

1 Introduction
How can abstract goals be transferred into appropriate sequences of movements? What are the neural structures where the representation is formed prior to execution of action? What is a motor centre? The history of the concept of action representation starts at the end of the 19th century, when motor physiology was dominated by the sensory-motor theory of action generation. Voluntary actions were first considered as a variety of reflexes, with some degree of autonomy with respect to external stimuli. Indeed, if actions were to be generated from within, their generation should require the existence of an internal state where they would be encoded, stored and ultimately performed independently of the external environment: this requirement for an internal state (a representation) was far from clear in the early days of physiology.
In this paper, we will examine some of the major issues raised by the origin of actions. One of these issues was the existence of excitable areas in the cerebral cortex. Although it was soon recognized that these areas were connected, via the pyramidal tract, to the spinal cord and ultimately to the muscles, the way they were fed by motor ideas to give rise to actions was difficult to conceive. For some, this was an automatic process where consciousness took no part. For others, the emission of motor commands was a conscious process, which gave rise to the sense of effort and the sense of the position of the limbs. Observations made about consciousness of action in patients with anaesthetized or paralyzed limbs sometimes added confusion to the debate, mainly because the observers only considered distal aspects of motor performance, rather than its more central ones. Action generation may not be explained by purely sensorimotor mechanisms, those where a causal relationship between an immediate cause and a visible effect can be identified. Instead, the solution to this problem is to be looked at a more elaborated level, where actions are considered, not as mere responses to a stimulus, but as the expression of cognitive states. These issues encompass a wide range of technical as well as theoretical problems in neuroscience, psychology, and even philosophy.
2 The concept of motor centres. From Hitzig to Cushing
The concept of motor centres slowly emerged during the second half of the 19th century from the work of physiologists and clinical neurologists who made direct observations on animals and in human patients. In 1870, Eduard Hitzig with his colleague Fritsch discovered the effects of direct electrical stimulation of the precentral cortex in dogs [1]. These experiments revealed the existence of an ordered (somatotopic) organization of the cortical representation of movements. According to the authors, “a part of the convexity of the hemisphere […] is motor, another part is not motor. The motor part, in general, is more in front, the non motor part more behind.” [1 (p. 81)] Fritsch and Hitzig took their result as a demonstration that “some psychological functions and perhaps all of them, in order to enter matter or originate from it need certain circumscribed centres of the cortex” (p. 96). In spite of this unambiguous finding, Fritsch and Hitzig interpreted their own results along with the current notion of kinaesthetic images (see [2]). The ablation of these areas, they claimed, provoked paralysis because of the destruction of “motor images” of the voluntary movements. The animal was unable to perform movements, not because of paralysis of the muscles, but because of the impossibility to represent these movements. Fritsch and Hitzig considered that the excitable part of the cortex was an intermediate between “that part of the brain that harbours the origin of the volition of the movement and lower muscular mechanisms” [1 (p. 92)]. Ferrier, who also had undertaken stimulation experiments, first on dogs and then in monkeys, also considered the motor centres as distinct from the cortical zones where sensations, including muscular sense, were represented [3]. His conception of the origin of movements was purely sensory-motor: sensory tracts carried impressions to the sensory centres in the cortex, which were connected with motor centres, which in turn projected down to the muscles. The motor centres were “the centres for voluntary initiation of the same movements as result from faradization” [4]. The fact that dogs in which the motor areas had been destroyed shortly recovered quasi-normal movements, as observed by many authors, was explained by Ferrier by the role of basal ganglia, which he considered as centres for automatic movements, as opposed to the voluntary nature of the movements produced by the cortical centres. What was true for dogs, however, did not apply for monkeys, in which the paralysis persisted for much longer time.
