

Abridged English version
The Eupelmid Eupelmus vuilleti occurs in the Sahel region of Africa, in a parasitic community that includes two other parasitoids (another Eupelmid E. orientalis, and a Pteromalid Dinarmus basalis). Eupelmus vuilleti is a primary, solitary ectoparasitoid of the larval stages of the Bruchids Callosobruchus maculatus and Bruchidius atrolineatus. In a context of intense competition for healthy hosts, E. vuilleti displays ovicide and larvicide behaviours towards the Pteromalid D. basalis during its development (kleptoparasitism), and in an extreme expression of kleptoparasitism, the E. vuilleti females oviposit within the final larval stage (L5 stage) of D. basalis.
In this study, the adaptation capacity of the males developed on secondary hosts when 24 h old was determined from the quantity of spermatozoids stored in the seminal vesicles and the quality of insemination determined by the quantity of spermatozoids stored in the spermatheca of the females after a single mating, and latter was estimated by the number of daughters produced. However after having developed as hyperparasitoids, the numbers of daughters which reveals the ability of the males to inseminate the females, provides a way of determining some aspects of the reproductive success of females via the migration of the spermatozoids to the reservoir of the spermatheca, and by the mobilization of these spermatozoids at the moment of the fertilization of an oocyte.
In parasitoids, in which the reproductive system involves arrhenotoky parthenogenesis, the selection of hosts and the sex of the descendants are often correlated to the size of the host involved, taken to be an indicator of its quality. However, after developing in the context of hyperparasitism, the size of the adults E. vuilleti is miniaturized and measured by the fresh weight when they emerge; this size is on average significantly lower than that of adult males and females developed on the primary host C. maculatus.
The miniature male E. vuilleti that hatch out from a secondary host retained the ability to mate, particularly as the gregarious nature of the hosts promoted mating at the emergence site.
The ability of the females to produce daughters was also assessed from the size of the stock of spermatozoids in the reservoir in their spermatheca. However, despite the significant differences between the production of spermatozoids in males depending on their origin (in the males that had developed in a context of hyperparasitism, the counts ranged from 600 to 3000 spermatozoids, whereas the counts in males that had developed as primary parasitism ranged from 1800 to 4200 spermatozoids), it was noteworthy that after the first mating, the potential numbers of spermatozoids that could be mobilized by the females to produce daughters were not significantly different whatever the origin of the females: on average spermatozoids were stored by the hyperparasitoid females, and by the primary females.
The differences observed between the copulation times of the males depending on their origin did not have any repercussions on the production of daughters, since, regardless of their origin, as many daughters were produced during the first 12 days of reproductive activity of the inseminated females ( daughters for the couples developed on secondary hosts). The production of daughters by these couples was significantly different from that of couples with higher bodyweight developed on primary hosts ( daughters).
To conclude, the expression by E. vuilleti of hyperparasitism classified as optional leads to the emergence of adult males that are smaller than those that had hatched out of primary hosts, but they have all the capabilities required to allow the parasitism of a population of primary hosts once the competitive pressure is reduced.
1 Introduction
En Afrique occidentale, une communauté hôtes–parasitoïdes cohabite dans un habitat semi-clos, de type grenier utilisé pour le stockage des graines de la légumineuse Vigna unguiculata contaminées dans les champs de culture par les Bruchidae Callosobruchus maculatus et Bruchidius atrolineatus [1]. Le stockage des graines favorise la prolifération des Bruchidae et, au bout de huit mois, la totalité du stock peut être contaminée [1]. Ces deux Bruchidae sont attaquées par trois hyménoptères parasitoïdes larvo-nymphaux, Dinarmus basalis (Pteromalidae), Eupelmus vuilleti et E. orientalis (Eupelmidae) [2,3]. La prolifération des Bruchidae assure aux parasitoïdes une grande disponibilité en hôtes potentiels, mais le chevauchement des générations, dû à la grande diversité des stades larvaires susceptibles d'être parasités, conduit à une variabilité dans la qualité de ces hôtes, variabilité augmentée par la présence d'hôtes parasités [2].
Dans un tel environnement, la compétition pour l'hôte conduit les femelles E. vuilleti à développer un comportement de cleptoparasitisme qui consiste à tuer les œufs, les larves néonates et celles du premier stade larvaire du pteromalidae concurrent D. basalis [4,5]. Ce comportement permet à E. vuilleti de détourner au profit de sa descendance la nourriture représentée par l'hôte phytophage [4–10]. Chez les femelles E. vuilleti et E. orientalis, le cleptoparasitisme évolue jusqu'à l'hyperparasitisme du dernier stade larvaire de parasites en cours de développement [9,10]. Cet hyperparasitisme, qualifié de facultatif chez les Eupelmidae, s'exerce sur le cinquième stade larvaire du Pteromalidae Dinarmus basalis, trois fois plus léger que celui de leur hôte primaire C. maculatus. Cet hôte secondaire représente une ressource alimentaire quantitativement limitée en comparaison de celle offerte par l'hôte primaire C. maculatus [6].
