

Abridged English version
A study of the eye's role, and more precisely the pigmentary distribution, in the control of the rhythm of daily displacement of Daphnia longispina was carried out.
D. l. exhibits daily vertical migrations (DVM). Though reverse migrations may occur, in most cases, zooplankters more upward at dusk and downward at dawn. The movements coincide more or less with the natural changes of light intensity. They persist under constant conditions of illumination, suggesting the existence of an endogenous component occurring in the control of DVM mechanism. This component was investigated in the eye of Daphnia.
Several structural and ultrastructural modifications of the eyes of various organisms were observed between daytime and nighttime. Most of these studies were based on only two samples, one taken under darkness and one under at light. To our knowledge, no author tackled the issue of the interplay between structural modifications and behaviour of the organisms. To put in evidence such a link, we monitored the evolution of the structure of the eye of Daphnia during daily cycles of migration. We measured during 24 h the pigmentary repartition of organisms either in situ or kept under permanent light (LL) in the laboratory. Such a measurement was made by analysing pigmentary surfaces on the basis of colour differentiation.
For the in situ cycle, we observed an increase of the pigmentary surface both after dawn and dusk. These increases begun in both cases one hour after the change of illumination. They occurred simultaneously with the spatio-temporal modifications of the organisms.
The pigmentary changes observed under LL present some analogies with those observed in situ, except in their progress. In LL, they occurred prior to the ascent of Daphnia, instead of coinciding. This was the expression of an endogenous rhythm. The rhythm put in evidence was of circadian type.
We collected indications of the link between daily migrations of daphnies and structural modifications of their eyes. In D. l., an internal oscillator intervenes in the control of the DVM. In such a process a structure must intervene by receiving the alternation day/night information and by triggering the descent of the organisms at dawn and their coming back up at dusk. Therefore migration of Daphnia seems of a nycthemeral type. In situ, the illumination at dawn suffices to synchronise the migrations on the day/night alternation.
1 Introduction
Les migrations verticales journalières (DVM) du zooplancton sont étudiées depuis fort longtemps. Cependant, nombre de points dans leur mécanisme ne sont pas encore clairement expliqués. Dans le but de progresser dans cette voie une thèse a été menée à bien [1]. Les matériels biologiques choisis sont des Cladocères : Daphnia longispina et Bosmina longirostris. La première espèce effectue de tels déplacements dans les lacs profonds au contraire de la seconde, qui servira de témoins. Dans le présent travail il ne sera question que des daphnies.
Les migrations verticales des daphnies coı̈ncident plus ou moins avec les changements naturels d'intensité lumineuse. Les animaux montent en fin de journée et descendent au matin [2]. Des migrations tout à fait identiques quant à leur modalités peuvent être observées au laboratoire [3–5]. Cette coı̈ncidence fait que, dans la plupart des études, essentiellement phénoménologiques, les auteurs considèrent que les variations d'éclairement jouent le rôle de déclencheur des déplacements des animaux. Mais ces déplacements persistent en conditions constantes d'éclairement [2,4,6,7]. Ceci suggère l'existence d'une composante interne intervenant dans le contrôle du mécanisme des DVM chez Daphnia [3,5,8,9]. Or, une telle structure n'exige qu'un seul signal lumineux par 24 h pour se synchroniser sur l'alternance jour/nuit [10].
Le rôle de la lumière paraissant prouvé [7,11–16], on peut penser que les photorécepteurs sont impliqués dans ce contrôle.
Plusieurs modifications structurales et ultrastructurales des yeux ont été observées entre le jour et la nuit chez divers organismes [17–21]. Aucune n'a été mise en relation avec une modification comportementale, a fortiori migratoire. Pour mettre en évidence un tel lien, nous avons suivi l'évolution de la structure de l'œil de Daphnia au cours de cycles journaliers de migration. Après avoir montré un lien entre l'œil et l'horloger interne, en l'occurrence une variation de la répartition des granules pigmentaires des cellules neurosensorielles coı̈ncidant avec la diminution de l'éclairement, mais se maintenant en conditions constantes d'éclairement (LL) [22]. Nous nous proposons de vérifier ici trois points. Tout d'abord que les variations de la densité pigmentaire intéresse l'ensemble de chaque ommatidie, ce qui suppose une alternance de synthèses et de dégradations. Puis que ces variations suivent le même rythme et sont en phase avec le DVM et enfin qu'elles se maintiennent, comme les DVM, en conditions constantes d'éclairement (LL).