The contribution of clinical studies to the definition of cortical motor centres was also of a great importance. At the end of the 1860s, Hughlings Jackson had described patients with unilateral epileptic spasm which he attributed to irritating or discharging lesions of the cortex, and had called attention on cortex as the possible source of movements. His insight into cortical organization, based on the sole observation of the ‘march’ of the epileptic seizure, is surprisingly accurate. His conclusion was that “the part of the body where the convulsion begins indicates the part of the brain where the discharge begins, where the discharging lesion is situated. But from the focus discharging primarily, the discharge spreads laterally to the adjacent ‘healthy’ foci. […] The order in which the different parts of the body become involved, reveals the arrangement of the corresponding foci in the precentral convolution” (quoted by O. Foerster in his Jackson Lecture [5]). Indeed, Ferrier explicitly acknowledged the fact that his own experiments were aimed at testing Jackson's theory. In France, Jean-Martin Charcot, although he apparently ignored the work of Fritsch and Hitzig, warmly acknowledged the influence of both Jackson and Ferrier on his own thinking. Yet, his map of motor cortex, based on post-mortem examination of patients with motor impairments like hemiplegia or monoplegia, still bears the imprint of the kinaesthetic images theory: his motor area of the cortex includes both the precentral and the postcentral gyri. This may not be surprising, due to the frequent association of motor and sensory disorders following vascular accidents involving the sylvian artery in the patients that were observed by Charcot [6,7]. Charcot's description was also influenced by an erroneous interpretation of Betz findings about the repartition of giant pyramidal cells in the cerebral cortex [8]. These cells, which were considered as the origin of the pyramidal tract, had been clearly assigned by Betz to the anterior central gyrus, but Charcot thought that they were spread over both side of the sulcus [9].
Finally, the modern conception of a motor cortical area was firmly established by Sherrington by electrical stimulation experiments in apes [10,11]. The electrically responsive zone corresponded to the cortical area where the giant pyramidal cells described by Betz were observed, and where the corticospinal pyramidal tract took its origin. Holmes and May [12] studied the retrograde degeneration of Betz cells in monkeys with unilateral section of the pyramidal tract: the cell loss was selectively localized in the infragranular layer of the cortex in the precentral gyrus opposite to the section, with no involvement of the postcentral gyrus. The same results were also obtained in two patients with accidental sections of the spinal cord. Later on, Cushing was able to demonstrate, by systematically using the method of electrical stimulation in man, that the sensory cortex was located posterior to the central fissure: during the stimulation, the patients reported cutaneous feelings located in specific body parts (see Fig. 1). The correspondence of the localization of the feeling and the stimulated zone allowed Cushing to establish the cortical map of somatic sensations [13]. Cushing's findings were made in epileptic patients. During the stimulation of the post-central area, the feelings reported by the patients corresponded both in their localization an in their nature, to the aura that the patients experienced prior to their epileptic seizures. These findings were to be amply confirmed and expanded by Penfield and his colleagues [14]. Yet, although the problem of the existence of the motor centres was solved, the origin of the actions that were mediated by these centres still remained a mystery.

Per-operative map of human sensorimotor cortex. Anterior to the central fissure, Xs indicate motor responses (flexion of the thumb or contraction of vocal cords). Posterior to the fissure, Ss indicate sensations elicited by the electrical stimulus and referred by the patient to the back of his hand or his index finger, according to the stimulated zone. This per-operative map is the first description of human sensorimotor cortex using the electrical stimulation method. From Cushing, 1909.