Le passage du parasitisme primaire vers l'hyperparasitisme facultatif, avec le cleptoparasitisme comme étape intermédiaire, s'est produit sans qu'il y ait la nécessité d'une adaptation spéciale pour pondre et se nourrir aux dépends d'un hôte secondaire ; il induit un niveau trophique supplémentaire, induisant la formation d'adultes miniaturisés du fait de la baisse pondérale de la quantité de nourriture disponible [6,7,9–11]. En effet, après un développement en hyperparasitisme, tous les paramètres pondéraux entrant dans le calcul des rendements d'ingestion et d'assimilation sont inférieurs à ceux mesurés chez les femelles et mâles Eupelmidae développés en parasitisme primaire [6,12]. Cependant, comme le rendement de conversion des éléments nutritifs assimilés est indépendant du mode de développement de la larve, la conversion en énergie utilisable par l'adulte se fait dans les mêmes rapports dans les deux voies possibles du développement larvaire primaire ou secondaire [6]. Les potentialités énergétiques des mâles et femelles E. vuilleti développés en hyperparasitisme permettent d'évaluer le coût de l'investissement de chaque parent dans la procréation d'une nouvelle génération de parasitoïdes adultes, qui accéderont à une population d'hôtes primaires. L'évaluation du coût cet investissement est possible du fait de la corrélation positive entre la quantité de nourriture ingérée par la larve parasitoïde au cours du développement et le poids de l'adulte à l'émergence [13–15].
Dans ce travail, la capacité de reproduction des mâles hyperparasites est mesurée par la quantité de spermatozoïdes stockés dans les vésicules séminales, par la qualité de l'insémination définie par le nombre de spermatozoïdes stockés dans la spermathèque de la femelle après un premier accouplement, et par les effectifs de filles que ces femelles inséminées engendreront. L'analyse des effectifs de filles, révélant la capacité des mâles à inséminer les femelles, permet d'accéder au succès reproducteur des femelles développées en hyperparasitisme par le biais de la migration des spermatozoïdes vers le réservoir de la spermathèque et par les effectifs de filles engendrées [16,17].
2 Matériels et méthodes
2.1 Insectes
Originaires du Burkina Faso, le Bruchidae C. maculatus et son cortège parasitaire E. vuilleti et D. basalis, sont élevés dans des enceintes où toutes les conditions abiotiques sont contrôlées [3,18]. Les élevages de masse des parasitoïdes ont été initiés à partir d'adultes émergés de graines de Vigna unguiculata contaminées par C. maculatus et collectées dans la région de Niamey (Niger).
Chez D. basalis, le développement larvaire passe par cinq stades, suivis d'un état prénymphal puis d'une nymphe [19]. Dans nos conditions expérimentales, le dernier stade larvaire (L5) s'observe en moyenne 10 jours après la ponte sur C. maculatus, tandis que le stade nymphal est atteint trois jours plus tard [19]. À cette étape du développement, l'hôte phytophage étant réduit à un reliquat, la larve parasite (au stade L5 ou nymphal) représente une source alimentaire de remplacement qui peut être à son tour parasitée. Tous les parasitoïdes adultes E. vuilleti utilisés dans les expériences sont issus de larves développées en hyperparasitoïdes sur des larves L5 du pteromalidae D. basalis.
2.2 Étendue de la variabilité de la taille des hyperparasitoïdes
Pour étudier la variabilité observée dans la taille des hyperparasitoïdes mâles ou femelles, cette dernière a été quantifiée par le poids frais (en milligrammes) 24 h après l'émergence des adultes, et par la largeur de la tête (prise entre les bords externes des yeux à l'aide d'un système automatique de lecture). L'intensité de la relation entre le poids et la taille de la tête (mesurée par le coefficient de corrélation) a permis par la suite de regrouper les hyperparasitoïdes mâles sur le critère « largeur de la tête » pour étudier leur production en spermatozoïdes (l'accès aux vésicules séminales nécessitant leur sacrifice).
