

1 Introduction
Drugs of the anthracycline group, in particular doxorubicin (Adriamycin; DXR), are among the most effective and extensively used anti-cancer agents [1]. Severe cardiotoxicity, both acute and chronic, presents, however, an important complication of this therapy [2–4]. Acute effects can occur immediately after treatment and are characterized by transient arrhythmias, reversible hypotension and pericarditis [3,5]. Far more serious is chronic cardiotoxicity that can manifest itself years to decades after treatment. It is characterized by progressive left ventricular dysfunction leading to irreversible congestive heart failure. Mortality rate associated with DXR-induced congestive heart failure is estimated to be at least 20% [6].
Different hypotheses, accentuating various mechanisms and/or targets, have been proposed to explain DXR cardiotoxicity (see [2–4]), but no single one of these is considered fully satisfying. Rather, DXR cardiotoxicity seems to be a multifactorial process that leads to cardiomyocyte apoptosis as the terminal downstream event [2].
Impairment of cardiac high-energy phosphate metabolism has been recognized as an important feature of both acute and chronic anthracycline cardiotoxic action. Recent progress in understanding cardiac bioenergetics and new experimental techniques support the role of cellular energy deficits in the development of cardiac myophathies of different origins [7–9], and this seems to be valid also for cardiomyopathy and congestive heart failure induced by DXR. Energetic deficits caused by DXR have been for quite some time associated with compromised mitochondrial function [10]. Recently, some evidence has suggested that regulatory and compensatory components of myocardial energy metabolism, such as the phosphotransfer network of creatine kinase (CK) or the signalling pathway of AMP-activated protein kinase (AMPK) are among the highly sensitive and early targets of anthracycline-induced damage (see Fig. 1). Here, we will review the defects in the energy-providing system of the myocardium that are caused by DXR and discuss their functional consequences. We will first summarize the experimental evidence for DXR-induced changes in high-energy phosphate levels (ATP and phosphocreatine, PCr), as well as in mitochondrial ATP production. We will then briefly mention accompanying changes in the profile of cardiac substrate utilization. In a last section, we will emphasize DXR-induced defects in the CK energy storage and transport system, as well as the energy sensing and signalling AMPK pathway. Although we will mainly focus on data concerning the effects of DXR in heart, most of the considerations will be valid also for other anthracyclines. For further information on DXR, including general aspects, please refer to numerous excellent reviews covering DXR chemical structure and related oxidative chemistry [1,2,4], interaction with iron [11], intracellular generation of DXR-induced free radicals [6], generation and action of DXR secondary metabolites [2,3,12] distinction in therapeutic and toxic action of DXR [2], or clinical and histological characteristics of DXR-induced heart failure [3–5].
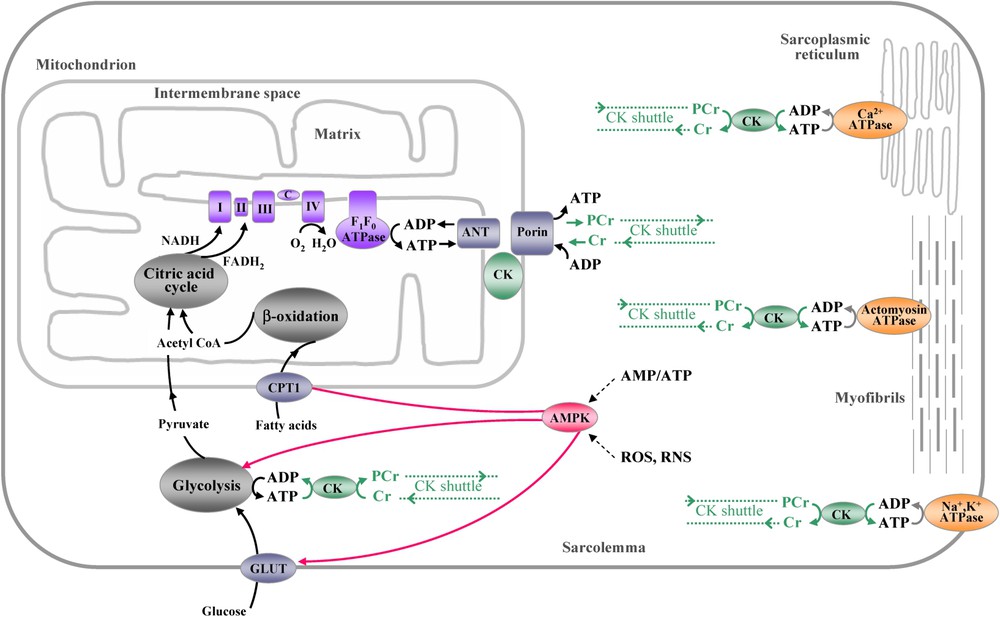
Schematic overview of ATP production, consumption and transfer of high-energy phosphates in cardiac muscle. Mitochondria represent the main site of myocardial ATP production. Glycolysis in the cytoplasm and fatty acid oxidation in the mitochondrial matrix drive the citric acid cycle producing NADH and FADH2 that supply electrons to the respiratory chain in the inner mitochondrial membrane. The generated electrochemical proton gradient across the inner mitochondrial membrane is used by ATP synthase to drive ATP production. Adenine nucleotide translocator (ANT), associated with sarcomeric mitochondrial