

1 Introduction
The symmetrical or asymmetrical nature of the gaits used by quadrupeds was emphasized in 1966 by Hildebrand, who based his definition on the phase relationship between fore and hindlimbs footfalls. The gait diagram that he proposed to specify symmetrical gaits is founded on two variables: ‘the duration of contact of each foot expressed as percentage of the stride interval’ (called duty factor by Alexander [1]), and the ‘percent of the stride interval that the footfall of a forefoot lags behind the strike of the ispilateral hind foot (the hind foot on the same side of the body)’. These variables are respectively plotted on the abscissa and ordinate axes of a graph, the origin on each axis being at the upper right corner. The onset of the cycle (called stride interval) is by convention that of one hindlimb. From 0 to 50%, the limbs phase indicates what is called lateral sequence, because the sequence of the limbs action within a single cycle is kept on one side of the body (hind, then fore of the same side). When the phase is over 50%, the forelimb situated on the other side of the body lands before that which is on the same side as the reference hindlimb. This is called the diagonal sequence, because the forelimb following the hindlimb is set on the diagonal. Furthermore, percent of the total stride interval during which an animal was supported only by diagonally opposite feet is indicated on the graph by concentric squares. The record of several hundred of gait formulae for 156 genera of terrestrial vertebrates permitted to outline an area for natural gaits. Hildebrand indicated that the proposed diagram was valid only if the anterior contacts have the same duration as the posterior contacts and he gave adjustments for exceptions to this rule. Such exceptions were noticed in salamanders and lizards, which often show a shorter time of contact for the fore limbs [2], or for the hindlimbs in running crocodiles [3]. Thus, these irregularities may illustrate natural cases according to peculiar body geometry that results in specific limbs coordination. The Hildebrand's grid system provided a powerful tool for comparison of the gaits performed by diverse four-footed animals, giving a quantified definition of gaits terminology [4].
2 Results
By convention, Hildebrand chose one hindlimb as reference to calculate footfalls variables. However, in experimental conditions, it may often happen that the forelimb stride only can be completely recorded. In such cases, the Hildebrand's model can be also applied as follows. Let us take from [5] (Fig. 3-2) the examples of the pigmy hippopotamus, characterized by a single foot in lateral sequence, and the monkey, characterized by a trotting walk in diagonal sequence. A simple horizontal shift in the associated footfalls diagram changes the reference limb from the left hindlimb to the left fore limb (Fig. 1). Consequently, the percentage that hind footfall (LH) follows fore (LF) on the same side becomes 80 in the case of the pigmy hippopotamus and 40 in the case of the monkey. The gait is plotted in the same position within the graph if we reverse the ordinate axis. One hundred percent has to be placed on the ordinate at the lower left corner of the graph when the reference limb is the hind one, whereas it has to be placed at the upper left corner when a forelimb is used. There is no change in the location of the lateral and diagonal sequences on the graph. Of course, the change of reference does not affect the sequence of limbs action. It may be noticed that a forward lateral succession and a backward diagonal crossing occur in a lateral sequence, whereas a backward lateral succession and a forward diagonal crossing occur in a diagonal sequence. The terms used by [6] and also by [7] are based on this observation and may bear sense for dynamic consideration.