2.3 Production des spermatozoïdes chez les mâles
La production des spermatozoïdes est analysée chez les mâles développés en hyperparasitoïdes sur les larves L5 du Pteromalidae D. basalis. L'influence du développement sur la spermatogenèse est évaluée par la quantité de spermatozoïdes produite puis stockée dans les vésicules séminales des mâles vierges de 24 h d'âge [20,21]. Pour ce faire, l'abdomen est disséqué dans une goutte d'une solution saline de Beadle (128,3 mM NaCl, 4,7 mM KCl, 2,3 mM CaCl2), et les deux vésicules séminales sont transférées dans 100 μl de la solution saline, puis éclatées à l'aide d'un potter. Par agitation douce, les spermatozoïdes sont mis en suspension. Deux gouttes de 3 μl de la solution homogénéisée sont déposées sur une lame dégraissée. Après fixation à l'alcool et coloration au DAPI (agent intercalant de l'ADN), les spermatozoïdes présents dans chaque goutte ont été comptés à l'aide d'un microscope à fluorescence [20]. Le nombre de spermatozoïdes stockés dans les deux vésicules séminales correspond à la moyenne de trois comptages, multiplié par le facteur de dilution. La production en spermatozoïdes des mâles développés sur un hôte secondaire est comparée à celle des mâles développés sur un hôte primaire.
2.4 Spermatozoïdes stockés dans la spermathèque des femelles
Les femelles vierges (âgées de 2 h) sont accouplées par des mâles de 24 h d'âge [22]. La spermathèque des femelles est prélevée 24 h après l'accouplement et la totalité de son contenu est étalé sur une lame de microscope dans 3 μl d'une solution saline de Beadle [20,21]. Les spermatozoïdes sont comptés successivement deux fois et la quantité retenue est la moyenne des deux comptages.
Deux expériences indépendantes sont menées en parallèle. Dans une première expérience, on dénombre les spermatozoïdes stockés dans la spermathèque de femelles primaires , accouplées par des mâles hyperparasites. Dans une deuxième situation expérimentale, le comptage concerne les spermatozoïdes stockés dans la spermathèque de femelles hyperparasites accouplées par des mâles primaires.
2.5 Production en filles nées des croisements impliquant des adultes développés sur un hôte secondaire
Le succès reproducteur des mâles développés sur un hôte secondaire est analysé par la capacité des femelles (développées en parasitisme primaire ou en hyperparasitisme) à produire des filles. Cette production en filles est comparée à celle de couples dont les deux partenaires se sont développés sur des hôtes primaires. Dans toutes les situations, les accouplements se font entre des femelles vierges âgées de 2 h et des mâles de 24 h d'âge [20–22].
L'activité reproductrice est mesurée pendant les 12 premiers jours de vie des femelles, qui englobe la période d'activité reproductrice maximale [3,19,20]. L'expérience débute 24 h après l'émergence des femelles, et l'activité reproductrice individuelle des femelles est suivie en décomptant les filles engendrées. Toutes les 24 h, on présente aux femelles 10 graines de Vigna unguiculata, abritant chacune deux hôtes de C. maculatus au stade L4-prénymphe de leur développement [23–25]. Ce choix du stade larvaire de l'hôte est déterminé par le fait que les femelles E. vuilleti modulent le sexe de leurs descendants en fonction de la taille de l'hôte : un maximum d'œufs fécondés (filles) étant pondus sur les hôtes de plus grande taille [23–25].
2.6 Traitement statistique
Les variables mesurées selon le type de développement subi par les parents sont analysées par l'ANOVA d'XLSTAT-Pro 6.1 pour Windows, avec un risque d'erreur de 5 %. Lorsque les variances sont égales, les valeurs moyennes ont été comparées par le test de Student (XLSTAT-Pro 6.1 pour Windows).
La régression linéaire multiple d'XLSTAT-Pro 6.1 pour Windows a été utilisée pour tester les relations la largeur de la tête et du poids frais des adultes mâles développés sur des hôtes secondaires.
3 Résultats
3.1 Taille des adultes développés sur un hôte secondaire
Les distributions du poids frais des parasitoïdes mâles et femelles permettent de différencier les échantillons de population en fonction du développement primaire ou secondaire (Figs. 1 et 2). Les mâles développés en hyperparasitisme ont des poids étalés de 0,155 à 1,10 mg, et ceux des mâles primaires se distribuent entre 0,650 et 1,500 mg (Fig. 1). La différence observée entre les poids moyens des deux catégories de mâles est significative : mâles hyperparasites ; mâles primaires (Fig. 1). De même, les poids moyens des femelles primaires ou hyperparasites sont significativement différents : femelles primaires, ; femelles hyperparasites, (Fig. 2). Pour un type de développement (primaire ou hyperparasite), les poids moyens des femelles sont significativement différents de ceux des mâles (ANOVA : développement primaire : et ; développement secondaire : et ).