creatine kinase (CK) and porin (VDAC), ensures channelling of substrates and products of oxidative phosphorylation between mitochondrial matrix and cytosol. Newly synthesized ATP channelled by ANT to the intermembrane space is used by CK to phosphorylate Cr. The thus formed PCr is exported to the cytosol by porin, while ADP is channelled back to the matrix. Similarly ATP produced during glycolysis is converted to PCr by cytosolic CK isoforms that are coupled to glycolytic enzymes. Rapidly diffusing PCr connects sites of energy production with sites of energy consumption (CK energy shuttle). The main myocardial ATP consuming sites are actomyosin ATPase in myofibrils, the Ca2+-ATPase of sarcoplasmic reticulum and the Na+, K+-ATPase in the sarcolemma. At energy consuming sites, CK isoforms generate ATP from PCr to maintain constant ATP/ADP ratios even during elevated workloads (CK energy buffer). AMPK, activated by a fall in cellular energy state, in particular by an increase in the AMP/ATP ratio, as well as oxidative stress, plays a role as the key energy sensor, signalling system and regulator of energy substrate utilization, mainly fatty acids and glucose. All the depicted enzymes, transporters and metabolic pathways seem to be affected to a different extent by doxorubicin; accumulation of such defects may finally lead to irreversible cardiac failure (for details, see text).
2 Myocardial high-energy phosphates
Heart requires large amounts of energy to sustain its contractile performance. DXR has been reported to reduce cardiac energy reserves, in particular ATP and PCr levels. This effect has been consistently observed in different animal models for cardiotoxicity, as well as in patients. A large number of studies have shown that DXR reduces intracellular ATP and/or PCr concentrations in cultured cardiomyocytes [13–22]. For example, as shown by Jeyaseelan et al. [17], 1 μM DXR decreased ATP of cultured cardiomyocytes, calculated per milligram of protein, by more than 50% during 24 h. Vidal and colleagues [22] reported markedly decreased nucleotide levels in cytosolic and mitochondrial fractions of cultured cardiomyocytes already several minutes after incubation with 800 μM DXR. With this very high DXR dose, cytosolic ATP dropped to about 10% of the initial level after 60 min. In a model of isolated rat heart, Pelikan et al. [23] found that a lower effective DXR dose, corresponding to a 70-min perfusion with 10 μM DXR, induced a 20% decline in both ATP and PCr levels and similar data have been reported by Ohhara et al. [24]. Cini Neri et al. [25] showed that in hearts isolated from rats treated for 3 weeks with DXR (3 mg/kg per week), ATP and GDP concentrations were reduced by about 50%. Zhou et al. [26] found an about 30% decrease in the ATP content of myocytes isolated from hearts of rats, which received 6 weekly injections of DXR (2 mg/kg per week). Using in vivo 31P-NMR spectroscopy in rats, Nicolay and colleagues [27] evaluated acute and longer-term (up to one week) effects of DXR on cardiac ATP and PCr levels. In this study, a dose-dependent decline in cardiac PCr content occurred acutely within 30–60 min after injection of DXR and reached about 20% at 25 mg/kg DXR, the maximal dose used. The same authors observed diminished PCr levels and PCr/ATP ratios several days after a single higher dose or after multiple lower doses. It is important to note that the longer-term effects of accumulated multiple lower dosages of DXR were more pronounced than acute effects of an equivalent single dose. Finally, a study with 31P-NMR demonstrated a similar decrease in the PCr/ATP ratio in children as late as four years after completion of the treatment [28]. In some cases, the PCr/ATP ratio has been diminished even in the absence of clinical manifestations of cardiomyopathy.
