

Abridged English version
Over the past 10 years, there has been a notable resurgence of interest in microphytobenthos, an important component of marine sedimentary ecosystems. The unicellular algae and cyanobacteria constituting this group proliferate in the upper centimetres of intertidal sediments, providing an estimated 50% or more of the primary production for such ecosystems. As access to these areas is often difficult, most studies of the structure and dynamics of microphytobenthos assemblages have been based on a limited number of samples, which means that the spatial and temporal structures of these assemblages are not easy to resolve on a meso- or macro-scale.
This study applied SPOT satellite multi-spectral remote sensing (visible–infrared) techniques to assess microphytobentos assemblages of a shellfish ecosystem in the Bourgneuf Bay, south of the Loire Estuary in France. Microphytobenthic organisms form biofilms on the sediment surface that are coloured by photosynthetic and accessory pigments in the cells. These pigments are characterized by reflectance spectra and can be detected by visible-infrared sensors. Data were measured using a GER 3700 spectroradiometer for the main constituents of the study area (sand, mud, microphytobentos, macroalgae, etc.) in order to calibrate SPOT satellite images. Spectral signatures of microphytobenthos biofilm (composed of 99% diatom cells) were analysed and compared with the spectral signature of a phaeophyceae (Fucus spp.) common on rocky substrates of the bay. Both groups showed common absorption bands for visible wavelengths due to cellular pigments: chlorophyll a at 673 nm and a mixing of chlorophylls a and c at 632 nm. As macroalgae produced a strong reflection in the infrared band, as compared to microphytobenthos, this feature was used to discriminate both components on SPOT images after calculation of the normalised vegetation index (NDVI). These results were confirmed by field observations.
Comparison of SPOT images (acquired at low tide in spring or autumn between 1986 and 1998) required calibration with the same light conditions, which was performed on the basis of an ‘empiric line correction’, using bright (white) and dark (black) references on the field and the image (respectively dry sand and non-turbid water). After calibration, the NDVI provided accurate information on vegetation coverage. The microphytobenthos of the mudflat area exhibited NDVIs of 0 to 0.3, whereas those of rocky areas covered with macroalgae were greater than 0.3. Microphytobenthos proliferated mainly along a north-south strip, essentially localised around the +2-m isobath (Fig. 5). These results suggest that bathymetry is one of the factors controlling the spatial distribution of microphytobenthos in the Bourgneuf Bay mudflat. A second factor is the great abundance of microphytobenthos close to oyster-farming areas, which suggests a relationship between cultivated oysters and microphytobenthos. This supports the hypothesis that a retroactive process allows benthic microalgae to use the dissolved organic matter excreted by bivalves as well as the nutrients released after mineralisation of their faeces and pseudo-faeces. A third factor, the Falleron River and its channel, seems to control the spatial structuring of microphytobenthos assemblages. This large outlet from nearby marshes, with a mean annual flow of , is responsible for high nutrient input.

Répartition spatiale du microphytobenthos : résultats du traitement des six images SPOT (Tableau 1) après étalonnage, en terme d'indice de végétation normalisé (NDVI). L'échelle de couleurs associée au NDVI (0>NDVI>0,3) renseigne sur l'importance de la couverture végétale, y compris celle du microphytobenthos (Tableau 3). Sont exclus de l'échelle les pixels dont le NDVI est inférieur à −0,1 et supérieur à 0,3. Les isobathes −2 m, 0 et +2 m des cartes marines sont reportées, les parcs ostréicoles de la Coupelasse sont indiqués par la lettre C encadrée. Les flèches noires indiquent les zones de seillons et banquettes où peuvent se développer des biofilms microphytobenthiques. L'échelle représente 2 km.
Finally, the specificity of the micophytobentos study was lower because of the limited spectral and spatial resolution of images. SPOT images do not have sufficient spectral resolution to distinguish the absorption band at 673 nm (chlorophyll a) from that at 632 nm (chlorophylls a and c). Microphytobenthos could only be mapped in this case by the NDVI based on broad wavelength bands and with the support of field knowledge. Thus, the use of hyperspectral sensors appears essential to study the pigment assemblage of microphytobenthos and map the biomass more correctly.
1 Introduction
Les écosystèmes côtiers comportant de grandes surfaces de sédiments intertidaux font l'objet depuis ces dix dernières années d'un effort de recherche concernant un maillon jusqu'alors négligé, bien qu'essentiel pour la compréhension de leur fonctionnement, le microphytobenthos. Ce terme désigne l'ensemble des algues unicellulaires eucaryotes et des cyanobactéries se développant dans les premiers millimètres du sédiment. Ces producteurs primaires benthiques font de ces habitats un des plus productifs de la biosphère [1]. Des travaux récents ont mis en évidence le rôle majeur de cette production primaire benthique par rapport à celle d'origine phytoplanctonique [2]. Elle peut représenter jusqu'à 50 % de la production primaire totale dans certaines régions estuariennes [3] et être dominante sur les vasières intertidales [4,5]. Remis en suspension par les courants de marée ou le clapot [6], le microphytobenthos occupe ainsi une part importante, souvent sous-estimée, dans les flux de matière au sein des écosystèmes côtiers [7,8].
Jusqu'à présent, la structure et la dynamique du microphytobenthos des vasières littorales ont fait l'objet d'études basées sur des échantillonnages ponctuels [4,5,9,10]. En effet, ces écosystèmes s'avèrent souvent difficiles d'accès en raison de la faible cohésion des sédiments fins. Cette pénibilité limite le nombre de mesures réalisées, pourtant indispensables pour appréhender la structure spatiale et temporelle des peuplements.