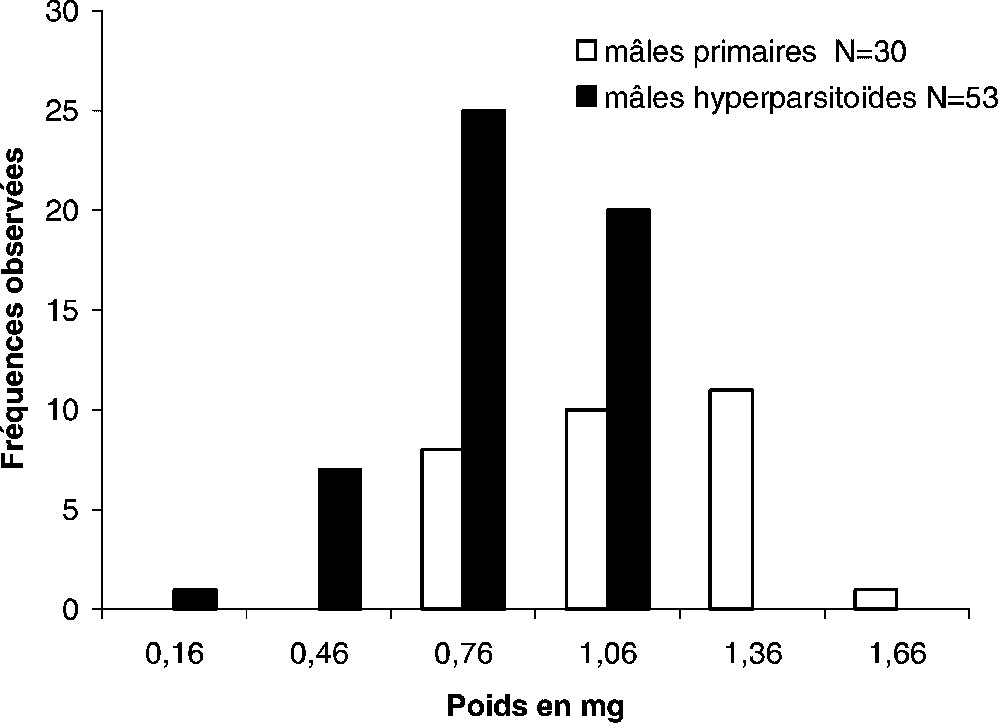
Répartition du poids frais des mâles primaires et hyperparasites de Eupelmus vuilleti. Mesurés 24 h après la naissance, les poids se répartissent de manière homogène (F calculé de Fisher : 1,142, pour α=0,05, valeur critique du F=1,788), et la différence observée entre les poids moyens est significative (moyenne ± intervalle de confiance de la moyenne : mâles hyperparasites 0,68±0,05 mg ; mâles primaires 0,965±0,10 mg ; test de Student t=5,226 au seuil de signification α=0,05.
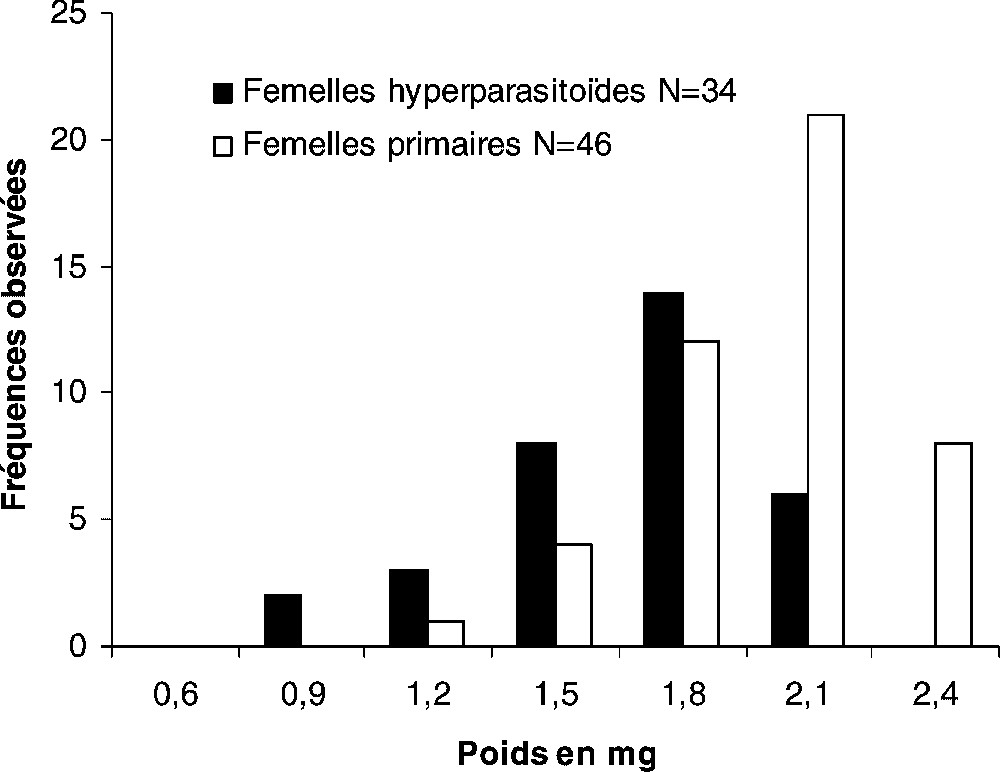
Répartition du poids frais des femelles primaires et hyperparasites de Eupelmus vuilleti. La différence observée entre les valeurs moyennes des poids est significative (ANOVA : ; P=0,0002 ; moyenne ± intervalle de confiance de la moyenne : femelles primaires, 1,90±0,30 mg, femelles hyperparasites 1,60±0,10 mg).
