

1 Introduction
Fire determines overall vegetation structure and composition of several biomes of the world, as well as temperature, precipitation, and water balance [1]. The fire-prone vegetation cover currently constitutes 40% of the world's land surface, including four Mediterranean regions [2,3]. In Mediterranean-type ecosystems, most shrublands have the climatic potential to be forest and are generally fire-maintained [1]. Although, in contrast with current opinion that often attributes the existence of these biomes to anthropogenic burning, Mediterranean shrublands seem to have naturally expanded in the Late Tertiary, with flammable C4 grassy biomes [4]. In the Mediterranean Basin, natural settlement of Pine forests has also played a determinant role in increasing fire frequency since the Miocene (ca. 9 Myr) [5]. Forest fires are generally considered as catastrophic events in this region, but the evolutionary significance of fire for driving landscape diversity, ecosystem heterogeneity, and species differentiation constitutes another important aspect (e.g., [6–8]). Fire indirectly induces environmental changes through alterations of temperature and oxygen concentration in soils, increases resource availability, and reduces plant competition [9]. In fire-prone environments, fire represents also a major selective force that shapes the evolution of plant reproductive traits such as resprouting ability and propagule persistence, and it can stimulate flowering or germination, which favour species persistence [9–11].
However, most of the studies focused on post-fire recovery of woody plants, and adaptations to fire of other life forms are less examined (but see [12,13]). This is the case for the geophytic flora of Mediterranean-climate ecosystems, which represents an original component of the Mediterranean biodiversity, notably in the Cape Flora of South Africa [3,14,15]. Within the Mediterranean Basin, we can estimate that ca. 1500 geophytes species occur, i.e. 8% of the total flora (Médail, unpubl.). If the geophytes of the Mediterranean Basin are mostly rare and narrow endemic species, their persistence by longevity through seeds and storage organs often reduces their local extinction [16]. A detailed analysis of patterns of plant rarity in southeastern France also suggests that bulbous and tuberous geophytes are more prone to rarity and extinction than rhizomatous geophytes [17].
The remarkable geophyte diversity that occurs within these Mediterranean ecoregions is probably linked to the pronounced rainfall seasonality and the occurrence of fire, which, respectively, promote resource acquisition and limit the competition from shrubs and grasses [15]. Geophyte abundance could be also attributed to their adaptation to fire, with a dormancy period corresponding to the fire season [9]. During this dormancy phase, aboveground tissues die back and subterranean storage organs are poorly affected if fire intensity and severity is reduced. Furthermore, characteristics of several geophytes such as an important seed bank [18], or fire-stimulated flowering [19] may contribute to the persistence of these species after fire events. Nevertheless, ecological consequences of fires on rare plants are poorly investigated in the Mediterranean Basin, and patterns of post-fire demography of endemic geophytes are still unknown, despite their often-high conservation value and their narrow distribution range.
In this context, we examine the effects of a natural fire on the demography of an endangered and narrow endemic geophyte, Acis nicaeensis (Ardoino) Lledò, Davis and Crespo (Alliaceae). This study was realized to answer the following questions. (1) What are the characteristics of the A. nicaeensis' habitat, i.e. the Mediterranean xerophytic grassland vegetation, after fire? (2) What are the effects of fire and/or surrounding vegetation on demographic stages and reproductive success of A. nicaeensis? Then, we explain why the processes induced by this disturbance constitute an important topic to be integrated within a conservation strategy for spatio-temporal persistence of threatened geophytes in Mediterranean fire-prone environments.