L'objectif de ce travail est d'évaluer l'intérêt de la télédétection multispectrale dans les domaines des rayonnements électromagnétiques visible et infrarouge, pour l'étude des peuplements du microphytobenthos à l'échelle d'un écosystème. Ces microorganismes, migrant à la surface du sédiment lors des marées basses diurnes [8,11,12] y forment des biofilms colorés par la présence de pigments cellulaires photosynthétiques et accessoires. Ces biofilms peuvent être caractérisés par leurs spectres de réflectance et détectés par des capteurs satellitaires ou aéroportés [13,14]. Certains de ces pigments sont présents chez les macrophytes aquatiques et sont déjà utilisés pour leur cartographie par télédétection [15–18].
L'étude ici décrite des peuplements du microphytobenthos repose sur le traitement d'images satellitales SPOT étalonnées à partir de mesures radiométriques de terrain. Une cartographie de la répartition spatiale de ces peuplements à des dates différentes entre 1986 et 1998 a été obtenue pour la baie de Bourgneuf, France, et ont permis de poser des hypothèses quant aux facteurs intervenant dans la structuration spatiale de ces peuplements : la bathymétrie, la présence de parcs ostréicoles et l'apport d'éléments nutritif par un des plus importants exutoires des marais avoisinants, le Falleron. Dans cet écosystème conchylicole affecté par de mauvaises croissances des huı̂tres en élevage [19], l'importance du transfert trophique du microphytobenthos vers les bivalves cultivés n'a jamais été quantifiée. La contribution potentielle de la télédétection visible–infrarouge à la problématique générale de la capacité trophique des écosystèmes conchylicoles est évoquée sur la base des contraintes liées aux résolutions spatiales et spectrales. De plus, le processus de migration verticale des microalgues a été pris en compte lors de l'interprétation de la répartition spatiale des peuplements. En effet, ce comportement semble être a priori une limitation à la cartographie du microphytobenthos par télédétection, en raison d'une possible sous-estimation de l'importance des ces peuplements. Enfin, la portée de ces réflexions dépasse largement le site d'étude et concerne la majorité des baies à vocation aquacole de la façade atlantique et de la Manche.
2 Présentation de la zone d'étude
La baie de Bourgneuf, située au sud de l'estuaire de la Loire (1°58′–2°15′W ; 46°53′–47°06′N) (Fig. 1) et soumise à l'action de la marée (marnage de l'ordre de 6 m lors des marées de vives-eaux), est le site d'une aquaculture extensive de l'huı̂tre Crassostrea gigas (Thunberg) qui la place au troisième rang des bassins d'élevage français, avec une production annuelle de l'ordre de 13 000 t. Elle comporte environ 10 000 ha d'espace intertidal, dont 1/10 est occupé par des parcs ostréicoles et des bouchots à moules. La turbidité y est élevée avec des concentrations journalières moyennes de matière particulaire en suspension fluctuant de 4 à [20]. La zone de la vasière étudiée occupe une surface de 2200 hectares (Fig. 1).
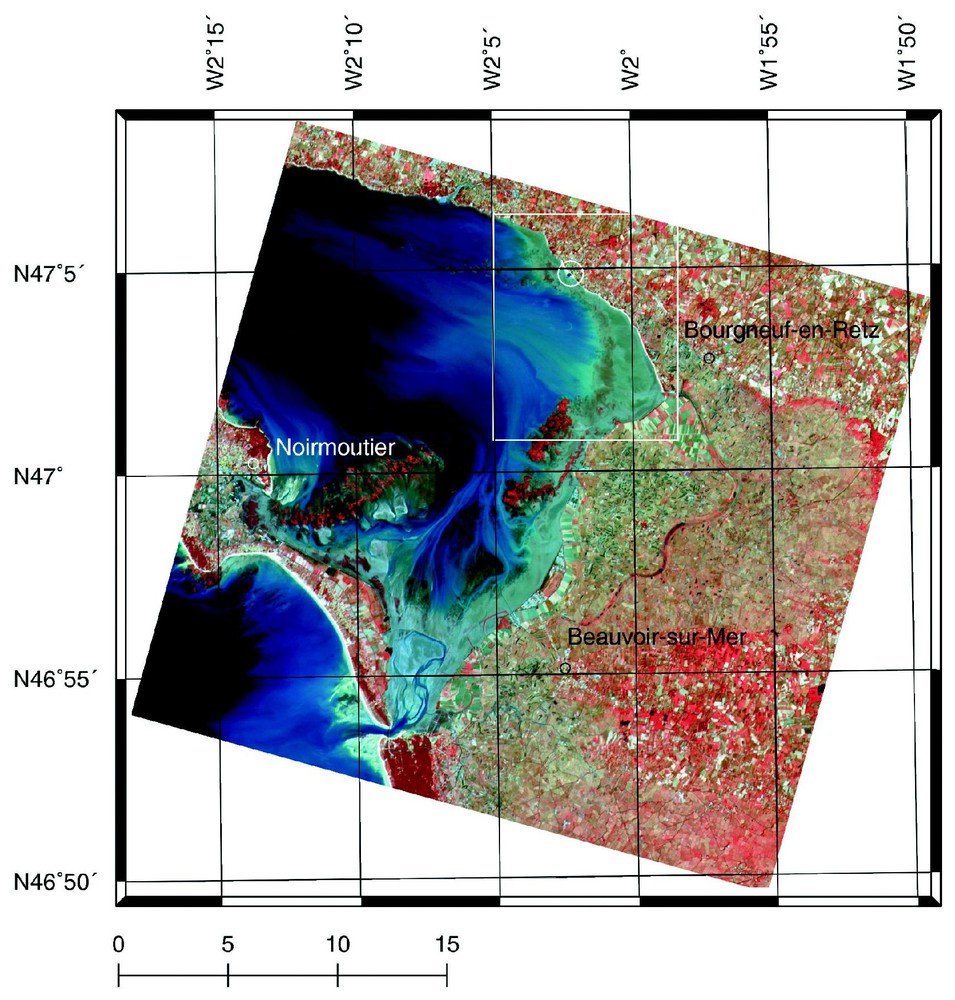
Baie de Bourgneuf (France) en vue satellitale SPOT. Extrait de la scène du 16 septembre 1998. La zone d'étude est encadrée, le site des campagnes de mesures radiométriques et la référence noire sont encerclés sur la commune de la Bernerie-en-Retz, à proximité des parcs ostréicoles. L'échelle est en kilomètres.