L'étude de la relation entre le poids et la plus grande largeur de la tête (analysée chez 24 mâles et 17 femelles hyperparasites) a mis en évidence une forte corrélation positive entre ces deux variables : chez les mâles, ; ; chez les femelles, ; . Cette corrélation positive permet de répartir les mesures de la largeur de la tête de 53 mâles hyperparasites entre trois classes de taille, définies comme petite (0,5–0,6 mm), moyenne (0,6–0,8 mm) ou grande (0,8–1 mm), les valeurs moyennes de ces trois classes étant significativement différentes (ANOVA : ; ; moyenne ± écart type de la moyenne : petite largeur : ; largeur moyenne : ; grande largeur : ).
3.2 Production de spermatozoïdes chez les mâles vierges de 24 h d'âge
Chez les mâles, primaires ou hyperparasites de 24 h d'âge, la distribution des spermatozoïdes stockés dans une vésicule séminale s'étale de 600 à 4200 spermatozoïdes (Fig. 3). La présence de deux classes modales sépare deux échantillons : la classe 1200 spermatozoïdes caractérise les mâles hyperparasites , et celle à 3000 spermatozoïdes les mâles primaires (Fig. 3). Les productions moyennes en spermatozoïdes des mâles développés sur un hôte primaire ou secondaire différent significativement : mâles hyperparasites spermatozoïdes ; mâles primaires spermatozoïdes (Fig. 3).
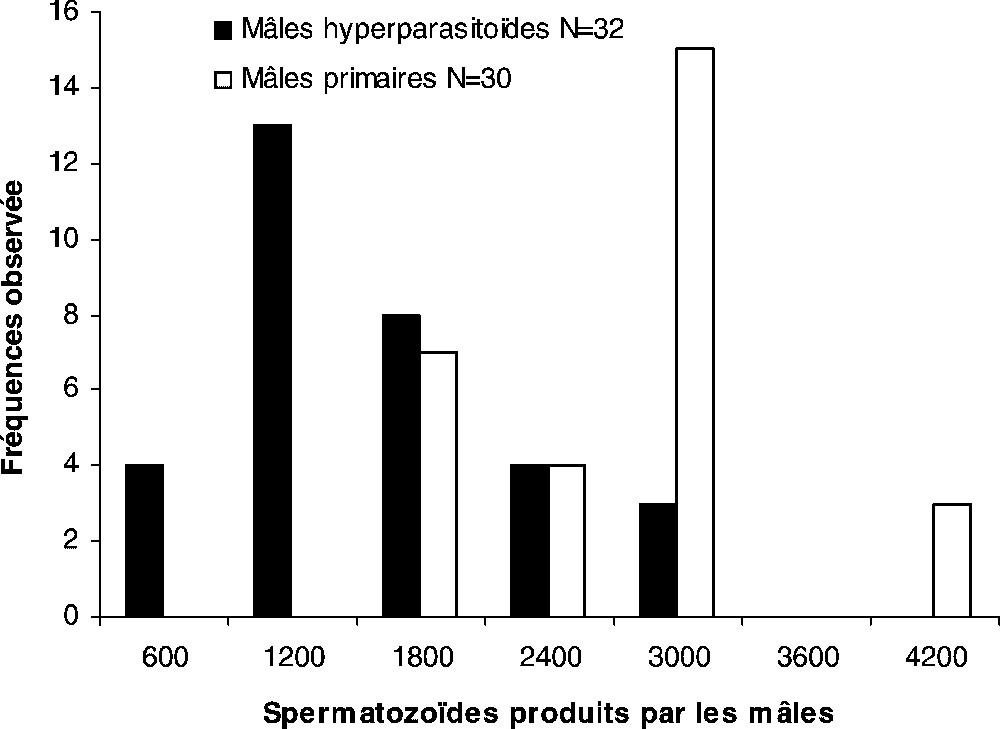
Répartition des spermatozoïdes produits par les mâles primaires et hyperparasites et stockés dans les vésicules séminales 24 h après l'émergence. Les productions étant homogènes entre les deux échantillons de populations (valeur observée du F de Fisher : 1,292 pour α=0,05, valeur critique du F=1,835), la différence observée entre les effectifs en moyenne stockés est significative (moyenne ± intervalle de confiance de la moyenne : mâles hyperparasites, 1300±240 spermatozoïdes ; mâles primaires, 2493±284 spermatozoïdes ; test de Student t=6,30 au seuil de signification α=0,05).