2 Materials and methods
2.1 Species and study area
Acis nicaeensis (Ardoino) Lledó, Davis and Crespo (previously Leucojum nicaeense Ardoino) is a narrow endemic and rare geophyte from the southern part of the Maritime Alps (Fig. 1), between the ‘Col de Vence’ and the French–Italian border near Menton, with a reduced and patchy distribution () on some calcareous rocky grasslands [20]. This geophyte is characterized by one (rarely two or three) white flower(s), with a style slightly exceeding six stamens, black seeds with a strophiole, and a bulb as a storage organ. The plants possess a relatively high capacity for vegetative propagation. The bulb is subterranean and can be radially swollen to form a daughter bulb when it matures. Bulb development appears to be tied to growth of the aerial organs, which occurs during the winter until the beginning of the spring growing season. In this area, flowering takes place in mid-April and seed is set around six weeks later. Irrespective of whether flowering or not, aerial organs die back to ground level at the beginning of the summer. Seeds are dispersed through barochory, but myrmecochory can occur sporadically [20]. As with many Mediterranean geophytes, an important bulb bank (i.e. bulb in dormancy) and seed bank is stored in the soil (Fig. 2).

Distribution map of Acis nicaeensis, location of the study site (black arrow), and photo of a flowering individual in the burned area of the ‘Baou de Saint-Jeannet’.
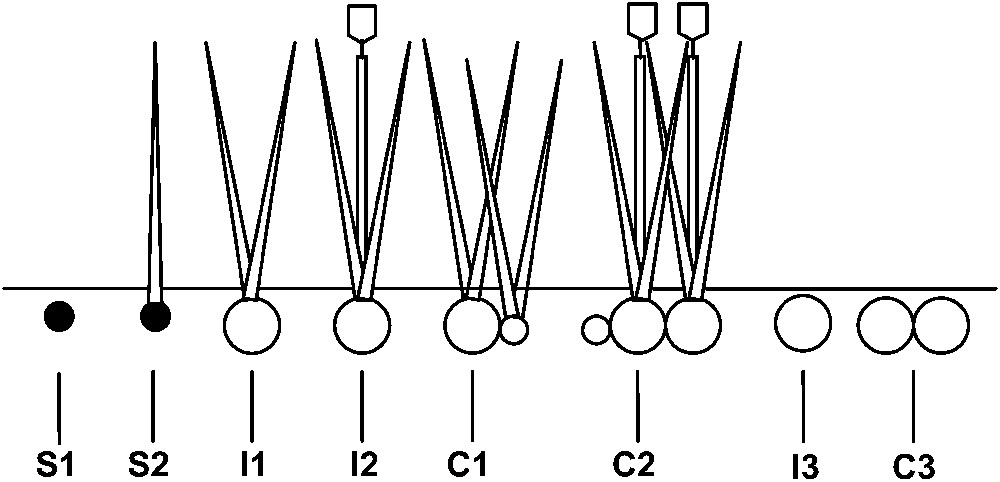
Demographic stages of Acis nicaeensis. S1: Seed, S2: seedling, I1: non-flowering individual, I2: flowering individual, I3: individual in dormancy, C1: vegetative clump, C2: reproductive clump, C3: clump in dormancy.
This study was conducted in a 3-ha site, on the southeastern slope of the ‘Baou de Saint-Jeannet’ (5.34/48.615 gr) on a 20–25° slope. This small calcareous mountain (222 ha) is situated on the western course of the Var River (‘Alpes-Maritimes’, southeastern France), under a meso-Mediterranean bioclimate. This population is known at least since 1899 [21], and established on a surface of . The principal vegetation type is a xerophytic grassland [22] characterized by Thymus vulgaris L., Lavandula angustifolia Mill., Genista hispanica L., Fumana ericoides (Cav.) Gand., Bromus erectus Huds. and Helianthemum oelandicum (L.) Dum. Cours., with sparse woody plants (Quercus ilex L., Q. pubescens Willd., Juniperus phoenicea L. and Pistacia lentiscus L.). The substrate is dominated by calcareous rocks, within a more or less karstic environment. In August 2003, a rapid and small unintended fire burned 3.5 ha of the surface where the population was established (source: Prométhée database). Based on soil and vegetation [23], the study was performed on a homogeneous slope that showed no difference in mesological and vegetation characteristics before burning. In this way, unburned sites could be considered as control sites.