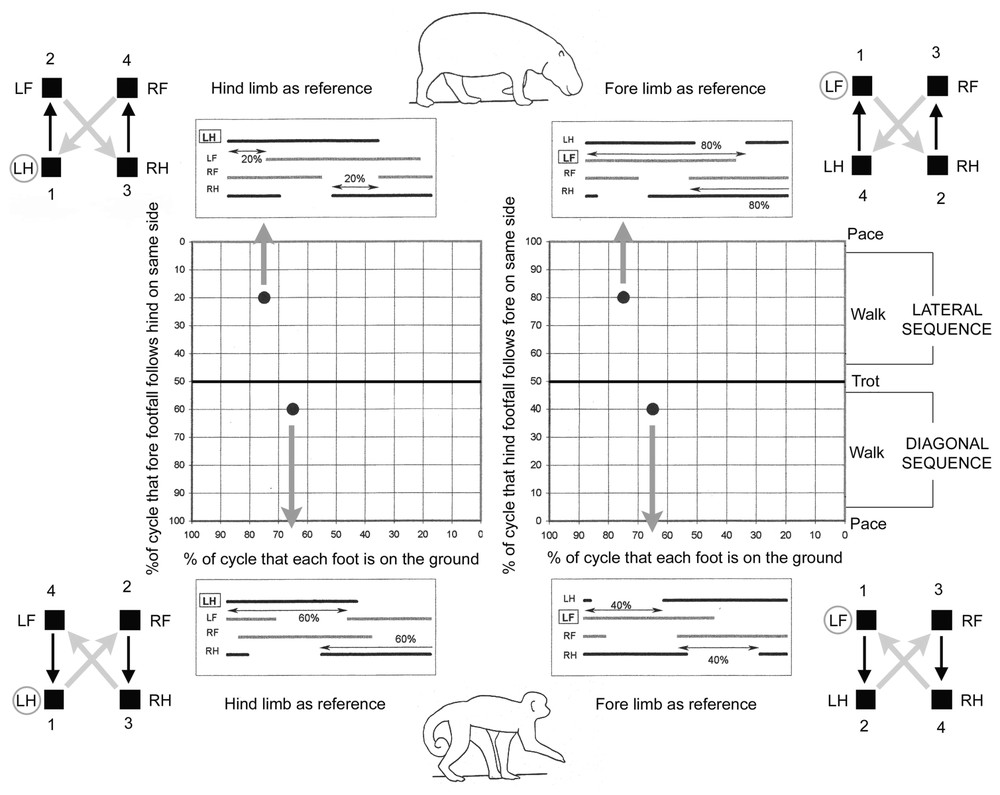
Examples taken from Hildebrand 1985: walk in lateral sequence for a pigmy Hippo and walk in diagonal sequence for a monkey. On the left side, conventional graphs obtained when the hindlimb is taken as reference (number one in the series of limbs actions). Footfall pattern of each animal permits the determination of the gait position on the Hildebrand graph. On the right side, one forelimb is taken as reference (number one in the series of actions). The simple reversal of the scale on the vertical axis demonstrates that the gaits of the animals have identical position on the Hildebrand graph.
We suggest a second addition to the Hildebrand model concerning asymmetrical gaits. He specified these gaits as follows [8]: “asymmetrical gaits have the footfalls of the two feet of a pair (fore or hind) unevenly spaced in time”, and in contrast for symmetrical gaits “since symmetrical gaits have the footfalls of a pair evenly spaced in time, the particular instant a given foot strikes always comes midway between two footfalls of its opposite”. For instance, the strike of a right hindlimb happens at 50% of the left hind cycle. The same time lag occurs between the right and the left forelimbs. These two variables, time lag between the footfalls of a pair of fore limbs and time lag between the footfalls of a pair of hindlimbs, can be plotted on a graph (Fig. 2). The points located at 50% on each axis (±5%) correspond to symmetrical gaits, and all those that are aside the range have to be considered as asymmetrical. Such a complement to the Hildebrand method could discriminate on the same graph the basic gait patterns of different species. The analysis of asymmetrical gaits necessitates peculiar variables [9]. To characterize the two kinds of flying phases when no limbs are on the ground that can appear in gallops, was used percent of stride interval that fore midtime (middle of the contact time by one or both forelegs) follows hind midtime, related to percent of stride interval that the body is supported by one or both hind feet. Another important feature is the lead sequence that is the time lag between the footfalls of the limbs within a pair. When a moderate fore lead (between the trailing and the leading fore limbs) appears the gait become a half bound. In the bound, forelimbs hit the ground together (no lead), followed by a flight phase before the hindlimbs hit also the ground together. In a gallop, the sequence of footfalls is transverse when the fore feet had the same lead as the hind (RF first and LF; RH first and LH). It is rotary when they had an opposite lead (RF first and LF; LH first and RH). As important character of diverse asymmetrical gaits, lead sequence can be plotted on a graph (Fig. 3). The hind lead (positive when the trailing hind leg is the same as the trailing fore leg, or negative when the trailing hind leg is opposite to the trailing fore leg) is indicated on the X-axis in percent of the stride interval; the fore lead is plotted on the Y-axis also in percent of the stride interval. Rotary gallops are placed on the left part of the graph, transverse gallops on the right, half-bounds directly on the ordinate and bounds at the axis intersection.