3 Spectroradiométrie de terrain
3.1 Méthodologie
La spectroradiométrie in situ établit le lien entre les principaux constituants du terrain et leur réponse spectrale, dans le but d'étalonner des images satellitales SPOT. La radiance R(i,λ) ou luminance de ces principaux constituants a ainsi été mesurée in situ avec un spectroradiomètre de terrain (GER 3700), en entre 300 et 2500 nm. La Fig. 2A montre l'exemple d'un biofilm de microphytobenthos se développant sur la vasière de la baie de Bourgneuf. L'énergie électromagnétique solaire incidente étant estimée à partir de la radiance d'un diffuseur parfait (Spectralon®, Fig. 2A), le calcul du rapport donne la réflectance ρ(i,λ) des principaux constituants du terrain (Fig. 2B). On s'affranchit ainsi des absorptions de l'atmosphère pour présenter une réflectance qui présente l'avantage de ne plus contenir qu'une signature caractéristique de la surface analysée, quelles que soient les conditions atmosphériques. Les spectres mesurés in situ sont représentés dans cette étude pour une gamme de longueurs d'onde couvrant les domaines visible et proche infrarouge (entre 400 et 1000 nm), pour lesquels les pigments cellulaires photosynthétiques et accessoires des organismes photosynthétiques présentent des bandes d'absorption spécifiques [21]. Au-delà de 1000 nm, l'absorption de la lumière infrarouge par la pellicule d'eau qui demeure à la surface de la vasière ne permet plus l'interprétation du signal.

Spectres mesurés sur la vasière de la baie de Bourgneuf, en septembre 1999, entre 400 et 1000 nm. (A) Spectres des radiances. En noir, l'énergie électromagnétique solaire réfléchie par le Spectralon®; en gris, la radiance d'un biofilm de microphytobenthos. Les flèches noires indiquent les bandes d'absorption dues à la vapeur d'eau de l'atmosphère, les grises, celles dues à l'oxygène. (B) Spectres des réflectances, calculées à partir des radiances. En noir, exemple d'une macroalgue brune (Phaeophycées) Fucus spp.; en gris, biofilm de microphytobenthos constitué à 99% de cellules de Diatomées; en ligne plus fine et noire, exemple de vase sans biofilm. Les flèches indiquent les bandes d'absorption spécifiques de la chlorophylle a (Chl a) à 673 nm et du mélange chlorophylle a et c (Chl c) à 632 nm.
Une première campagne de mesures radiométriques a été réalisée en septembre 1999, à proximité des parcs ostréicoles de la Bernerie-en-Retz (2°1,20′W ; 47°4,02′N) (Fig. 1). Les radiances des principaux constituants de cet écosystème sont alors mesurées : sable, vase, biofilm de microphytobenthos et macroalgues (Fucus spp. de la classe des Phaeophycées). Les stations de microphytobenthos sont choisies en fonction de l'intensité des colorations très variables du biofilm depuis les plus discrètes changeant à peine la couleur de la vase, jusqu'au brun sombre masquant totalement la vase sous-jacente. Les mesures radiométriques sont faites par le détecteur placé à 1 m au-dessus du sol. Les stations sont au nombre de 24 et pour la moitié d'entre elles, un prélèvement du biofilm permet l'analyse taxonomique et le comptage des cellules constituant le microphytobenthos. Une seconde campagne, en novembre 2001, sur le même site, a permis de suivre le comportement migratoire du microphytobenthos au sein des premiers millimètres du sédiment. Le spectroradiomètre est stabilisé pour la durée du suivi, le détecteur (une fibre optique) étant placé à 30 cm au-dessus du sol. Le signal spectral du biofilm est enregistré à intervalles de quelques minutes entre l'heure de la marée basse (10h30 U.T., coefficient de 89) et l'arrivée du flot (12h06 U.T.). Chaque spectre est ensuite standardisé à sa moyenne entre 400 et 1000 nm, pour éliminer les variations d'intensité lumineuse dues à une couverture nuageuse très variable ce jour-là. Les spectres sont présentés associés à un spectre correspondant à de la vase sans développement de biofilm microphytobenthique (Fig. 3).