2.2 Field sampling
Ten permanent plots of 1 m × 1 m were randomly assigned in each treatment (unburned vs. burned). However, plots were located when at least two individuals of A. nicaeensis were present. The number of individuals within each demographic stage of A. nicaeensis was recorded in each plot of (i.e. density) and year (2004 and 2005). In addition, the reproductive success was estimated in each plot as (i) the reproductive phase success, i.e. the ratio of mature fruits, flower abortion, and fruit predation to the total produced reproductive structure, and (ii) the number of seeds produced per fruit and per plot. Moreover, the asexual reproduction was also recorded, taking into account the number of clumps and the mean number of individuals per clump (Table 1, Fig. 2). The phytoecological surveys [24] were carried out in springs 2004 and 2005 in order to compare species richness and composition between burned and unburned situations (Table 1).
Variables recorded for each plot of 1×1 m (n=20) during the study period
| Variables | Description |
| Demographic stages | Total number individual, number of non-flowering individuals, flowering individuals and seedlings |
| Number of clumps (asexual reproduction), mean number of individuals per clump | |
| Reproductive success | Proportion of fruit production, flower abortion, fruit predation (%) |
| Total seed number produced in the plot and mean seed number per fruit | |
| Floristic composition | Specific plant specie cover following the abundance-dominance of phytosociological method and census Braun-Blanquet [24] (%) |
| Vegetation height | Mean vegetation height (cm) |
| Vegetation cover | Total vegetation cover (%) |
| Substrate cover | Cover of blocks, rocks, stones, leaf litter, bare soil (%) |
2.3 Data analysis
A preliminary principal component analysis (PCA) was performed on data characterizing substrate and vegetation cover. Then, a correspondence analysis (CA) was carried out to evaluate the contribution of all the different plant species of each treatment and plot using CANOCO 4.0 [25]. One hundred and nineteen plant species and 20 plots for each year were included in this analysis. The aim of these two analyses was to visualize the distribution of plots and variables in a multivariate space and to detect vegetation and substrate differences among plots.
Treatment and year variations for mean vegetation height, vegetation cover, each parameter of substrate cover, demographic stages of A. nicaeensis, mean seed production per fruit and total seed number per plot of A. nicaeensis were analyzed with repeated ANOVAs using PROC GLM [26]. Treatments (unburned and burned) and years (2004 and 2005) were considered as fixed effects. We performed a log-linear model analysis to evaluate variations in the reproductive phase success of A. nicaeensis, under treatments and years, on the numbers of mature fruit, flower abortion and fruit predation (PROC CATMOD) [26]. Data were arcsin-transformed for proportions, log-transformed for measures and square root-transformed for numbers to homogenise the variances. Finally, correlation coefficients between demographic stages of A. nicaeensis and each parameter of vegetation and substrate covers were evaluated using PROC CORR [26].
3 Results
3.1 Vegetation and substrate characteristics after fire
The principal component analysis (PCA) based on vegetation and substrate cover shows a clear difference between plots in response to fire (Fig. 3). One-year-after-fire burned plots are grouped together, largely depending on the bare soil cover. In contrast, two-year-after-fire burned plots are mixed with unburned plots. This result indicates that the structures of vegetation and substrate are roughly similar between the two treatments, only two years after fire. The distance between burned plots, one and two years after fire, clearly illustrates a change in the vegetation structure and in the substrate composition. This change concerns a decrease of bare soil cover, and an increase of vegetation and leaf litter covers.