For a critical evaluation of the demonstrated effects of DXR, it should be kept in mind that the choice of a correct, clinically relevant DXR concentration depends on the model system. For isolated mitochondria, such relevant concentrations were evaluated to be in the range 5–30 μM [12], while for isolated cells they may be less than 1 μM [29]. For perfused heart, a good indication can be the peak plasma concentrations of the drug in patients, which reach up to 2–6 μM after bolus injection, with a typical value of 1–2 μM [29]. In models with a short life time and thus short time of drug persistence like perfused heart or isolated mitochondria, very often supraclinical DXR concentrations have been applied, up to several hundred μM. However, such an approach, which is intended to simulate the effects of lower DXR doses over longer time, must always be evaluated with care.
3 Mitochondrial function
3.1 Respiratory chain
More than 90% of the ATP utilized by cardiomyocytes is produced by mitochondrial respiration [9]. Therefore, any alteration of mitochondrial structure and function are fundamental for cardiomyocyte function. Progression of DXR cardiotoxicity is associated already at an early stage with morphological abnormalities of mitochondria, including mitochondrial swelling that is typical for apoptotic cells. Such ultrastructural changes have been observed in different cardiotoxicity models, as well as in patients [3,30,31]. In parallel, a decrease in mitochondrial oxidative capacity has been reported upon DXR treatment. Incubation of isolated heart mitochondria with DXR rapidly affected mitochondrial respiration, both through complex I and II substrates [32–35]. It has been shown that respiratory state 3 (oxygen uptake in presence of ADP) is always inhibited by DXR, while state 4 (oxygen uptake in the absence of ADP) is often stimulated [34], possibly due to uncoupling [35]. Instantaneous effects on isolated mitochondria require, however, rather high DXR concentrations of several hundred μM ([34,35] see also [36]), with the exception of succinate oxidation under phosphorylating conditions in presence of hexokinase, which is inhibited already at an IC50 of about 5 μM DXR [35]. Here, hexokinase mimics an ATP utilizing and ADP regenerating intracellular system, which can interact with mitochondrial outer membrane porin for local nucleotide exchange [37]. It remains to be established to which extend impaired mitochondrial respiration contributes to decreased cardiac function in situ and how this process evolves over years in chronic DXR toxicity. In cardiac preparations, as, e.g., perfused heart, it seems that DXR can acutely affect cardiac performance without marked change in mitochondrial function. In mitochondria isolated from rat hearts that were previously perfused for 1 h with 10 μM DXR, which is a slightly supraclinical concentration, mitochondrial respiration and oxidative phosphorylation remained unchanged in spite of an important decline in heart function [38]. It is to note that this acute setting was sporadically linked to an increase in respiratory state 4. Similarly, increased state 4 was observed in mitochondria isolated from rats 24 h after injection of DXR at 8 mg/kg [39]. On the other hand, several authors reported inhibition of respiratory state 3 in heart mitochondria isolated from treated animals. Yen et al. [40] demonstrated that mitochondria from mice treated with 20 mg/kg of DXR for 5 days display a 20–30% decrease in state-3 respiration rate, both with pyruvate/malate and succinate as substrate, while there was no change in state 4. Positron emission tomography (PET) was used to study DXR effects on heart oxygen metabolism of patients [41]. In this study, no significant effect of DXR on blood flow or metabolic flux through the citric acid cycle was observed, neither acutely, 24 h after administration of 50 mg/m2 DXR, nor after 3 weeks after cumulative administration of 300 mg/m2 DXR, despite a small but significant decrease in left ventricular ejection fraction. The authors interpret the lack of DXR effects on blood flow and citric acid cycle flux as indicative for the absence of changes in the oxidative phosphorylation, although this conclusion is fairly indirect.