3 From the muscular sense to the feelings of innervation. James against Wundt
Parallel to the evolution of ideas on the motor centres, another debate was running about the nature of the signals that could be used to generate actions and monitor them consciously. One of the early formulations of the opposition between the tenants of peripherally controlled and centrally controlled actions can be tracked in the discussions about the origin of the muscular sense. We feel, in one way or another, that we are moving. Movements trigger sensations. Charles Bell's formulation can be used to set the terms of the debate: “At one time, he said, I entertained a doubt whether this (the consciousness of the positions of our limbs, or ‘muscle sense’) proceeded from a knowledge of the conditions of the muscles or from the consciousness of the degree of effort which was directed to them in volition” (quoted by C. Phillips in his Sherrington Lecture [15]; see also [16]). These two opposite conceptions of position sense were deeply rooted in the classical 19th-century physiology. The peripheral conception, before it was heralded by Sherrington and became for a time the dominant theory (see below) was considered by many authors as unsatisfactory because of a lack of experimental evidence. Waller, for example, said that muscle sense, “in the sense of centripetal process from muscle is not supported by any direct proof, and so long as the alternative hypothesis of expended energy in ‘motor’ centres is not disproved, it is not possible to admit that the feeling is entirely of peripheral origin, nor that the muscular contribution is the predominant factor among its peripheral constituents.” [17]
The hypothesis claiming the existence of an ‘expanded energy’ of a central origin, playing the role of a signal for the sense of position of the limbs had strong proponents. One of them was Alexander Bain: “The sensibility accompanying muscular movement, Bain stated, coincides with the outgoing stream of nervous energy, and does not, as in the case of pure sensation, result from any influence passing inwards by in-carrying or sensitive nerves.” [18] (See a discussion of this point by Lewes [19] and Bastian [20 (pp. 336–337)].) Bain thought that there was a feeling of the exerted force and that this feeling was the “concomitant of the outgoing current by which the muscles are stimulated to act”. This theory originated from several sources. One of them was clinical observations of patients with complete anaesthesia of one limb (including the loss of sensations generated by passive displacement), who were still able to produce voluntary movement with that limb. According to Duchenne de Boulogne [21], these patients still presented “muscular consciousness” (conscience musculaire), although they had lost muscular sense, that is, sensations generated by muscular activity. Duchenne was therefore led to the interesting conclusion that muscular consciousness could exist independently of muscular sensations. This point was clearly formulated by Lewes [19], who introspectively distinguished between the “motor feeling” accompanying the active contraction of a muscle and the “sensation” generated by this contraction. Lewes thus considered that the complex experience arising from a voluntary movement was the sum of both the “sense of effort” and the “sense of effect”.
At one time, the debate on peripheral vs central origin of action generation and monitoring became known as the “Two Williams debate”, a term that epitomized the controversy between William James and Wilhelm Wundt around 1890. William James defended the opinion that the consciousness of our movements is based a posteriori on information from sensory organs, whereas Wilhelm Wundt, on the contrary, held that our knowledge is based a priori on efferent information of a central origin, i.e., in other words, that we can perceive a specific feeling, which he called the feeling of innervation (Innservationsgefühl), when we produce a movement. Wundt was a proponent of the notion that the mind has a mental content. This content is the result of mental processes, in the sense that the mind is actual and not substantial: it has no substance as a fixed, static conception would hold, it is not an object, but a process (Wundt's theory of actuality, see [22]). Wundt's ideas of mental content and mental processes were at the origin of the methodological concept of mental chronometry, still in use nowadays. Mental chronometry refers to the fact that mental processes take time to build up and the observed delay before giving a response that involves mental processes reflects the contribution of the corresponding central mechanisms. It is therefore not surprising that Wundt favoured the existence of central efferent processes accounting for the generation and conscious monitoring of action.
In the late 1880s, however, the most frequently held concept accounting for the production of action was that of ‘kinaesthesia’, introduced by Charlton Bastian. According to this concept, kinaesthetic images were formed from sensory traces left by prior movement, stored in motor cortex, and reactivated when the same movement was executed again [23]. Bastian was influenced by his friend Herbert Spencer who thought that sensory impressions arising from the muscles during a movement were revived when the same movement had to be produced [2]. Thus, the cortical centres for movements were actually sensory centres where kinaesthetic images could be stored: lesions of these centres produced paralysis because they made the patient unable to recall the appropriate movement patterns. In his words, the patient had lost the “muscular consciousness” (a concept quite different from that of conscience musculaire proposed by Duchenne de Boulogne to explain the effects of peripheral anaesthesia, see above). Bastian's theory was shared by Hermann Munk, who claimed that the so-called “motor cortex” in fact “consisted of organs at the service of a special memory, the memory of movements, and constituted a sort of cerebral register that stored the information acquired by the sensory organs” [24]. Bastian found his main support in the results obtained by Charles Sherrington in a famous monkey experiment with Mott [25] showing that the suppression of sensory input from one limb produced a paralysis of that limb, that is, the same effect as the destruction of the cortical centre (see [20]). Mott and Sherrington had indeed observed that, following a section of the dorsal roots on one side, which suppresses sensory input from the corresponding territory to the central nervous system, the deafferented limb became useless and almost paralyzed. The animal could only produce awkward movements with that limb when forced to use it. Hence Mott and Sherrington concluded that movements owed much to the periphery for what concerned both their initiation and their execution. This view was maintained throughout the first half of the last century until deafferentation experiments were repeated by Bizzi and his colleagues in the late 1960s. They showed that a monkey with bilateral deafferentation of the forelimbs could perform reasonably accurate monoarticular pointing movements directed to a visual target, in the absence of vision of the limb. The entire structure of the movements was preserved, including, not only their initial, ballistic, phase but also their low-velocity phase up to the endpoint [26].