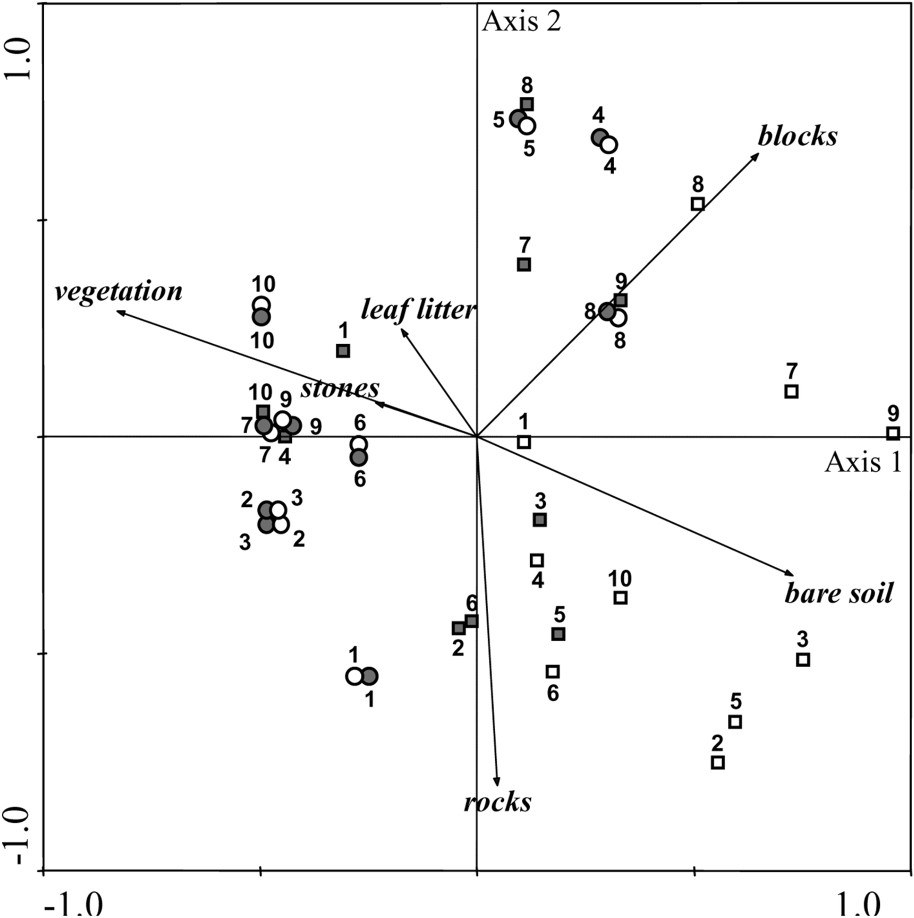
Principal component analysis (PCA) of vegetation and substrate parameters, defined by the first two axis, is used to identify changes between treatments (unburned plot: circle; burned plot: square) and years (2004: white; 2005: grey). The first two axes represent 36.7% and 28.8%, respectively, of the total variance.
A significant effect of year and a significant interaction between year and treatment occur for vegetation height, vegetation cover, and bare soil cover, but not for leaf litter cover (Table 2). In addition, a significant difference between treatments is found for vegetation, leaf litter, and bare soil covers, while no difference is found for vegetation height. This analysis reflects the between-treatment and the between-year differences found in the PCA (Table 2, Fig. 3).
Analysis of variance between year (2004 and 2005) and treatment (TRT: burned vs. unburned) for vegetation cover and height, and substrate cover. Results obtained using repeated ANOVAs (mean square: MS, ddl=1, 36) with statistical differences expressed as ; ; , and ns: not significant
| Source of variation | ddl | Vegetation height (MS) | Vegetation cover (MS) | Litter cover (MS) | Bare soil cover (MS) |
| Year | 1 | 1370** | 3.60*** | 0 ns | |
| TRT | 1 | 23.90 ns | 4.90* | 4.90*** | |
| Year ∗ TRT | 1 | 1381** | 3.60*** | 0 ns | |
| error | 36 | 181 | 0.55 | 0.23 | 0.25 |
The correspondence analysis (CA) based on floristic composition clearly discriminates the plant communities associated with the unburned vs. burned situations (Fig. 4). Vegetation plots are well structured on the CA scattergram, except for the burned plot No. 4, which is clearly separated on the first axis (10.8%). This plot is located on a deeper soil and plant composition encompasses mesophilous plants (Rubus sp., Daucus carota L., Ranunculus bulbosus L. …). The second axis of the CA (9.5%) discriminates two plant communities. This axis splits the vegetation with annual species (Logfia gallica (L.) Coss. & Germ., Hedypnois cretica (L.) Dum. Cours., Ononis reclinata L., Euphorbia exigua L., Brachypodium distachyon (L.) P. Beauv., etc.) from plant communities of the burned plots, and the vegetation of some xerophytic grasslands surrounded by a dense cover of small ligneous species (Thymus vulgaris, Euphorbia spinosa L., etc.) and herbs (Brachypodium rupestre (Host) Roem. & Scholz, Carduus tenuiflorus Curtis, etc.) from plant community characteristics of the unburned plots. In contrast with the PCA (Fig. 3), this analysis based upon floristic composition reveals clear differences between unburned and burned plots two years after fire. Nevertheless, the floristic composition of the burned plots evolves rapidly toward the plant composition of the unburned xerophytic grassland, with a shift from annual to perennial species (Fig. 4).