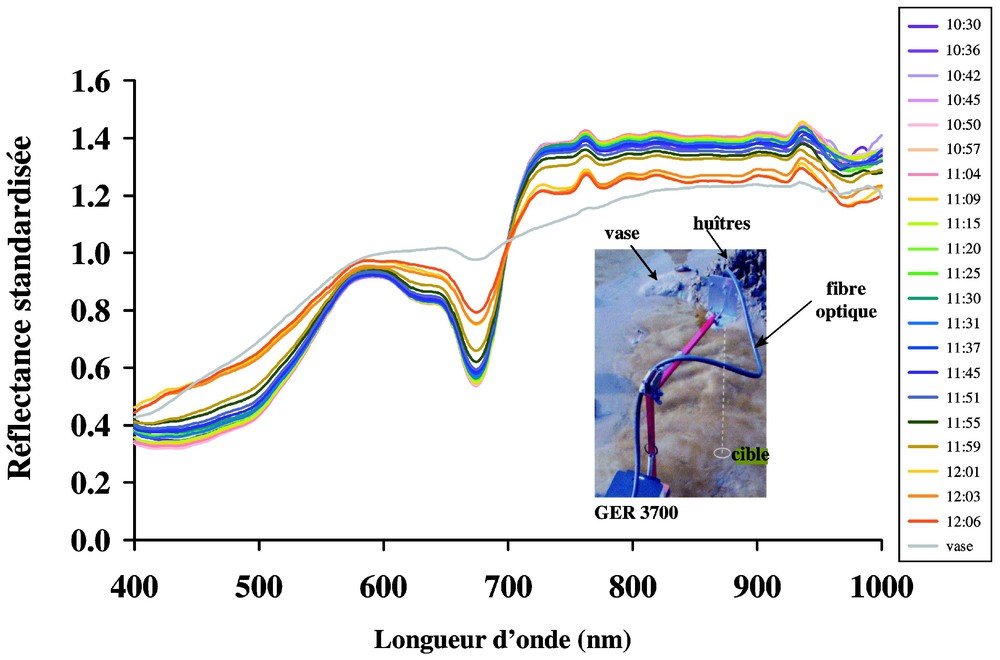
Suivi de la migration verticale du microphytobenthos dans les premiers millimètres de sédiment, avec le spectroradiomètre, en novembre 2001. Les mesures ont été réalisées entre l'heure de la basse mer (10h30 U.T.) et le retour du flot (12h06 U.T.), à intervalle de quelques minutes. Chaque spectre est standardisé à sa moyenne entre 400 et 1000 nm. Le spectre de vase correspond à de la vase sans biofilm microphytobenthique, mesuré à proximité. La photographie montre la fibre optique reliée au spectroradiomètre de terrain, les deux sont fixes durant toute la durée de l'émersion.
3.2 Résultats
Les spectres en radiance mesurés sur le terrain sont marqués par de nombreuses bandes d'absorption dues aux composants atmosphériques (Fig. 2A), qui disparaissent lors du calcul de la réflectance (Fig. 2B). Deux bandes d'absorption dans le domaine visible caractérisent la signature spectrale du biofilm microphytobenthique. Une principale, centrée sur 673 nm, correspond à l'absorption spécifique de la chlorophylle a (présente chez tous les végétaux) et de ses produits de dégradation. Une seconde, centrée sur 632 nm, est due à la présence de chlorophylle c, en plus de la chlorophylle a (Fig. 2B). Entre 400 et 500 nm, l'absorption marquée du rayonnement est peu discriminante, puisque résultant de l'absorption de l'ensemble des pigments photosynthétiques et accessoires. L'association pigmentaire chlorophylle a et chlorophylle c est caractéristique au sein du microphytobenthos de la classe des Diatomées. Ceci est confirmé par l'analyse taxonomique et le comptage des cellules des microalgues composant le biofilm, qui indique la présence majoritaire à 99 % de cellules de diatomées, dominées par les taxons Staurophora amphioxys (23 % du nombre total de cellules comptées), Pleurosigma aestuari (20 %) et Plagiotropis vitrea (15 %). Le spectre correspondant à de la vase sans développement de biofilm, enregistré à proximité, ne montre pas de bandes d'absorption marquées (Fig. 2B). Les macroalgues de la classe des Phaeophycées, communes en baie de Bourgneuf et fixées sur des substrats durs, possèdent également de la chlorophylle a et de la chlorophylle c. Cependant, leurs bandes d'absorption à 632 et 673 nm ne sont pas aussi marquées. De plus, ces végétaux ont une réflexion dans l'infrarouge proche (Fig. 2B) qui peut être utilisée pour distinguer les macrophytes du microphytobenthos lors du calcul d'un indice de végétation à partir des bandes spectrales de SPOT (cf. § 4.1.2).
Le suivi temporel de la réponse spectrale du biofilm entre l'heure de la basse mer et le retour du flot sur la vasière montre que le signal reste constant pendant la quasi-totalité de l'émersion. Ainsi, la cartographie du microphytobenthos par étalonnage d'images spectrales n'est pas limitée par le phénomène de migration verticale. En effet, une diminution rapide de l'amplitude du signal n'apparaı̂t que 7 min seulement avant le retour de l'eau sur le site d'étude (entre 11h59 et 12h06 U.T., Fig. 3). La disparition totale des bandes d'absorption à 632 nm, due aux chlorophylles a et c et entre 400 et 500 nm, indique que toutes les cellules en bon état physiologique migrent en profondeur. La bande d'absorption à 673 nm, quant à elle, ne disparaı̂t pas totalement, ce qui correspondrait à la présence de cellules en mauvais état physiologique ou mortes, qui resteraient en surface. En effet, les produits de dégradation de la chlorophylle a (chlorophyllide et phéopigments) participent aussi à l'absorption à 673 nm.