2 Matériel et méthodes
Les daphnies (Daphnia longispina) sont prélevées dans la retenue de Grangent, sur la Loire.
Dix prélèvements à la bouteille fermante aux profondeurs −5 et −20 m et espacés de 2 à 3 h sur un nycthémère permettent les observations in situ. Les organismes sont immédiatement anesthésiés à l'eau de Seltz, puis fixés.
Ceux destinés à l'expérimentation au laboratoire sont prélevés le même jour au filet Juday. Ils sont transportés dans de grands conteneurs remplis d'eau du lac. Au laboratoire, les daphnies sont introduites sans délai dans deux colonnes en altuglas, situées dans des salles climatisées où l'on accède par un sas. L'ensemble du dispositif est décrit en détail par Berthon et Brousse [5] et Cellier et al. [22]. Les deux colonnes (3×0.25 m) sont éclairées par un générateur de lumière froide (Schott KL 1500), contrôlé par un programmateur. L'éclairement correspond à une intensité lumineuse de .
Après 48 h d'acclimatation, les animaux sont placés en éclairement constant (LL). L'aube et le crépuscule subjectifs sont fixés respectivement à 7 h 00 et 21 h 00. Quelques individus sont ensuite régulièrement prélevés dans chaque colonne, durant 3 jours environ. On utilise pour cela un petit conteneur rempli d'eau de Seltz, relié à une pompe à vide à 17, 20, 22, 1, 4, 6, 8, 11 et 14 h et immédiatement fixés [1].
Après préparation et inclusion dans du SPURR, des coupes sagittales de la région de l'œil sont pratiquées. L'épaisseur choisie – 400 nm – correspond au diamètre moyen des grains de pigments. Les coupes sont observées en microscopie photonique et les vues traitées en analyse d'images (Logiciel Samba 2005).
L'estimation de la densité numérique des granules pigmentaires contenus dans chacune des huit cellules rétiniennes entourant l'axe des ommatidies est effectuée en analysant leur surface sur la base d'une différence de coloration. Pour cela, la division naturelle de chaque ommatidie en 3 zones est utilisée (Fig. 1). La première zone contient le cristallin, la seconde est immédiatement en dessous. La ligne séparant les zones 2 et 3 se situe au niveau de la petite constriction du rhabdome. La dépendance des données impose une analyse de variance multidimensionnelle (MANOVA) pour examiner l'évolution des répartitions pigmentaires en fonction du temps et des conditions d'éclairement. Parallèlement, une analyse de variance (ANOVA) de la somme des granules pigmentaires des trois zones en fonction du temps est effectuée.

Principe de la division des ommatidies en trois zones.
La réalisation de ces tests requiert que les populations soient gaussiennes et de même matrice de variance–covariance, ce qui est vérifié.
Les aires délimitant les zones analysées sont similaires et ne présentent donc pas de variations.
3 Résultats
Les résultats présentés ne portent que sur les 24 premières heures d'observation. En effet, bien que le patron mis en évidence se reproduise les jours suivants, sans dérive apparente en conditions constantes d'éclairement, le taux de mortalité élevé des daphnies dans les colonnes ne permet pas de traiter les données de façon pertinente au delà de 28 à 30 h.
Lors du cycle in situ (Fig. 2), les surfaces pigmentaires par zone et la somme des pigments présentent des différences significatives en fonction du temps (Tableaux 1 et 2). L'aube et le crépuscule sont suivis d'une augmentation de la surface pigmentaire débutant dans les deux cas une heure après le changement d'éclairement. Ces modifications de l'œil coı̈ncident donc avec celles de la répartition spatio-temporelle des organismes.