The lead (time separating the touch down of limbs within one pair) for the fore pair plotted against the lead of the hind pair. This simple graph permits to discriminate symmetrical and asymmetrical cycles performed either by individuals in the same species at diverse velocities, or by diverse species. Of peculiar interest is to look at the pair of limbs, which initiates the passage in one of the asymmetrical quadrants in introducing a lead higher than 55% of the cycle duration (from personal unpublished data in house mouse).
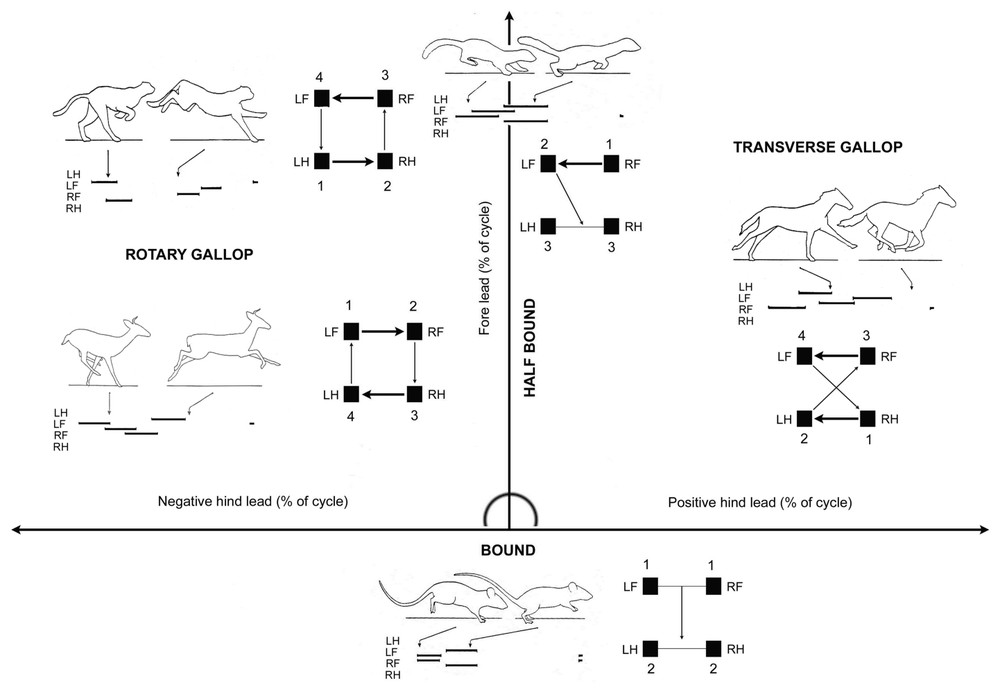
Examples taken from Hildebrand [5]: within asymmetrical gaits, the leading limb can be on the same side in both pairs (positive hind) or on opposite sides (negative hind). The absence of hind lead characterizes half-bound, and no lead in both pairs, the bound. Thus any kind of asymmetrical gait recorded can be plotted on one of the three axes of the graph.
Gaits were usually analysed to characterize behavioural differences between taxa, and the Hildebrand grid system, based on quantitative parameters, placed this descriptive phase on sound basis. However, neuromuscular control of locomotion has also been intensively studied. There is now a demand to understand the complex organization, which underlies the diverse resulting gaits. A more integrative approach, compiling biomechanical, neurophysiologic, behavioural and ontogenetic data, is necessary [10]. Comparative locomotion studies are now questioning how animal's motion takes its place in the life history, relating form, function and evolutionary strategies. This implies the use of analytical tools, which can be scaled in correspondence: for instance, kinematic parameters have to agree with neurophysiological time scale.
Acknowledgements
We thank J. Davenport for his help and his judicious comments in the English revision of the manuscript.