Besides the analysis of global DXR effects on mitochondrial respiration, several studies have tried to identify the specific respiratory complexes affected by DXR, using mitochondria, submitochondrial preparations, or isolated complexes. Nicolay and de Kruijff [42] analyzed immediate DXR effects on different respiratory chain activities in mitochondria and inner membrane ghosts isolated from rat and bovine hearts in presence of uncoupler CCCP to avoid indirect effects of the drug, caused, e.g., by drug-induced uncoupling. According to this study, DXR-sensitive sites were mainly located in complex III and IV, and to a minor extent in complex I, but were almost absent in complex II. The IC50 of DXR was 150–200 μM for enzymes of complex III and IV, such as coenzyme Q-1, duroquinone, or cytochrome c oxidase, and 425 μM DXR for NADH-duroquinone oxidoreductase in complex I, while ferricyanide and coenzyme Q-1 reduction in complex I were essentially unaffected up to 1 mM DXR. For the combined activities of complex I–III (NADH-cytochrome c oxidoreductase activity), II–III (succinate–cytochrome c oxidoreductase activity), I–III–IV (NADH oxidase activity), or II–III–IV (succinate oxidase activity), IC50-values in the range of 60–200 μM DXR were observed. However, in another study, inhibition of complex I–III at a much lower IC50 of about 3 μM has been demonstrated in bovine heart mitochondria exposed to DXR [43,44]. Using bovine heart submitochondrial preparations that were incubated with 25–30 μM DXR for 15 min, Marcillat and colleagues [45] showed a 70% decrease in activity of NADH dehydrogenase, while other respiratory enzymes such as succinate dehydrogenase, succinate oxidase, and cytochrome c oxidase required DXR concentrations of 500 μM or more for a 50% inhibition. From these studies, it seems that multiple steps in electron transport through complexes I, III and IV are a target of DXR toxicity, with a possibly higher susceptibility of NADH dehydrogenase and cytochrome c oxidase. It is important to mention that DXR activated by horseradish peroxidase and hydrogen peroxide seems to be a more potent inhibitor of enzymatic activities. Studies of Muraoka and Miura [46], using pig heart submitochondrial particles, have shown that mitochondrial succinate dehydrogenase is inactivated by 50% within 30 min already by about 0.8 μM DXR when the drug is activated by the horseradish peroxidase/hydrogen peroxide system.
The effect of DXR on respiratory complexes has been also studied with animal models. Mitochondria isolated from rats receiving total doses of 15 mg/kg DXR over 2 weeks showed decreased activity of complex I, as assessed by NADH oxidation. An effect was seen 3 weeks after the treatment, well before left ventricular ejection fraction and fatty acid metabolism became significantly impaired [47]. In mitochondria from mice treated with 20 mg/kg of DXR for 5 days, complexes I and II were found inhibited [40]. Finally, 48 h after a single DXR injection of 40 mg/kg into mice, activity of respiratory complexes IV and I–III declined to about 50% of the initial value, while complex II activity remained unchanged; similar values were reported for animals several weeks after a chronic DXR administration [30].
Discrepancies between the reports cited above can have numerous reasons in addition to the obvious differences in the experimental systems and parameters analyzed (e.g., overall mitochondrial function or activity of specific elements of the electron-transport chain). Apparent activities of respiratory enzymes depend on the composition of assay media, e.g., the presence of detergent, which can affect accessibility of sites of interest [42]. In experiments with intact organelles, the functional state of mitochondria is of prime importance and a treatment of isolated mitochondria with DXR appears to be particularly delicate. Isolated mitochondria are per se very fragile organelles and exhibit regulatory properties, which are to some extent different from mitochondria maintained within the cell. In this experimental system, also the definition of toxicity conditions in terms of time and dose, as well as the interpretation of results in regard to other experimental or clinical settings require particular care. It should always be kept in mind that mitochondria selectively accumulate DXR to concentrations much higher than plasma levels (see [12]). Moreover, similarly as in cultured cardiomyocytes, any comparison of data has to consider the effective DXR dose, i.e. drug concentration per mg protein in the experimental system, which is not always clear in the literature.
3.2 Oxidative phosphorylation
Besides defects in the electron transport chain, DXR also impairs the coupled synthesis of ATP. Although effects of DXR on mitochondrial ATPase activity has been reported for heart mitochondria [33,35], there is evidence that other mechanisms are more important. A major one may be the uncoupling activity of DXR, which is indicated by the frequently observed stimulation of respiratory state 4 [34,35,38,39].