Un échantillon de population de 32 mâles hyperparasites, comprenant des mâles de trois tailles différentes (séparés par la largeur de la tête : petite, moyenne ou grande), a permis de comparer les productions de spermatozoïdes de ces trois classes de mâles. Ces mâles, différenciés par leur largeur de tête, ne produisent pas les mêmes quantités de spermatozoïdes (ANOVA : ; ; ), et le classement des productions moyennes en spermatozoïdes sépare les productions des mâles de petites et moyennes tailles (moyenne ± intervalle de confiance de la moyenne : respectivement, et spermatozoïdes), de celles des moyennes et grandes tailles (respectivement, et spermatozoïdes).
3.3 Stockage des spermatozoïdes dans le réservoir de la spermathèque
De 50 à 450 spermatozoïdes peuvent être stockés dans le réservoir de la spermathèque de femelles primaires ou hyperparasites 24 h après un accouplement (Fig. 4). La distribution des effectifs de spermatozoïdes dénombrés dans les réservoirs des femelles ( femelles primaires ; femelles hyperparasites) ne sépare pas les femelles en fonction du type de développement, puisque la classe modale de chaque échantillon de population se situe à 150 spermatozoïdes (Fig. 4). Les valeurs moyennes du nombre de spermatozoïdes par réservoir ne différent pas significativement : femelles hyperparasites ; femelles primaires (Fig. 4).
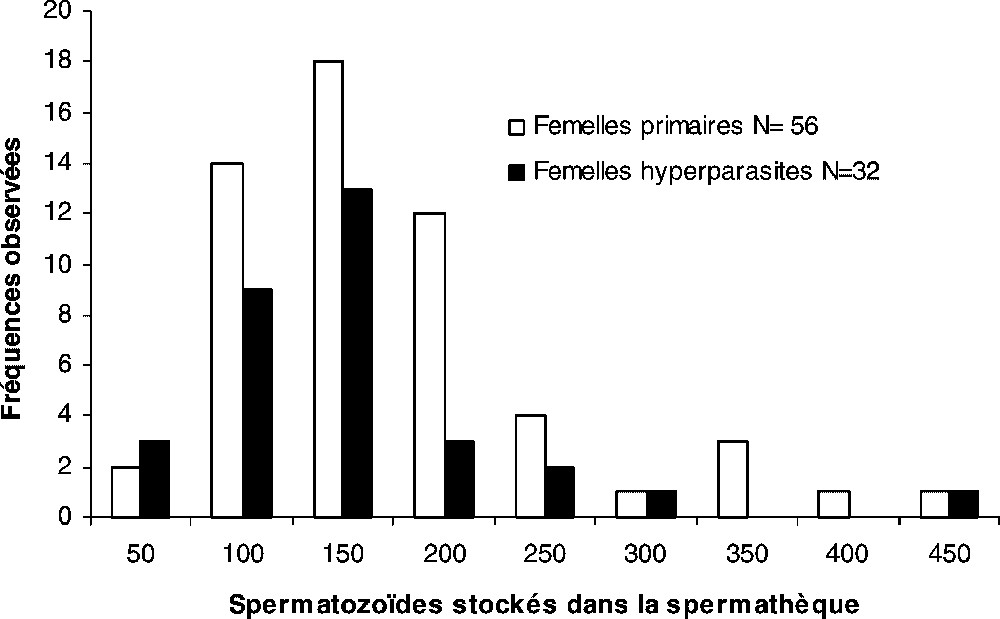
Répartition, 24 h après l'accouplement, des spermatozoïdes stockés dans le réservoir de la spermathèque des femelles primaires et hyperparasites. Les variances calculées des stocks de spermatozoïdes chez les types de femelles étant égales (valeur observée du F de Fisher : 1,119 pour α=0,05, valeur critique du F=1,737), la différence n'est pas significative entre les effectifs moyens de spermatozoïdes stockés par réservoir (moyenne ± intervalle de confiance de la moyenne : femelles hyperparasites 123±28 spermatozoïdes ; femelles primaires 150±22 spermatozoïdes ; test de Student t=6,30 au seuil de signification α=0,05).
3.4 Filles engendrées
Dans notre modèle expérimental, il y a une différence significative au niveau de la moyenne des filles engendrées entre les couples mâle hyperparasite × femelle primaire ( couples) couples hyperparasites ( couples), et couples primaires ( couples) (ANOVA : ; ; 21,20 % de la variabilité observée en filles s'expliquent par le type de croisements). Le classement des moyennes sépare significativement deux groupes : celui des parents développés en parasitisme primaire (moyenne ± intervalle de confiance de la moyenne : filles pour couples) de celui des couples dont au moins un partenaire a eu un développement sur un hôte secondaire. Quand les deux partenaires sont hyperparasites ou si seulement le père l'est, la production en filles n'est pas significativement différenciable : filles en moyenne pour couples hyperparasites, et filles chez couples où seul le père est hyperparasite.