Scattergram of correspondence analysis (CA) on floristic composition performed on plots within treatments (unburned plot: circle; burned plot: square) and years (2004: white; 2005: grey). (Axis 1: eigenvalue = 0.447 and inertia = 10.8%; Axis 2: eigenvalue = 0.389 and inertia = 9.5%.) Only the species with the highest contribution in the CA are indicated.
3.2 Changes in demographic stages and adult plant performance of A. nicaeensis after fire
For the unburned treatment, demographic stage densities did not differ among years except for the density of non-flowering individuals (Table 3). In contrast, demographic stage densities of the burned treatment differ significantly among years, and decrease between the two years for the densities of flowering individuals, seedlings, and clumps. Except for the non-flowering individual density, the demographic stages significantly differ between the two treatments in 2004, i.e. the first growing season after fire. Flowering individuals, seedlings, numbers of clumps and individuals per clumps are significantly higher in the burned treatment (Table 3). However, two years after fire, no significant difference is found between the unburned and burned treatments. The increase of seedlings observed the first year after fire (2004) is not followed by an increase in the number of individuals of A. nicaeensis the year after. Overall, the densities of demographic stages for the two treatments and years reveal an increase of seedling recruitment one year after fire for the burned treatment, but this recruitment strongly decreases the next year and is absent in the unburned treatment (Table 3). In addition, temporal trends are significantly different in the two treatments for flowering individuals, seedlings, and number of clumps (significant interaction).
Density of demographic stages, numbers of clumps and individuals per clump, total number of seeds per square meter and mean number of seeds per fruit for A. nicaeensis in the two treatments (TRT), one and two years after fire. Mean values ± S.E., and F-values (generalised linear models, ddl = 1, 19) associated with variations of demographic stage and in relation to treatment and year (; ; , not significant: ns). F-values and the significance from the variance analyses are given for the interaction () and the main effects ( and )
| Unburned | Burned | 2004 | 2005 | ||||||||
| between treatment | between treatment | ||||||||||
| 2004 | 2005 | F-value | 2004 | 2005 | F-value | F-value | F-value | ||||
| Number of individuals/m2 | |||||||||||
| Total | 3.9 (±1.9) | 3.9 (±2.8) | 12.9 (±2.5) | 4.7 (±1.9) | |||||||
| Flowering individuals | 8.63** | 18.92*** | 12.28** | 3.7 (±1.8) | 3 (±2.6) | 0.19 ns | 10.5 (±2.1) | 4 (±2.0) | 18.78*** | 23.04*** | 0.35 ns |
| Seedlings | 5.41* | 5.21* | 5.21* | 0 | 0 | 1.2 (±1.0) | 0.1 (±0.2) | 4.44*** | 5.49* | 1.00 ns | |
| Non-flowering individuals | 1.21 ns | 0.02 ns | 3.76 ns | 0.2 (±0.2) | 0.9 (±0.5) | 6.78* | 1.2 (±1.1) | 0.6 (±0.4) | 1.01 ns | 3.08 ns | 0.87 ns |
| Number of clumps | 7.28** | 10.51** | 12.84** | 0.6 (±0.6) | 0.7 (±0.6) | 0.04 ns | 2.7 (±0.9) | 0.7 (±0.4) | 16.22*** | 13.93*** | 0.11 ns |
| Number of individuals per clump | 6.55* | 0.83 ns | 3.45 ns | 2.3 (±0.6) | 2.5 (±0.6) | 0.29 ns | 2.8 (±0.8) | 1.9 (±1.2) | 1.36 ns | 14.88*** | 1.38 ns |