Other targets of DXR that potentially impair oxidative ATP production are proteolipid complexes involved in channelling of ATP and phosphocreatine from mitochondria into the cytosol. These multienzyme transmembrane complexes, together with the mitochondrial respiratory chain and the ATP synthase are necessary for efficient mitochondrial ATP synthesis and export. They contain adenine nucleotide translocator (ANT) of the inner mitochondrial membrane and porin (voltage dependent anion channel, VDAC) of the outer membrane. Another frequent component is mitochondrial creatine kinase (MtCK), which is situated in the intermembrane space and functionally and structurally associated with ANT and VDAC [48–51]. As suggested by several lines of evidence, DXR compromises the functioning of these ‘channelling complexes’. DXR affects ANT1, the ANT isoform mainly expressed in heart [17,52] and the inorganic phosphate carrier, an ANT homologue [53,54], and inhibits the function of MtCK. The interference of DXR with MtCK is described in more detail below.
3.3 Involved mechanisms
All available evidence points to pleiotropic effects of DXR in mitochondria and thus to a multitude of molecular mechanisms involved in DXR-induced mitochondrial damage. High-affinity binding of DXR to cardiolipin, an anionic phospholipid specific for the inner mitochondrial membrane, has been suggested as a possible common cause of inactivation of several enzymes involved in respiration and oxidative phosphorylation [55,56]. Besides nuclei, mitochondrial membranes have been identified as the most important sites of doxorubicin accumulation in the cell, where local concentration of the drug becomes much higher than in plasma [12,29,57]. Insertion of DXR into membranes would modify the lipid environment, which in turn could modify the kinetic properties of membrane associated enzymes, as, e.g., cytochrome c oxidase [44,58,59]. Bound DXR would also induce dissociation of cardiolipin associated proteins from the inner mitochondrial membrane, e.g., cytochrome c and MtCK. However, as it was shown for MtCK, DXR can also directly modify the enzyme structure through oxidative damage [60]. Accumulation of DXR in mitochondria would obviously favour multifactorial molecular damage, which can be mediated by free radicals, iron or doxorubicin metabolites [3,4,11].
DXR effects in mitochondria can be also mediated and/or amplified by mitochondrial proteins. Mitochondrial oxidoreductive enzymes, mainly NADH dehydrogenase, have been identified, together with cytochrome P-450 reductase, and xanthine oxidase, as the principal mediators of DXR reduction in the cell [3,61–64]. One-electron reduction initiates a very dangerous process of redox cycling of the drug, deviating electrons from the respiratory chain and leading to the generation of reactive oxygen and nitrogen species and resulting in DXR-associated oxidative stress.
Moreover, the response of mitochondrial respiration in situ can be modulated by other parameters related and sensitive to DXR, such as oxygen and/or substrate availability, levels of ATP, ADP, or PCr, the degree of mitochondrial coupling, peroxidation of mitochondrial membranes, or calcium homeostasis. The latter seems especially important, because in myocytes calcium and respiratory control are intrinsically linked. DXR-induced alterations in calcium handling and their possible consequences are beyond the scope of this study, but are described in several original papers and reviews [3,11,20,26,52,65–67]. It should be added that besides inducing functional changes, DXR was shown to downregulate expression of several mitochondrial enzymes involved in respiration and oxidative phosphorylation, as, e.g., cytochrome c oxidase [31,68], Reiske iron-sulphur protein, and ANT1 [17,52]. In fact, a coordinated downregulation of mitochondrial enzymes controlling ATP synthesis is typical for different heart pathologies [8].
4 Myocardial substrate utilization
4.1 Fatty acid oxidation
The heart metabolizes multiple substrates to synthesize ATP. Fatty acids are known to be a major cardiac energy source under aerobic conditions. Using cell culture and animal cardiotoxicity models, it has been shown that DXR affects fatty acid metabolism in the heart [69–71]. Animals show increased serum free fatty acid level following DXR treatment [72,73], and perturbed fatty acid metabolism has also been found in DXR-treated patients [74]. DXR inhibits the oxidation of palmitate, a long-chain fatty acid, either acutely within minutes as in isolated cardiomyocytes treated with up to 2 mM DXR, or in a chronic situation as in cardiomyocytes isolated from DXR-treated rats (18 mg/kg divided in 6 doses over 2 weeks). As underlying mechanism, impairment of carnitine palmitoyl transferase I (CPTI) and/or depletion of its substrate l-carnitine was proposed [69]. External l-carnitine and probucol, a drug with hypolipidemic and antioxidant action, have been shown to diminish cardiotoxic side effects in treated animals [75–81]. Such DXR-induced downregulation of fatty acid oxidation represents a further feature of disturbed mitochondrial function that is characteristic for heart pathology.