4 Discussion
Chez les parasitoïdes où le système de reproduction est une parthénogenèse arrhénotoque, la sélection des hôtes et la distribution du sexe des descendants sont souvent corrélées à la taille de l'hôte considérée comme un indicateur de sa qualité [26,27]. En particulier, chez les parasitoïdes idiobiontes et solitaires qui ne consomment qu'un seul hôte, la sélection a favorisé l'évolution vers l'utilisation optimale des ressources alimentaires limitées à un seul hôte. Ces ressources fixées par la taille de l'hôte influent sur la taille des parasitoïdes en développement, de telle sorte que les tests qualitatifs concluent à une corrélation positive entre la taille des hôtes, la survie et le succès reproducteur des adultes émergeants [25–31]. L'histoire des traits de vie des femelles E. vuilleti et E. orientalis développées sur leur hôte primaire C. maculatus a été analysée par l'étendue de la période de reproduction, de la production journalière en œufs et des descendants [3,23,25]. Quant au succès reproducteur des mâles primaires E. orientalis, il a été évalué par leur disposition à inséminer avec succès les femelles et la capacité de ces dernières à engendrer des filles [20].
Ces liens entre taille des adultes et qualité des hôtes existent aussi chez les espèces capables de parasiter des parasitoïdes en développement en cas de forte compétition interspécifique pour l'appropriation des hôtes primaires, ce qui leur permet d'accéder à des ressources alimentaires complémentaires [5,11,12]. Dans un habitat de l'hôte relativement clos (grenier de stockage des graines de Phaseolinae), quand le niveau trophique de l'hôte primaire (stades larvaires des Bruchidae) est saturé par les parasitoïdes primaires (E. vuilleti, E. orientalis, D. basalis), les E. vuilleti femelles expriment une plasticité comportementale très agressive, consistant à hyperparasiter le dernier stade larvaire du Pteromalidae D. basalis [4,5,9].
Ce choix comportemental d'hyperparasiter représente un compromis évolutif vers une reproduction future sur des hôtes primaires. En effet, après un développement sur un hôte secondaire, les adultes E. vuilleti ont une taille miniaturisée, quantifiée par un poids frais à l'émergence significativement plus bas que celui des mâles et femelles développés sur l'hôte primaire C. maculatus. Ces adultes miniatures représentent un échantillon de population de parasitoïdes dont le succès reproducteur peut être évalué par l'issue de la compétition pour l'accouplement entre mâles de tailles différentes, par le nombre de femelles qu'ils sont aptes à inséminer au long de leur vie (lié à la longévité des mâles), par les effectifs de filles engendrées. Selon les espèces, l'étude de la capacité des mâles à s'accoupler selon la taille révèle une hétérogénéité de réponses. Par exemple, au cours de la parade pré-copulatoire, les petits mâles qui produiraient moins de stimulations auraient moins de succès dans le maintien d'une position dominatrice [32]. Au contraire, ceux de Pimpla nipponica (Ichneumonidae), en présence d'une gamme de tailles de femelles, seraient avantagés à l'accouplement, dans le cas d'une compétition mâle–mâle, tandis que chez l'Encyrtidae Anagyrus kamali, les petits ou grands mâles auraient autant de succès avec des femelles de taille moyenne [33,34]. Chez l'Eupelmidae E. vuilleti, les mâles miniatures ont gardé la capacité à s'accoupler, d'autant plus que le grégarisme des hôtes favorise les accouplements sur le lieu d'émergence [6,35]. Bien que trois fois plus légers que les femelles primaires, les mâles miniatures les inséminent, et la copulation dure en moyenne , alors qu'elle atteint chez les couples primaires (observations personnelles d'Éric Darrouzet : ce laps de temps ne comprend pas les séquences pré- et post-copulatoires). Cependant, la taille ne peut pas être considérée comme le facteur-clé du pouvoir reproducteur des mâles hyperparasites, car les petits mâles de Lariophagus distinguendus engendrent autant de filles que ceux de plus grande taille [36]. Si la valeur adaptative des mâles désigne selon la définition classique « the expression of an individual's success of passing on its genes to future generations » (R. Dawkins [37]), c'est l'investissement des mâles dans les futures générations mesuré par les effectifs de filles engendrées qu'il faut connaître. Or, cet investissement des mâles dépend de leur capacité à produire des spermatozoïdes, puis de la quantité de spermatozoïdes transmise aux femelles à la copulation et en dernier lieu de la quantité de spermatozoïdes stockée dans la spermathèque des femelles. Par exemple, un mâle E. orientalis primaire, vierge et de 24 h d'âge, a en réserve approximativement 5500 spermatozoïdes en moyenne dans une vésicule séminale, en transfère au cours d'un accouplement 1650 en moyenne, mais seulement 21 % de la quantité transférée est stockée dans la spermathèque d'une femelle de 24 h d'âge, qui n'en utilisera que les trois cinquièmes tout au long de sa vie [20].