4 Analyse multi-temporelle des images SPOT
4.1 Méthodologie
L'analyse porte sur six images multispectrales (pixels de résolution 20 m×20 m) acquises pendant des marées basses entre 1986 et 1998, à la faveur de marées de vives-eaux (Tableau 1). Ces images couvrent les périodes estivale et automnale. Dans la baie de Bourgneuf, comme sur l'ensemble du littoral atlantique français, ces retraits de forte amplitude culminent aux alentours du midi solaire, heure qui coı̈ncide avec celle du passage du satellite SPOT.
Dates et heures d'acquisition des six images SPOT, avec les heures et les coefficients de marées correspondants
| Date | Heure U.T. | Heure U.T. | Coefficient de |
| d'acquisition | marée basse (B.M.) | marée | |
| 09/06/1986 | 11h40 | 10h55 | 71 |
| 28/08/1991 | 11h36 | 11h36 | 90 |
| 20/08/1993 | 11h33 | 11h30 | 111 |
| 16/09/1996 | 11h33 | 11h45 | 87 |
| 18/10/1997 | 11h20 | 11h15 | 111 |
| 21/09/1998 | 11h32 | 10h19 | 89 |
4.1.1 Étalonnage en radiance et calcul des réflectances au sol
Pour corriger les variations d'intensité lumineuse des images (exprimées en Comptes Numériques, CN) il faut les ramener à des conditions identiques d'éclairement. A défaut de pouvoir proposer un étalonnage absolu par des mesures in situ contemporaines à chaque image, un étalonnage relatif des images a été réalisé à partir de mesures radiométriques de terrain obtenues en septembre 1999. Sur le principe de l'étalonnage par empiric line correction [22,23], des références blanches et noires ont été échantillonnées sur l'image et sur le terrain : respectivement du sable sec situé en haut de plage et de l'eau de mer non turbide. La référence noire utilisée correspond au bassin d'eau de mer situé sur la commune de la Bernerie-en-Retz (2°08′W ; 47°04,7′N, Fig. 1). Sur la Fig. 4 sont représentés les spectres en radiance des références blanche et noire mesurés sur le terrain avec le spectroradiomètre, ainsi qu'un spectre intermédiaire de vase sans biofilm. Ces spectres sont dégradés à la résolution spectrale de SPOT (Fig. 4) afin de transformer les CN des images à l'aide de l'équation suivante (voir définition des variables du Tableau 2) :
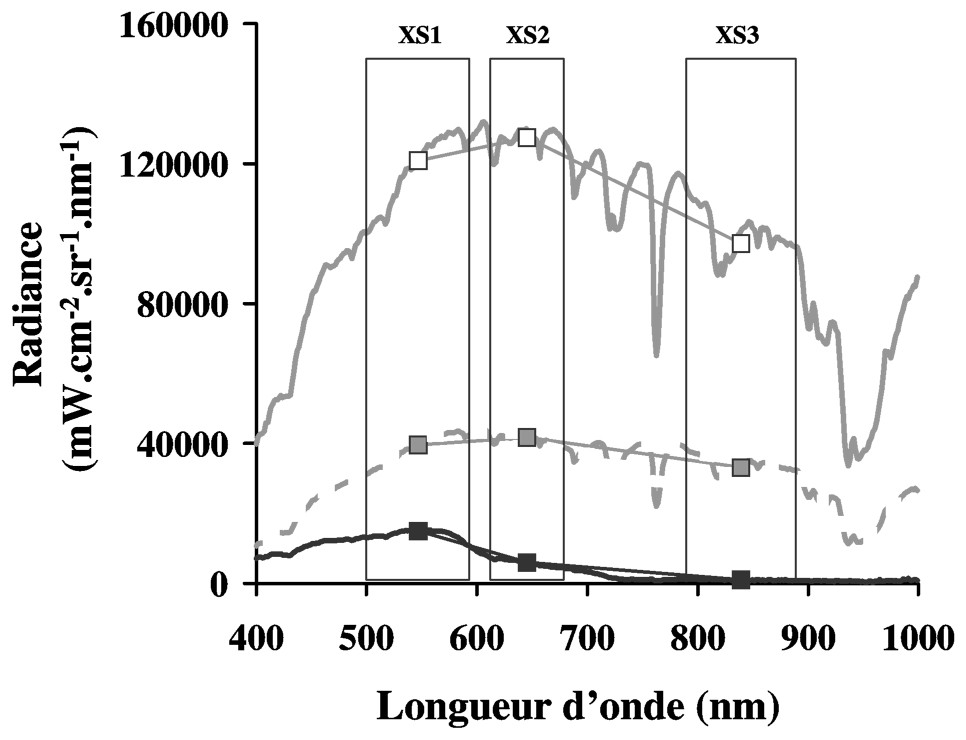
Étalonnage des images : mesures des radiances sur le terrain en septembre 1999. En noir, la référence noire, l'eau de mer non turbide (bassin d'eau de mer de la Bernerie-en-Retz) ; en gris, la référence blanche, sable de haut de plage ; en tiretés, spectre de vase sans biofilm microphytobenthique. Les carrés représentent leurs spectres dégradés à la résolution SPOT (3 canaux : XS1, XS2 et XS3), utilisés pour l'étalonnage des images. En noir, la référence noire ; en blanc, la référence blanche ; en gris, spectre de vase.