4.2 Glucose transport and glycolysis
During the development of cardiac pathologies, at least in their early stages, decreased fatty acid oxidation is accompanied by upregulation of glucose utilization as a compensatory response. The situation following DXR treatment seems different. As shown in rat models, DXR-induced cardiomyopathy is associated with a decreased utilization of both substrates, fatty acids and glucose [70]. Similar results were obtained with cultured cardiomyocytes, although here higher DXR concentrations were requited for inhibition of glucose oxidation as compared to fatty acid oxidation [69]. Diminished glycolytic rate may be due to DXR effects on glucose supply and/or the ability of cells to stimulate it. In cardiomyocytes, treatment with 1 μM DXR led to a transient increase in glucose uptake of about 50% after 1 h, followed by marked decrease [82]. Another reason for decreased glycolysis can be the impairment of phosphofructokinase (PFK), the rate-limiting enzyme of glycolysis. As shown by Jeyaseelan and colleagues for cardiomyocytes [17], exposure to 1 μM DXR for 2–3 h decreases the mRNA level for PFK by about 50%. These findings demonstrate the limitations of DXR-treated heart to compensate a low cellular energy state with increased glycolytic ATP generation.
5 High-energy phosphate storage and transfer
DXR not only diminishes general energy production, but already very early leads to a more specific impairment of creatine kinase isoenzymes. In heart, cytosolic and mitochondrial isoenzymes of creatine kinase (CK), together with easily diffusible creatine (Cr) and phosphocreatine (PCr), provide an important cellular energy buffer and energy transport system, bridging sites of energy production and consumption (see Fig. 1; for reviews see [83,84]). Octameric mitochondrial creatine kinase (MtCK) is functionally coupled to oxidative phosphorylation, and part of dimeric cytosolic CK isoforms (MM-, MB-, BBCK) to glycolysis and to various ATP consuming pathways, including the main cardiac ATP utilizing sites such as the actomyosin ATPase in myofibrils, the Ca2+-ATPase of sarcoplasmic reticulum and the Na+, K+-ATPase in the sarcolemma.
Interference of DXR with CK has been found at different levels of complexity, from the purified enzyme in solution to animals and some involved molecular mechanisms have been elucidated.
Our group has shown that DXR impairs structure and function of purified CK isoforms in vitro, including MtCK [60] and cytosolic CK isoforms (M. Tokarska-Schlattner, unpublished data). DXR-induced damage not only leads to inactivation, which was observed with all CK isoforms, but also to further, specific injury of the MtCK isoform, namely dissociation octamers into dimers and inhibition of MtCK binding to mitochondrial membranes, in particular to cardiolipin [60,85,86]. Most importantly, the cardiac isoform sMtCK occurred to be more sensitive to DXR than the ubiquitous uMtCK, which could be explained by the slightly different molecular properties and structures of these isoforms [87–89]. Such differential sensitivity of s- and uMtCK to DXR may contribute to the selective toxicity of the drug to heart tissue. As shown for purified enzyme, the DXR effect on MtCK membrane binding is immediate, probably due to a competitive binding of the drug to the main MtCK receptor, cardiolipin. In contrast, enzymatic inactivation and dissociation of MtCK octamers into dimers by 5–750 DXR occur in solution only after hours to days. Inactivation is due to direct oxidative modification of the active site cysteine at DXR concentrations below 100 μM, while generalized damage of different residues, partially mediated by superoxide anions, becomes significant at higher DXR doses. It must be stressed that for in vitro experiments with purified MtCK, the clinically relevant concentrations should be higher than for isolated mitochondria, i.e. of the order of a few tens of μM, since MtCK and DXR share cardiolipin as a common binding partner in the inner mitochondrial membrane, leading to high local concentrations of DXR in the microenvironment of MtCK. In addition, activation of the drug by peroxidase/H2O2 or its complexation with iron have been shown to accelerate inactivation of purified CK or CK in heart homogenate [90,91]. In such systems, 10–30 μM DXR induce CK inactivation within a few tens of minutes. This mechanism, as well as the more intensive production of reactive oxygen and nitrogen species in situ [4,92,93] can enhance inactivation of CK that is very sensitive to oxidation and nitration, in particular in case of MtCK situated at the very origin of radical generation.