In the Two Williams Debate, James position was partly influenced by Bastian. His main line of thought was that “voluntary movements must be secondary, not primary functions of our organism.” [27 (II, p. 487)] In order to perform voluntary movements, we need first to experience the effects of involuntary performance. “When a particular movement, having once occurred in a random, reflex, or involuntary way, has left an image of itself in the memory, then the movement can be desired again, proposed as an end, and deliberately willed.” [27 (II, p. 487)] Ideas of movement can be remote effects of previous movements, or resident effects, those that are directly produced by the body parts that are actually moved. James considered the latter as equivalent to Bastian kinaesthetic impressions, arising from muscles and joints (although he confessed his ignorance about the sensory organs that might generate these impressions). James based his conception on observations made in patients with various forms of anaesthesia, including Strümpell's “wonderful anaesthetic boy”, whose only sources of feeling were the right eye and the left ear (see James, 1890, II, pp. 489–490). This patient, when blindfolded, had no sense of the position of his limbs, no sense of fatigue, although he could correctly execute movements on command. Yet, if his intended movement was prevented, he was not aware of not having moved and was surprised, when he opened his eyes, to see the position of his limb.
Note that the interpretation of the effects of anaesthesia differs according to authors. Bastian considered anaesthesia as a cause of paralysis. James was ready to accept that an anaesthetized patient was not paralyzed, but was simply unaware about whether his intended movement had taken place or not. The effects of anaesthesia of pathological origin were going to become a key argument for determining the respective roles of sensory afferences and central commands in action generation and consciousness. In his discussion about action consciousness, James uses the case of another patient with anaesthesia of both arms reported by Gley. This patient could not determine the difference between objects of different weights that he was asked to lift. In other words, he was unable to use the feelings derived from his effort to estimate the weight, hence demonstrating, according to James, his inability to consciously monitor his feelings of innervation. At the beginning of the 20th century, Karl Lashley reported the case of a patient with a deafferented leg following a gunshot injury of the spinal cord. Despite the complete absence of sensations from that leg, the patient was capable, even when blindfolded, of bending his knee at a given angle, or placing his foot at a height indicated by the experimenter [28]. In subsequent papers, Lashley noted that a great number of our movements are executed too rapidly for any sensory control to intervene. He pointed out that, during the playing of a musical instrument, for example, finger alternations can, in certain instances, attain the frequency of 16 strokes per second, which exceeds the possibility for any sensory feedback to influence the command system. Thus, the succession of such rapid movements must be centrally encoded before they are executed [29]. Further clinical observations, since Lashley, have confirmed this point of the independence of the central command from the periphery. A patient suffering a severe sensory neuropathy, and who had lost all somatosensory cues from his limbs was studied by Rothwell et al. [30]. In spite of his sensory impairment, this patient, when blindfolded, was able to perform a wide range of motor tasks such as tapping, fast flexion extension movements of the elbow, or drawing figures in the air. Furthermore, the EMG pattern of these movements was closely similar to those observed in a normal subject. Through these accounts of the effects of anaesthesia reported by different authors, a distinction seems to have to be made between two aspects of the motor function in the absence of sensory input, namely, “being able to make movements” and “feeling one's own effort”. Concerning the former aspect, the role of sensory input has been ruled out after the demise of the Mott and Sherrington claim (see above). The latter, however, still remains a matter of discussion. Recent experiment in another deafferented patient reports that she can correctly calibrate her own muscular force to match a given amount of force, despite the fact that she has no overt feeling of the force involved, and does not experience fatigue when a high degree of muscular contraction is required [31,32]. Coming back to the Two Williams debate, it is the existence of the feeling, and not the ability to make movements which was at the core of the discussion between James and Wundt.