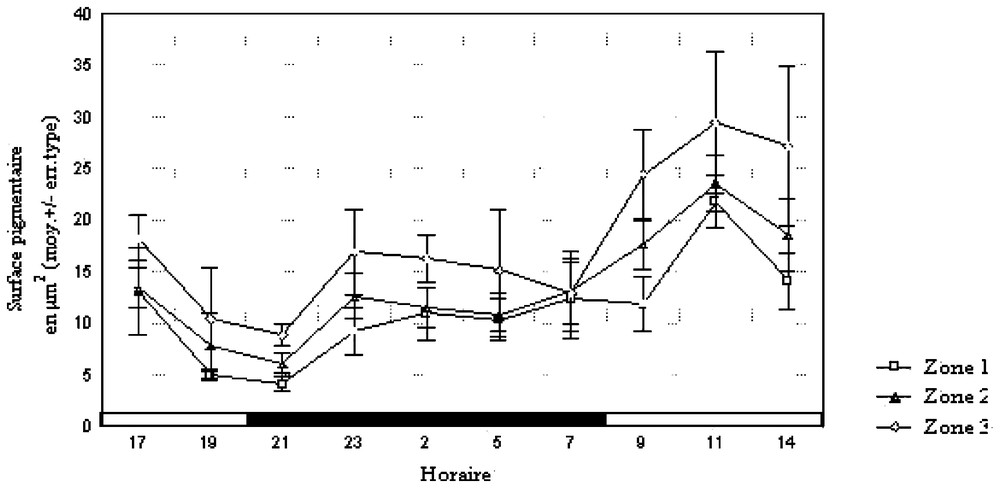
Évolution de la surface pigmentaire dans les différentes zones en conditions naturelles.
Table de MANOVA pour les différentes zones dans ommatidies, en conditions naturelles (in situ) et en conditions constantes d'éclairement (LL blanc)
| Valeur de la trace | P | |
| de Lawley–Hotelling | ||
| in situ | 1,784 | 0,0379 |
| LL | 1,515 | 0,0134 |
Table d'Anova pour la pigmentation en conditions naturelles (in situ) et en conditions constantes d'éclairement (LL blanc)
| DF | Somme des carrés | Carré moyen | P | ||
| in situ | heures | 9 | 9264,880 | 1029,431 | 0,0102 |
| résidus | 29 | 9689,773 | 334,130 | ||
| LL | heures | 8 | 2790,024 | 348,753 | 0,1017 |
| résidus | 34 | 6412,892 | 188,614 |
En conditions constantes d'éclairement (LL), la MANOVA indique qu'il y a un effet horaire pour les surfaces pigmentaires (Tableau 1). Les différences sont significatives lorsqu'on considère la somme des pigments des trois zones (Tableau 2). Les moyennes des surfaces pigmentaires présentent deux maxima, 15,61 et 13,51 μm2, à 22 et 14 h, et deux minima, 3,35 et 6,35 μm2, à 8 et 17 h (Fig. 3).

Évolution de la surface pigmentaire dans les différentes zones en conditions constantes d'éclairement (LL blanc).
4 Discussion
Contrairement à Röhlich et Törö [23], qui ne notent pas de différence dans la répartition des pigments de l'œil chez Daphnia entre les animaux adaptés à la lumière et ceux adaptés à l'obscurité (en limitant leur étude à une seule observation ponctuelle par état), nous constatons une modification significative de la répartition des granules pigmentaires au cours du nycthémère, tant dans les observations au laboratoire que sur le terrain. Les résultats obtenus in vitro en cycle LD et sur le terrain sont tout à fait comparables [1]. On peut donc considérer que les conditions expérimentales ne modifient pas le comportement migratoire des daphnies. C'est pourquoi nous comparons dans cette discussion les observations en conditions naturelles et en cycle LL au laboratoire.