In situ, acute effects of DXR on the CK system have been studied in the perfused heart model [94]. Perfusion was performed during 1 h at two DXR concentrations, 2 and 20 μM, corresponding to the peak plasma concentration in patients [29] and to the lowest concentration that produces substantial functional changes in the perfused heart, respectively [23,95]. In this acute situation, dimerization of MtCK and inhibition of cardiolipin binding were detectable already at 2 μM DXR, and became significant at 20 μM. At the latter concentration, also a mild decrease in total CK activity by about 20% was observed. In spite of these defects, which occurred relatively early, the CK system probably maintained its functionality under such acute conditions because of a compensatory shift in the expression between cytosolic CK isoforms, characterized by decreased MCK and increased BCK protein. This isoform shift was already apparent after 1 hour of perfusion of the heart at the low dose of 2 μM DXR. Re-expression of such a foetal CK isoenzyme pattern, that is characterized by BCK accumulation, is well known for chronic heart pathologies as cardiac hypertrophy or failure [9,96,97]. At short exposition times and/or lower DXR doses, when inhibition of specific CK activity is not yet very pronounced, the isoform shift can indeed have a compensatory function. Increased BCK may present a functional advantage related to its higher affinity for ADP/PCr as compared to MCK, possibly allowing more efficient ATP regeneration [96]. The shift in the CK isoenzyme pattern could be due to rapid changes in gene expression, either because of a general induction of a foetal expression profile in response to stress [96,98] or because of specific susceptibility of CK genes to DXR [99]. The latter is supported by treatment of cultured cardiac muscle cells for 24 h with 0.5 μM DXR, which led to a marked reduction of MCK mRNA [99]. In addition, MCK may be degraded upon DXR treatment or may partially leak from DXR-treated cardiac cells [6,100]. It should be noted that elevated plasma CK levels, resulting from cellular leakage, are a clinical marker for different cardiac pathologies, including DXR toxicity [6].
With prolonged DXR exposure, substantial direct and radical mediated molecular damage of CK accumulates gradually. In cardiomyocytes cultured with 50 μM DXR, a pronounced impairment of the CK system was observed after 24 h of treatment [100]. Interestingly, total CK activity decreased significantly in detached cells by about 67%, while it increased in attached cells by about 40% during 48 h of treatment. Increased specific CK activity of attached cells, as observed in cardiomyocyte experiments, may also be indicative for a compensatory mechanism within the CK system. The same report described a significant 50% decrease in total CK activity in hearts of DXR treated rats after 3 doses of 10 mg/kg given within 7 days, with MCK as the main affected cytosolic isoform [100]. In another study with rats, total CK activity decreased significantly between 1 and 6 weeks following a 13 week DXR treatment (2 mg/kg per week) [101]. A 15% decrease in activity of sMtCK has been found in mitochondria from mice treated with 20 mg/kg of DXR for 5 days [40]. Sarcomeric MtCK occurred to be more sensitive to inhibition as compared to the respiratory chain complexes, which would make this kinase a prime target of DXR-induced damage in vivo. Also in mice, 5 days after injection of DXR (20 mg/kg), Mihm and colleagues [102] observed a significant nitration of the myofibrillar CK isoform, MCK, accompanied by an about 30% reduction in its activity.
Long-term CK damage could be at the origin of numerous deleterious processes that promote chronic DXR-induced cardiac dysfunction [2,9]. Compromised MtCK functions, including inactivation, dimerization and inhibition of its binding to cardiolipin, would not only impair the energy channelling between mitochondria and cytosol [83], but would additionally affect mitochondrial respiration. They would also contribute to destabilization of the so-called mitochondrial contact sites consisting of ANT, MtCK and VDAC [37]. It is to note that disappearance of mitochondrial contact sites has been observed in DXR-treated liver mitochondria [34]. In the view of a postulated role of MtCK in prevention of mitochondrial permeability transition [103], MtCK defects would sensitize cardiac cells to apoptosis. In the cytosol, where MCK is functionally coupled to the Ca2+-pump of sarcoplasmic reticulum, inhibition of this isoenzyme would result in Ca2+ imbalance that in turn would interfere with muscle contraction and relaxation [84] and could lead via chronic Ca2+-overload to apoptotic and/or necrotic cell death.