Chez les mâles hyperparasites, la production en spermatozoïdes est en moyenne de spermatozoïdes, alors qu'elle atteint spermatozoïdes chez les primaires. Dans nos conditions expérimentales, 72,30 % de la variabilité de cette production est liée à la qualité du développement des mâles, du fait des paramètres pondéraux plus élevés chez les parasitoïdes primaires. Ces mâles d'Eupelmidae, catalogués comme « mâles synspermatogéniques », produisent des spermatozoïdes tout au long de leur vie, et l'énergie nécessaire à cette production (liée aux protéines et aux amino acides) provient de la prise de nourriture de nectar ou/et pollen, mais aussi, de façon opportuniste, de l'hémolymphe de l'hôte, qui sourd par les blessures occasionnées par les piqûres des femelles au cours de la ponte [38,39]. Il est probable que ces ressources alimentaires à la disposition des adultes permettent le maintien ou/et l'augmentation des potentialités énergétiques des mâles miniaturisés et qu'elles participent à augmenter leurs chances de survie et leur pouvoir reproducteur. Toutefois, ces hypothèses restent à démontrer chez les mâles, alors que chez les femelles primaires les nutriments acquis au cours du nourrissage sur l'hôte sont stockés et utilisés graduellement pour la production d'œufs tout au long de leur vie [40].
La capacité des couples à engendrer des filles, évaluée par le stock de spermatozoïdes dans le réservoir de la spermathèque, ne varie pas de manière significative selon le développement larvaire de chaque partenaire (stock moyen des femelles primaires : spermatozoïdes ; stock moyen des femelles hyperparasites spermatozoïdes). Après un premier accouplement, il est remarquable de constater que le potentiel en spermatozoïdes mobilisables par les femelles pour engendrer des filles est équivalent, malgré la différence d'origine des femelles (primaire ou hyperparasite), et les quantités différentes de spermatozoïdes stockés dans les vésicules séminales des mâles primaires ou hyperparasites. Le stock en spermatozoïdes des femelles pourrait résulter des mécanismes régulateurs des comportements post-copulatoire et/ou de la migration des spermatozoïdes vers le réservoir après l'insémination.
Bien qu'ayant un potentiel équivalent de spermatozoïdes en réserve, les femelles primaires ou hyperparasites n'engendrent pas autant de filles. Sur les 12 premiers jours d'activité reproductrice, les couples hyperparasites engendrent en moyenne filles, alors qu'un couple de parasites primaires a en moyenne filles. Dans les deux voies du développement larvaire, la corrélation positive entre le poids du corps de l'adulte et la nourriture ingérée par les larves fait qu'il est possible de comparer le coût de l'investissement de chaque parent dans la reproduction [6,13,14]. Les réserves énergétiques des femelles primaires E. vuilleti acquises au cours du développement larvaire leur permettent de pondre en moyenne œufs par jour, alors que les femelles hyperparasites n'en pondent que ([41], observations personnelles D. Rousse). Ces écarts dans la production en filles peuvent ne pas être totalement dus au seul poids corporel des adultes impliqués dans les croisements, car des femelles primaires inséminées par des mâles hyperparasites n'engendrent que filles. En effet, si tel était le cas, les réserves énergétiques des femelles primaires, accumulées au cours du développement primaire pour couvrir leurs besoins métaboliques, devraient leur permettre d'engendrer autant de filles et ce, quelle que soit l'origine de leur partenaire. Or, comme cette hypothèse n'est pas vérifiée par l'expérimentation, les différences de production de filles doivent être expliquées autrement, et l'hypothèse émise et à tester est que ces différences sont fonction « d'un facteur mâle », par le biais de la quantité ou/et la qualité des spermatozoïdes et des sécrétions mâles qui leur sont associées.
L'apparition dans un habitat semi-clos de mâles et femelles miniaturisés de E. vuilleti reflète la grande plasticité comportementale de cette espèce. En pratiquant l'hyperparasitisme du Pteromalidae concurrent, les nouveaux adultes Eupelmidae miniaturisés conservent toutes leurs potentialités reproductives pour réinvestir une population d'hôtes primaires quand la pression compétitive s'abaissera. Cette pression compétitive due à la vie communautaire dans un habitat relativement clos (type greniers pour graines) favorise l'expression d'une coévolution entraînant l'espèce E. vuilleti vers une adaptation comportementale préservant sa reproduction future, alors que l'espèce D. basalis a adopté une stratégie d'évasion [4,42].