The theory of the feelings of innervation defended by Wundt [33] was severely attacked by James. “At the moment when we consciously will a certain act, a mental conception made of memory images of these [kinaesthetic] sensations, defining which special act it is, must be there.” [27 (II, p. 492)] “Now, James continues, is there anything else in the mind when we will to do an act?” [27 (II, p. 492)] His answer to this question is negative, in spite of the fact that “a powerful tradition in Psychology will have it that something additional to these images of passive sensation is essential to the mental determination of a voluntary act.” [27 (II, p. 493)] This additional element is a feeling attached to the current of energy going from the brain into the appropriate muscles. “This feeling of the current of outgoing energy has received from Wundt the name of the feeling of innervation. I disbelieve in its existence.” [27 (II, p. 492)] James arguments for denying the existence of feelings of innervation were twofold. First, he thought, they are useless, because we do not need to know how our actions are performed: on the contrary, “consciousness deserts all processes where it can no longer be of use.” [27 (II, p. 496)] “By virtue of this principle of parsimony in consciousness, the motor discharge ought to be devoid of sentience.” [27 (II, p. 497)] “We ought then, on a priori grounds alone, to regard the Innervationsgefühl as a pure encumbrance, and to presume that the peripheral ideas of movement are sufficient mental cues” [27 (II, p. 498)]. The second argument James used to deny the existence of the feelings of innervation is that there is no introspective evidence for them. This argument was a direct answer to Wundt who had used clinical reports about the effects of partial paralysis in support to his own theory of conscious monitoring: “A patient, Wundt said, whose arm or leg is half paralyzed, so that he can only move the limb with great effort, has a distinct feeling of this effort: the limb seems to him heavier than before, appearing as if weighted with lead.” [27 (II, p. 503)] James replied that such observations were incorrect in that they relied on an incomplete inventory of the afferent data: when attempting to move his paralyzed arm, the patient could have involuntarily contracted other muscles as a consequence of his efforts, and these contractions could have been felt as an increased effort.
The quarrel between Wundt and James was a reflection of the opposition between two schools of thoughts. However, the discussions bore on one particular aspect, which was the role of conscious sensations from either peripheral or central origin, in the generation of movements. The main reason why James proposed that feelings of one's own movements are generated a posteriori was because he assumed that actions are generated unconsciously. The second part of his chapter on Will, on “Ideomotor action” is devoted to discussing this point (see below). In that sense, the issue of the conscious will may appear as an ill-posed problem, for which there is, perhaps, no answer to be expected. The real issue was in fact about whether the generation of a movement can be an endogenous process, i.e., independent from sensory input; and to this point, Wundt's contribution was essential [33].
4 Central mechanisms for action representation. Liepmann's contribution
Assuming the existence of voluntary actions generated in the absence of sensory input does not solve the problem of how these are centrally organized, or represented. As we now realize, the debate about whether movements were of a central or peripheral origin was in fact a debate about the nature and the content of motor representations. What we learn from physics is that even the simplest feedback regulation systems seem to require a minimal amount of representation of their internal state, against which the regulation can be exerted. This notion was already familiar to engineers (if not to biologists) during the early part of the 19th century [34]. Regulation of steam engines (by “governors”) implied a reference-state that the system was supposed to reach and maintain. Biological systems later appeared to be liable to the same mode of functioning. Claude Bernard, in his lessons published in 1878, made the point that systemic regulations were circular mechanisms aimed at maintaining the constancy of the internal milieu. Regulation of blood glucose for instance was based on constancy of glycaemia (the reference-state) at a level corresponding to metabolic needs. When glycaemia dropped below its reference value, processes were activated to restore the reference state. This mode of regulation can therefore be schematized as a predetermined system with an input stage (receptors), an output stage (effectors), and a central stage (the reference) that detects errors between reference and input and activates or deactivates the effectors. One of the important points in Claude Bernard's description was thus the discovery that self-regulating systems proceeded from a teleonomic conception of the representations [35].