Variables des équations
| Variables | Définition |
| CN(Xn,Yn,XSi) | intensité du pixel noir de référence de coordonnées Xn, Yn du canal XSi |
| CN(Xb,Yb,XSi) | intensité du pixel blanc de référence de coordonnées Xb, Yb du canal XSi |
| CN(X,Y,XSi) | intensité d'un pixel quelconque de coordonnées X, Y du canal XSi |
| R(b,XSi) | radiance ou luminance du site blanc de référence pour le canal XSi, dégradée à partir des spectres continus de terrain |
| R(n,XSi) | radiance ou luminance du site noir de référence pour le canal XSi, dégradée à partir des spectres continus de terrain |
| R(X,Y,XSi) | radiance du pixel corrigé de coordonnées X, Y du canal XSi |
| R(S,XSi) | radiance ou luminance du Spectralon de référence pour le canal XSi, dégradée à partir des spectres continus de terrain |
| ρ(X,Y,XSi) | réflectance d'un pixel quelconque X, Y du canal XSi |
4.1.2 Principe de télédétection du microphytobenthos
À partir des images en réflectance, l'indice de végétation normalisé NDVI [24], variant entre −1 et +1, a été calculé (Tableau 2) :
Le Tableau 3 récapitule les valeurs du NDVI utilisées dans les règles de décision permettant d'identifier le microphytobenthos par rapport aux autres éléments des images. Sur le domaine intertidal, les valeurs négatives du NDVI correspondent à l'eau de mer, tandis que les valeurs positives renseignent sur la couverture végétale. Les valeurs de NDVI supérieures à 0,3 signalent la présence de macroalgues. Le microphytobenthos se situe quant à lui dans une gamme étalée de 0 à 0,3. Cependant une confusion est possible avec un faible recouvrement de macroalgues. Dans ce cas, c'est la connaissance du terrain qui permet l'interprétation des images.
Valeurs de l'indice de végétation standardisé (NDVI) utilisées dans les règles de décision d'interprétation des images de répartition spatiale
| NDVI | |||
| Domaine | Valeurs négatives | Valeurs positives | |
| Inférieurs à 0,3 | Supérieurs à 0,3 | ||
| Vasière | eau de mer turbide | microphytobenthos | |
| Affleurements rocheux | eau de mer turbide | microphytobenthos ou/et macroalgues | macroalgues |
Sur les images traitées, seules les valeurs de NDVI variant entre −0,1 et 0,3 sont représentées, les autres valeurs sont exclues de l'échelle de couleurs. Afin d'interpréter les cartes de répartition spatiale du microphytobenthos en relation avec les sites de production ostréicole, les images ont été drapées sur un fond d'image SPOT panchromatique de 10 m de résolution. Par ce procédé, un même pixel couleur représentant une intensité de NDVI est divisé en 4 pixels de 100 m2, dont la couleur est étalonnée sur chacun des pixels de 100 m2. Ce procédé permet de faire apparaı̂tre sur les cartes de répartition spatiale des détails plus fins, tels que les concessions ostréicoles et de compenser en partie le manque de résolution spatiale.
4.2 Résultats
L'étalonnage des images SPOT par rapport à une même date permet les comparaisons directes entre images. Ainsi, les comptes numériques (CN), qui présentaient une grande variabilité d'une image à l'autre en raison des différentes conditions d'éclairement, sont standardisés par le calcul des radiances, puis des réflectances.
Les images SPOT ainsi étalonnées sont utilisées pour calculer l'indice de végétation NDVI (Fig. 5). Les règles de décision pour l'interprétation des images (Tableau 3) permettent de localiser les pixels correspondant au microphytobenthos sur la vasière. Ainsi, les peuplements microphytobenthiques représentés selon l'échelle de couleur par la couleur rouge (NDVI moyen de 0,2) apparaissent, sur la plupart des images, localisés en bas de l'estran. Ces peuplements forment des taches dont la superficie totale dépasse la centaine d'hectares. Les deux premières images, en 1986 et 1991, ne présentent pas de forts développements du microphytobenthos, occupant respectivement 6 % (soit 150 ha) et 10 % (soit 220 ha) de la surface de la vasière, comparé aux quatre autres où la superficie totale occupée par les peuplements microphytobenthiques peut atteindre 25 % (soit 550 ha) de la surface de la vasière (en 1996 et 1997). Sur ces quatre images, le microphytobenthos se développe préférentiellement le long d'une bande nord–sud suivant l'isobathe +2 m des cartes marines, particulièrement marquée en 1996 et 1997, images pour lesquelles les superficies atteintes sont les plus importantes. Sur les images de 1993 et 1998, les taches de microphytobenthos ont tendance à suivre l'isobathe +2 m, mais remontent vers le haut de l'estran, au niveau du chenal du Falleron, où la bathymétrie peu atteindre +4 m. Enfin, notons que sur les six images, quelle que soit l'importance du développement des peuplements microphytobenthiques, il y en a toujours qui sont associés aux parcs ostréicoles de la Coupelasse, au sud de la zone étudiée, ainsi que sur les parcs de la commune de la Bernerie-en-Retz, au sud du bassin d'eau de mer utilisé pour l'étalonnage des images.
À proximité du rivage et en l'absence de macroalgues, certains pixels présentent un NDVI voisin de 0,2, notamment sur les images de 1996 et 1997, au sud, en vis-à-vis des parcs de la Coupelasse, et au nord de l'embouchure du Falleron (flèches noires sur la Fig. 5). Il s'agit de peuplements microphytobenthiques dans les banquettes et seillons du haut de l'estran. En effet, ces formations sédimentaires dues à l'hydrodynamisme peuvent faire l'objet d'un fort développement microphytobenthique observable lors des sorties sur le terrain.
