values (SD)
| U-plots | D-plots | P | |
| n=12 | n=28 | ||
| Total plant cover (%) | 86.7 (8.1) | 91.7 (7.6) | ns |
| Plant structure | |||
| Tall trees (%) | |||
| All species (TT) | 41.7 (9.7) | 46.9 (12.3) | ns |
| Salicaceae (TTS) | 15.8 (10.5) | 31.2 (15.2) | 0.004 |
| Small trees (%) | |||
| All species (ST) | 1.7 (3.6) | 25.4 (21.9) | <0.0001 |
| Salicaceae (STS) | 0.6 (1.1) | 6.1 (10.4) | 0.001 |
| Bushy plants (%) | |||
| All species (B) | 11.9 (9.4) | 14.6 (16.0) | ns |
| Salicaceae (BS) | 5.6 (5.1) | 8.0 (8.1) | ns |
| Herbs and forbs (%) | 2.3 (1.0) | 27.6 (20.8) | <0.0001 |
| Bank slope | 3.0 (0.0) | 2.9 (0.3) | ns |
| Human frequentation | 1.3 (0.5) | 1.9 (0.7) | 0.016 |
| % of plots with ripraps | 16.7 | 78.6 | – |
4 Discussion
According to our results, in sites without a sandbank in the front, tall trees are good predictors for lodge location when they cover more than 37.5% of the riverbank, provided that more than a third are Salicaceae. On the Loire River, the diameter of branches cut by beavers averages 5.5 cm [1], and Barnes and Mallik showed that C. canadensis selects diameters lower than 5 cm to build dams [18]. Surprisingly, while bushes and young trees provide these trunk and stem sizes, the cover by bushy plants retained by the AID is weak (2.5%) and the 5–10 m trees do not influence lodge site selection. Actually, when tall trees around the lodge are not Salicaceae, beavers can use branches of small willows and poplars located in a neighbouring plot.
On the Loire River, as on the Rhône [8], abrupt banks are favourable to dig rooms and galleries to form burrows and hut-borrows, rather than huts. These unobtrusive kinds of lodges are more resistant to flow changes. Beavers continuously adapt and modify the structure in order to keep an entry under water in all seasons. Therefore, they avoid sites with a contiguous sandbank along the edge, not to be constrained to desert their lodge at low water level. However, sandbanks slowly move with the stream and lodges may progressively be sanded. In the aranaceous substrate of the Loire banks, beavers always burrow under a 10–15-m tall tree, using the strong root system as propping frame to prevent galleries from collapse. For this purpose, they often choose poplars, most common tall Salicaceae in this area. However, they are able to settle under different non-Salicaceae trees, particularly ashes that represent the most dominant species, and they also use roots of non Salicaceae trees.
Among the seven U-sites with lodges that did not fit the model, two of them (Ardouin and Parnay Islands), were occupied by beavers before 1995 [16]. From 1996 to 2000, the vegetation was uprooted from these islands leading to a severe degradation of the habitat, but beavers stayed in the vicinity of previous lodges. By contrast, the five other plots did not present any lodge before 1996 [16]. Abundance of 10–15 m tall trees at these sites is low (cover ⩽12.5%). Nolet and Rosell [2], who studied translocated beavers in the Netherlands, observed that they first established in the richest habitats before occupying the others. This result may explain why plots with recent lodges do not fit the AID.
Canopy cover by P. nigra and S. alba is clearly higher in D-area compared to the U-area. As home-range size decreases with the abundance of black poplars and white willows [1], the D-area should logically accommodate a higher density of lodges than U-area. Both structure and composition of downstream vegetation seem globally more favourable, regarding lodge building. With 28 suitable sites, results suggest an easy colonization across the D-area (36 km long). However, these sites may be available, provided that beavers adapt to ripraps that occur in 75% of them. Most of these embankments are discontinuous and made of small stones similar to those previously described upstream, where they limit possible lodge establishment in suitable sites. The others present uninterrupted flagstone pavements, and we do not know if beavers will choose to use that kind of banks. Downstream, beaver will also have to face a more intense human activity, as the river has been threatened for boats. Banks have been protected with ripraps, and groynes perpendicular to the stream have been built on both river sides to retain sand and to keep a channel in the middle of the river. All these alterations have led to a degradation of riparian woods. However, beavers are very plastic animals that can survive and reproduce in different landscapes [2]. On the assumption that beavers adapt to the altered habitat of the downstream part of the Loire River, we may expect that factors influencing lodge building will not be the same.
Preservation of willow woods dominated by S. alba and P. nigra is necessary to allow a further colonization of the downstream Loire River [7]. However, it seems important to preserve quiet places with 10–15 m tall trees to allow beaver settlement. As they may shelter lodges, tall trees are also to preserve in places where Salicaceae are sparse, beavers being able to move to feed or to cut branches from plants of neighbouring places. Bank protection with ripraps is not necessary incompatible with beaver settlement, provided they are built with stable stones with large interstices between them.