This idea of self-regulation was also inherent to the concept of homeostasis, which received a broad recognition among biologists early in the 20th century, and was used for explaining many different physiological functions. Homeostatic systems are classically considered as closed-loop systems aimed at maintaining a fixed internal reference. Their activation is thus automatically related to the monitoring of an error between their input and central stages. Interestingly, some of these characteristics seem to be present in the functioning of motor reflexes. It was Pflüger who first expressed the idea that spinal reflexes in animals with section of the spinal cord reflected a form of consciousness of the spinal cord (Rückenmarkseele [36]): they were ‘purposive’, in the sense that they were apparently organized so as to preserve the integrity of the animal organism in response to aggressions (see [37,38]): reflexes like the pupillary response to light, postural reactions, the vestibulo-ocular reflex, function as self-regulating systems for compensating discrepancies between their central stage and their input stage.
This discussion therefore suggests no discontinuity in the mode of regulation of movement types. References and goals should be considered as different levels of motor representations. The hierarchical status of each given level of representation would only depend on the degree of independence of the corresponding movements with respect to the external milieu. The prerequisite for accepting the possibility of voluntary actions independent from the environment is that they are preceded by a representation that can be formed in the absence of sensory input, so that, when the action comes to execution, the representation can provide the elements necessary to perform it. Bastian, James or Munk had already proposed the notions of “mental conception of action”, or “motor idea”, to account for the role of memory images or remote impressions in shaping an action. However, the modern thinking about action representations came from a different background. It was Hugo Liepmann, starting from the background of clinical neurology, who made the necessary steps forward, first by discarding the issue of action consciousness and second, by abandoning the opposition between peripheral or central origin of the action. Instead, he proposed a completely new scheme in concentrating on how an action can be assembled from its elementary constituents. According to Liepmann, an action must proceed from an internal ‘plan’: “The main representation of the goal can only be reached if a plan (Entwurf) is built internally, concerning the direction, contiguity, succession and rhythm of the elementary acts.” [39 (p. 45)] To account for the implementation of the plan, he proposed that the elementary bits of the action were assembled according to the main representation: the result of this process was what he called a “movement formula” (Bewegungsformel), that is, an anticipatory hierarchical structure where all the aspects of an action were represented. Following the lead of Karl Wernicke, Liepmann presented his model as a diagram where the smaller elements were connected into a larger formula. This revealed a useful method for explaining pathological action impairments observed in brain-lesioned patients. Liepmann's patients were impaired in executing simple everyday actions on command, in the absence of motor paralysis or sensory deficit. Such ‘apraxic’ impairments had been included by his predecessors (e.g., Nothnagel, 1887, quoted by Liepmann [40]) into the broader group of psychic paralysis: Liepmann kept the term of apraxia which he re-qualified with detailed clinical descriptions: according to the level at which the movement formula was defective, patients presented either with apraxia of a ‘motor’ type (later called “ideomotor”), when the disconnection was between the central and the execution level, or apraxia of an ‘ideatory’ type, when the disconnection was between the elements of the movement formulas, within the central level (see [41]).
Liepmann was greatly inspired by the observation of a single patient who had suffered a cerebrovascular accident in December 1899. In his 1900 reports of this case, Liepmann had noted that the patient failed, with his own right arm or leg, to produce simple movements on verbal command and to imitate movements performed by another person. Motor performance with the left limbs was apparently correct [40]. When the patient died 16 months later, his autopsy revealed the existence of bilateral lesions in the inferior parietal lobule, predominating on the left side, and a large softening of the corpus callosum [42]. Liepmann's interpretation of this and many other cases that he reported in another paper in 1905, was that the brain area responsible for these motor impairments (the sensomotorium) was located in the left hemisphere. As a rule, Liepmann noted that left sided lesions produced a right hemiplegia with a limb apraxia of the left side, whereas right-sided lesions only produced a left hemiplegia with the right side unimpaired [43]. The interpretation given to this asymmetry was that the left cortical centre was dominant for action representations and sent its commands to the right hemisphere via the corpus callosum. When the left centre and/or the corpus callosum were severed, apraxia appeared. According to the model of a control of representations for both sides by the left hemisphere, apraxia resulting from lesion of the left centre should be bilateral, but the existence of the right hemiplegia usually masks the right-sided apraxia. Note that the first Liepmann patient, who had no hemiplegia, may be considered as an exception to the rule, as his apraxia was apparently limited to the right side. In 1920, Liepmann published his famous diagram (Fig. 2) showing the various possibilities of intra-hemispheric disconnection and their clinical consequences. This diagram bears the imprint of the associationist thinking that was the rule in Wernicke's school. Following lesions interrupting the association pathways between the left centre and other brain areas where sensory information is processed, the representation of the action was no longer possible.