5 Discussion
L'utilisation d'images SPOT multispectrales, étalonnées par spectroradiométrie de terrain, a permis la cartographie des peuplements du microphytobenthos dans l'écosystème conchylicole de la baie de Bourgneuf. Les résultats ont mis en évidence une répartition spatiale de ces peuplements selon une structuration marquée en bas de la zone de balancement des marées. En effet, le microphytobenthos a tendance à se développer le long d'une bande nord–sud le long de l'isobathe +2 m des cartes marines (Fig. 5). Contrairement aux résultats obtenus par Guarini et al. [5] à Marennes-Oléron, il semblerait que la bathymétrie dans la baie de Bourgneuf soit un des facteurs qui structure la répartition spatiale des peuplements microphytobenthiques. Cette structuration apparaı̂t nettement sur les images de 1996 et 1997, qui sont les images présentant les plus fortes superficies de microphytobenthos (Fig. 5). En contraste, les images de 1986 et 1991 ne présentent qu'un faible développement microphytobenthique. En 1986, ceci peut s'expliquer par le fait que l'image ayant été prise 45 min après l'heure de la marée basse, pour un coefficient de marée de 70 (Tableau 1), le flot turbide au moment de la prise de vue (11h40 U.T.) est déjà arrivé au niveau de l'isobathe +2 m. En 1991, l'heure de la marée basse coı̈ncide avec l'heure d'acquisition de l'image (Tableau 1) et le faible développement du microphytobenthos restreint à la partie sud de la zone d'étude ne s'explique, ni par le retour du flot, ni par le processus de migration verticale.
Entre 1993 et 1998, il existe des différences de répartition du microphytobenthos entre le Sud et le Nord de la zone étudiée. Sur les images de 1993 et 1998 (Fig. 5), il se situe plus haut sur l'estran, par rapport à sa position sur les images de 1996 et 1997. Cette situation des taches microphytobenthiques peut être mise en relation avec le Falleron et son chenal (Fig. 5), exutoires du bassin de drainage, avec un débit journalier moyenné sur l'année de 500 000 m3 [25], qui peut participer à l'apport d'éléments nutritifs pour le microphytobenthos. Cet exutoire semble donc être un des facteurs pouvant intervenir dans la structuration de ces peuplements.
Sur les six images, la localisation du microphytobenthos au niveau de certaines concessions ostréicoles suggère une relation entre ces peuplements et les cultures d'huı̂tres, particulièrement marquée au niveau des parcs de la Coupelasse (Fig. 5). Ce résultat est compatible avec l'hypothèse d'un processus rétroactif permettant aux microalgues benthiques d'utiliser les produits de l'excrétion dissoute des bivalves et ceux issus de la minéralisation de leur biodépôts, pseudofèces et fèces [25–28].
Le calibrage des images SPOT a permis de mettre en évidence les limites liées aux résolutions spectrale et spatiale du capteur Haute Résolution Visible (HRV), pour l'étude spécifique du microphytobenthos. S'il s'avère parfois difficile de différencier microphytobenthos et macroalgues à faible couverture à partir des trois bandes spectrales XS1, XS2 et XS3, la connaissance du terrain permet de lever le doute concernant certains pixels. En effet, sur la vasière entre le sud de la Bernerie-en-Retz et le chenal du Falleron, il y a très peu d'affleurements rocheux, ce qui limite ainsi la présence des macroalgues. Cette connaissance sédimentologique associée à des observations de terrain confirme l'identification du microphytobenthos sur les cartes de répartition spatiale (Fig. 5). Cependant, près des parcs ostréicoles de la Coupelasse, où les affleurements rocheux sont plus nombreux, la signature spectrale des pixels peut résulter d'un mélange de microphytobenthos et de macrophytes. La Fig. 6 présente ainsi la superposition d'un spectre de terrain de biofilm de microphytobenthos avec celui obtenu sur un affleurement rocheux, faiblement recouvert de fucus et entouré de vase. Les deux spectres mesurés par le spectroradiomètre, dégradés à la résolution spectrale de SPOT, ne peuvent pas être discriminés. En effet, la résolution spectrale trop large du capteur HRV ne permet pas d'exploiter la spécificité des bandes d'absorption des chlorophylles a et c (Fig. 2A), qui se retrouvent englobées dans la bande XS2 (610–680 nm) (Fig. 6).