6 Energy-state signalling
In a normal physiological setting, perturbations in intracellular nucleotide ratios activate signalling cascades, which would trigger a coordinated response to protect the cell from a dangerous fall in energy state. In heart, AMP-activated protein kinase (AMPK) plays a crucial role in such emergency signalling. AMPK, a key energy sensor, signalling system and regulator of cellular energy substrate utilization [104,105], is activated by a fall in cellular energy state, in particular the AMP/ATP ratio, as well as oxidative stress and some other extracellular stimuli. Activated AMPK triggers catabolic pathways that generate ATP, e.g., fatty acid oxidation, and glycolysis, and downregulates anabolic ATP-consuming processes that are not essential for short-term cell survival, such as the synthesis of lipids, carbohydrates, and proteins.
Recent work of our laboratory suggests AMPK as a new and highly sensitive target of DXR-induced damage in heart [94]. The toxicity limit in term of DXR dose and time of onset is very low. In the model of isolated rat heart, we have shown that 1-h perfusion with 2 μM DXR, which corresponds to peak plasma concentration of DXR in patients, downregulates AMPK protein, phosphorylation and activity on its downstream target acetyl-CoA carboxylase. In addition, compromised AMPK signalling was observed earlier than changes in myocardial function. The mechanisms of inhibition are not yet known, but they may include direct effects on AMPK, as well as compromised upstream signalling.
DXR induces both, energetic imbalance and generation of reactive oxygen and nitrogen species [3,93]. Under these conditions, downregulation of the fuel-sensing AMPK, which would be expected to initiate responses preventing such energy depletion, seems paradoxical. On the other hand, AMPK inhibition is consistent with several DXR effects reported earlier. Suppressed AMPK activity could explain a change in the profile of energy substrate utilization after DXR treatment, first of all a decreased fatty acid oxidation as observed in different studies [71,73]. Inactivated AMPK would increase acetyl-CoA carboxylase activity, which enhances inhibition of carnitine palmitoyl transferase (CPTI) by malonyl-CoA and thus leads to decreased mitochondrial fatty acid oxidation. Similarly, impaired AMPK signalling would decrease glycolytic rate by inhibition of PFK and limited stimulation of glucose uptake. Related to this scenario, it has been shown that AMPK mutations that possibly affect the AMPK sensing function, are at the basis for some types of familial hypertrophic cardiomyopathy [106]. In contrast, in cardiac pathologies other than DXR-induced, acute or chronic energy depletion trigger AMPK activation, as it would be expected [8]. Here, activated AMPK increases glucose uptake both in the case of short and long-term energy depletion. However, at least in chronic heart pathologies, active AMPK often fails to enhance fatty acid oxidation, probably due to a generalized decline in the capacity of the fatty acid oxidation pathway [8,107].
7 Conclusion
Alterations in myocardial energy metabolism have been recognized as a hallmark for different cardiac pathologies. This review summarized the amazing spectrum of changes in cardiac energy metabolism caused by DXR. DXR induces damage at various steps of cardiac energy metabolism, including a fall of basal high energy phosphate levels, PCr and ATP, a reduction of oxidative capacity of mitochondria, changes in the profile of energy substrate utilization with marked reduction of fatty acid oxidation, a disturbance of energy transfer between sites of energy production and consumption by the CK reaction, as well as defects in the AMPK signalling pathway. It appears that such changes in energy metabolism, typical for cardiac pathologies, belong to the very early manifestations of the DXR cardiotoxic action and does not develop secondary to other alterations, e.g., as a consequence of a failing myocardium. Finally, defects in bioenergetics can also affect the sensitivity of cardiac cells to apoptosis, as has become evident in studies with DXR-treated cardiomyocytes (see [2]).
It is of interest that in DXR-treated patients, structural changes in heart tissue appear much earlier than any clinical manifestation of heart failure. This implies that compensatory mechanisms are operative that can maintain cardiac function for a certain period of time, in spite of increasing myocardial damage. We support the view that an accumulation of various deficiencies in high-energy phosphate metabolism, involving also the failure of regulatory and compensatory mechanisms, may be a very critical step for the deterioration of cardiac function and for the onset of chronic clinical cardiotoxicity.
Acknowledgements
The related work from the author's laboratory was supported by Marie Heim-Vögtlin Subsidy from the Swiss National Science Foundation (Grant 3234-069276 to M.T.-S), Schweizerische Herzstiftung (to T.W. and U.S.), Wolfermann–Nägeli–Stiftung (to U.S. and T.W.), Schweizer Krebsliga (to T.W. and U.S.), Zentralschweizer Krebsstiftung (to U.S. and T.W.), and Novartis Stiftung für medizinisch-biologische Forschung (to U.S.).