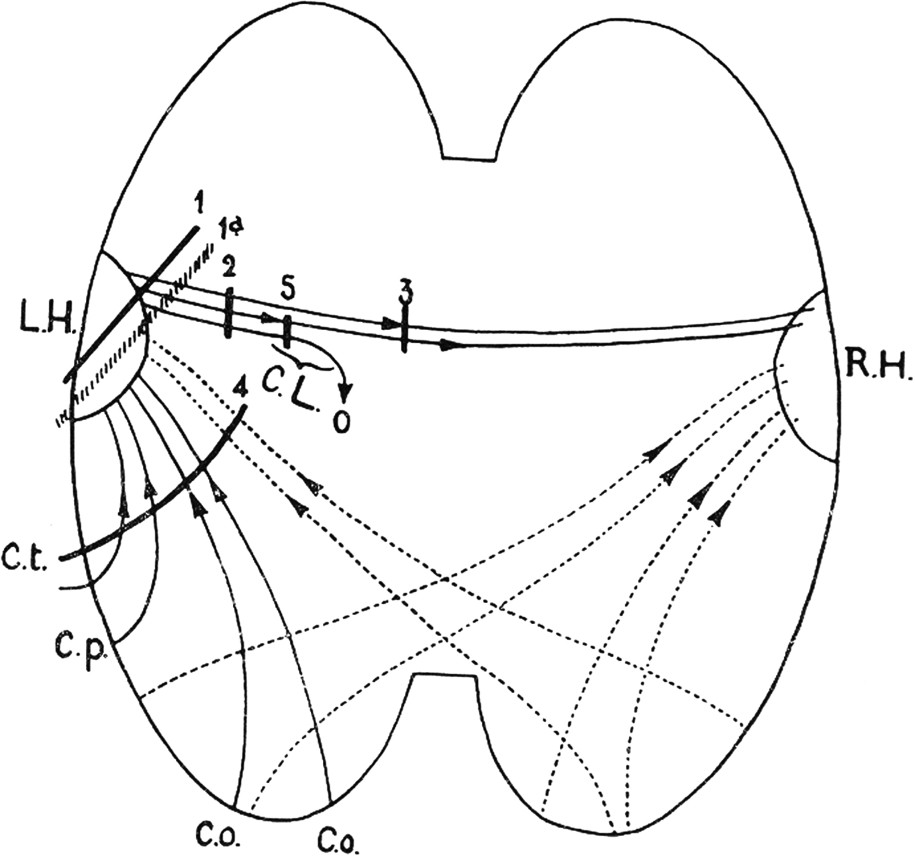
One of the Liepmann' diagrams showing the intra-cortical connections responsible for the representation of actions. The left cortical centre (LH) for the right hand receives connections from occipital, temporal and parietal sensory areas (Co, Cp, Ct). This centre sends commands to the right hand through the pyramidal tract and to the left hand via its connection through the corpus callosum with the right cortical centre (RH). Lesions in 1 and 2 produce a right hand paralysis and a left hand dyspraxia. Lesions in 3 produce a dyspraxia of the left hand. Lesions in 4 produce a bilateral ideomotor apraxia. Lesions in 5 produce a paralysis of the right hand and no dyspraxia of the left hand. From Liepmann, 1920.
Liepmann's legacy is still quite influential in neuropsychology and cognitive neuroscience of action. Later authors replaced the term of movement formula by those of “engram” [44], “schema” [45], or “internal model” [46], but kept the notion of a representational level with a hierarchical organization (see [47]). Bernstein had an interesting analogy for explaining this mode of organization that can be a valid conclusion for this paper. He thought that the representation of an action must contain, “like an embryo in an egg or a track on a gramophone record, the entire scheme of the movement as it is expanded in time. It must also guarantee the order and the rhythm of the realization of this scheme; that is to say, the gramophone record […] must have some sort of motor to turn it.” [47 (p. 39)]