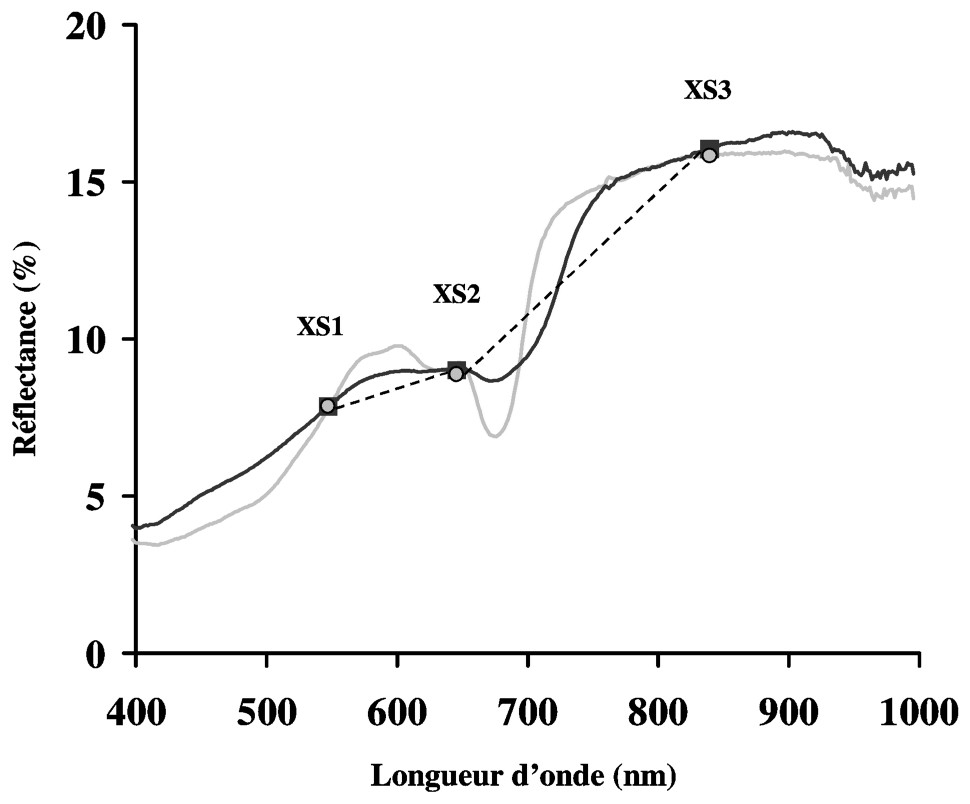
Comparaison de spectres continus de terrain associés aux réflectances dégradées à la résolution SPOT (XS1, XS2 et XS3). En noir (ligne et carrés) : affleurement rocheux, faiblement recouvert de fucus et entouré de vase ; en gris (ligne et points) : vase recouverte d'un biofilm de microphytobenthos.
En ce qui concerne la résolution spatiale de SPOT, les pixels de 400 m2 portent une information qui intègre la variabilité de la répartition spatiale du microphytobenthos à microéchelle. En effet, le microphytobenthos présente in situ une répartition centimétrique en taches colorées très inférieure à la résolution spatiale d'un pixel de l'image SPOT. Les couvertures maximales mesurées sur le terrain avec le spectroradiomètre seront donc rarement retrouvées sur une image. La mise en service de SPOT 5, qui possède une plus forte résolution spatiale (10 m pouvant être ré-échantillonnés à 2,5 m), devrait permettre d'obtenir des résultats plus précis à l'échelle de pixels de 100 m2 (subdivisés en pixels de 6,25 m2).
Enfin, une perspective réaliste de ce travail est l'obtention de cartes de biomasse microphytobenthique, à partir d'images hyperspectrales. De telles cartes seront la résultante de la relation mathématique entre l'information physique qu'est la signature spectrale (réflectance) et l'information biologique qu'est la biomasse (mg de chlorophylle a par mètre carré). En effet, nous avons vérifié que le processus de migration verticale des microalgues n'est pas une limite à l'estimation de la biomasse (Fig. 3). Les mesures de terrain permettent de confirmer l'hypothèse de Guarini et al. [5,8], basée sur les travaux de Serôdio et al. [12], postulant que la biomasse totale responsable de la photosynthèse se trouve à la surface du sédiment durant toute une marée basse diurne. Les cellules en bon état physiologique migrent en profondeur quasi instantanément au retour du flot, la faible absorption encore détectable à 673 nm étant due à la chlorophylle a et à ses produits de dégradation dans les cellules sénescentes ou mortes restant à la surface du sédiment.
Face à ces contraintes liées aux résolutions spatiale et spectrale, l'utilisation de spectromètres imageurs mesurant les radiances en mode hyperspectral (d'une dizaine à plusieurs centaines de bandes), tels CASI, AVIRIS, DAIS et ROSIS, s'avère alors indispensable dans le cadre de l'étude du microphytobenthos. Ils permettraient une distinction satisfaisante des macroalgues et du microphytobenthos [29], mais également au sein même des peuplements microphytobenthiques, sur la base des compositions pigmentaires, une discrimination éventuelle des classes algales : Cyanophycées, Chlorophycées et Diatomées [9,21]. Enfin, ce type d'image rendrait alors possible la cartographie du microphytobenthos en terme de biomasse. Ainsi, l'information sur la répartition spatiale du microphytobenthos pourra être couplée à la quantité de biomasse dont l'estimation à l'échelle de l'écosystème est une donnée essentielle dans la perspective d'analyser les flux de matière utilisables par les huı̂tres cultivées, problématique qui dépasse en portée le site d'étude de la baie de Bourgneuf et concerne la majorité des baies à vocation aquacole de la façade atlantique et de la Manche.
Remerciements
Nous remercions le conseil général de Loire-Atlantique pour l'attribution du prix de recherche Atlanthèse à Vona Méléder. Nous remercions SPOT IMAGE pour nous avoir permis d'acquérir les premières images grâce à ses programmes promotionnels de 1996 et 1998, et le programme ISIS du CNES, qui nous a permis d'acquérir les images de 1986, 1991, 1993 et 1997.
Le laboratoire de planétologie et géodynamique de Nantes remercie la région Pays de la Loire et le Programme national de planétologie (Insu–CNRS) pour leur contribution décisive à l'acquisition du spectroradiomètre GER 3700.