Ainsi, on peut distinguer des périodes de synthèse et de dégradation des pigments comme cela est signalé chez Leucophaea maderae par Ferrell et Reitcheck [24]. Ces changements paraissent induits par les variations d'éclairement, dans la mesure où ils coı̈ncident avec l'aube et le crépuscule. Toutefois, deux problèmes se posent pour accepter cette déduction. D'une part, comment expliquer que l'illumination produise le même effet que l'extinction sur l'ultrastructure des cellules rétiniennes des ommatidies (en l'occurrence une synthèse de pigments), lumière et obscurité ayant des actions opposées s'agissant des déplacements ? D'autre part, si la position des daphnies près de la surface permet une action de la lumière à l'aube, on peut douter qu'elles perçoivent l'extinction progressive crépusculaire alors qu'elles sont à plus de 20 m de profondeur (des mesures effectuées à l'aide d'une cellule photoélectrique immergeable (268WA310 Kalsico Int. Corp.) indiquent que, durant la période où les daphnies migrent, la limite Z1 %° varie entre −16,5 et −20,8 m). Au mieux l'intensité de la lumière alors perçue est extrêmement faible. Dans ces conditions le seuil de variation permettant de déclencher la réaction des daphnies n'est sûrement pas atteint. Nous pensons donc que si la lumière a une action sur la répartition des pigments de l'œil, au voisinage du crépuscule, ce ne peut être qu'à la fin d'un long processus biologique débutant à l'aube et en aucun cas une action directe. Dans ce processus doit intervenir une structure qui reçoit l'information de l'alternance jour-nuit et déclenche la descente des organismes au lever du jour et leur remontée au crépuscule, faisant que la migration des daphnies semble de type nycthéméral.
Par ailleurs, l'évolution de la densité numérique des granules pigmentaires présente des cinétiques voisines en conditions naturelles d'alternance jour–nuit et en éclairement constant. Cependant, si en LL on observe une augmentation pigmentaire, une heure après l'aube subjective, identique à celle observée in situ, celle qui suivait le crépuscule in situ, précède, en LL le crépuscule subjectif d'environ 3 h.
Or, si les DVM sont maintenues en conditions constantes d'éclairement, on observe que la montée des organismes précède souvent le crépuscule subjectif au lieu de coı̈ncider avec lui [4], indiquant ainsi que la variation de l'éclairement crépusculaire ne déclenche pas cette remontée.
Donc un schéma proche, mais un rythme différent. Tout se passe comme si en LL se manifestait le rythme propre d'une composante interne placée en libre cours, c'est-à-dire qui n'est plus synchronisée par le signal extérieur approprié.
Le rythme mis en évidence est de type circadien. L'absence de dérive du phénomène après trois jours d'observation en conditions constantes (LL) indique même que ce rythme doit être très proche de 24 h. Or, il concerne des cellules neurosensitives, considérées par les chronobiologistes comme des oscillateurs potentiels [25–27]. L'œil pourrait à lui seul assurer le contrôle du rythme des déplacements.
5 Conclusion
La mise en évidence, par l'expression de son rythme propre, d'une composante interne intervenant dans le mécanisme des migrations fait de celles-ci des phénomènes circadiens, mais permet aussi de comprendre leur apparence nycthémérale. En effet, le rythme circadien de l'oscillateur interne ne se manifeste qu'en libre cours. En conditions naturelles, un tel oscillateur se synchronise sur l'alternance jour-nuit, dès lors qu'il perçoit cette alternance (comme ici à l'aube lorsque les daphnies sont près de la surface). Le rythme du phénomène contrôlé devient nycthéméral. Or, une telle structure ne peut qu'être génétiquement déterminée. Dans ces conditions, les migrations verticales des daphnies pourraient avoir une signification beaucoup plus générale qu'une simple réponse adaptée d'une population à une condition locale défavorable, comme la présence d'un prédateur. Ces migrations pourraient être l'un des éléments essentiels des stratégies écologiques chez les Cladocères, indispensable notamment à la constitution de réserves énergétiques [4]. C'est ce qui expliquerait que, dans certaines populations, le patron des migrations ne s'établit que lorsque le plan d'eau est stratifié [16].
Remerciements
Nous remercions M. Réhaı̈lia pour sa participation aux traitements mathématiques.