| Number of seeds per m2 | 1.91 ns | 3.00 ns | 3.94 ns | 50.5 (±30.7) | 55.8 (±75.7) | 0.02 ns | 130.9 (±35.9) | 53.1 (±38.1) | 8.47** | 11.09** | 0.01 ns |
| Number of seeds per fruit | 0.01 ns | 1.38 ns | 0.28 ns | 13.9 (±6.0) | 10.9 (±7.7) | 0.98 ns | 12.4 (±2.0) | 12.8 (±3.6) | 0.39 ns | 0.22 ns | 0.10 ns |
Proportion of mature fruits is higher in 2004 than in 2005 for the two treatments (Fig. 5). Comparisons between years for the two treatments also show an increase of the flower abortion in 2005. Comparisons among years of the number of mature fruits, flower abortion, and fruit predation analysed together showed neither significant differences (Table 4) nor temporal trends in the two treatments (no significant interaction). Comparisons among treatments showed significant differences between mature fruits, flower abortion, and fruit predation (RS significant). Also, neither temporal nor treatment trends were different for the reproductive phase success (no significant interactions), as well as interactions between year, treatment, and reproductive phase success (Table 4). In addition, no significant difference is observed between treatments and between years for the mean number of seeds per fruit (Table 3). In contrast, the total number of seeds produced is significantly higher in burned plots during the first growing season after fire, but it does not differ between treatments the second year (Table 3). The increase of seed number observed the first year after fire (2004) is not followed by an increase in recruitment the year after.

Proportion of mature fruits, flower abortion and fruit predation on the total of reproductive structure produced of A. nicaeensis for unburned and burned treatment one and two years after fire.
Log-linear model analysis of the effect of year, treatment (TRT) and reproductive phase success of A. nicaeensis (RS: proportion of mature fruits, flower abortion and fruit predation), and their interactions in each treatment (; ; not significant: ns)
| Source | df | P | |
| TRT | 1 | 7.01 | ** |
| Year | 1 | 3.13 | ns |
| Year ∗ TRT | 1 | 0.10 | ns |
| RS | 2 | 104.0 | *** |
| TRT ∗ RS | 2 | 2.03 | ns |
| Year ∗ RS | 2 | 4.78 | ns |
| TRT ∗ Year ∗ RS | 2 | 2.52 | ns |
3.3 Effect of post-fire substrate and vegetation on demographic stages
For A. nicaeensis, both the densities of total number of individuals and flowering individuals are significantly higher in plots where vegetation is lower and bare soil cover higher (Table 5). In contrast, no significant correlation exists with the cover of blocks, rocks, stones, and litter. Seedling density is negatively affected by vegetation height and cover (Table 5), and seedlings are more numerous in plots where bare soil cover is higher. Also, clump density and mean number of individuals per clump show a positive significant correlation with the bare soil cover.
Spearman rank order correlations for each demographic stage of A. nicaeensis and vegetation or substrate parameters (n=40). (; ; , not significant: ns)
| Variables | Total individual density | Flowering individual density | Seedling density | Clump density | Mean number of individual per clump |
| Vegetation height | −0.296 ns | −0.075 ns | |||
| Vegetation cover | −0.275 ns | −0.237 ns | −0.208 ns | −0.023 ns | |
| Bare soil cover |
4 Discussion
4.1 Effect of fire on vegetation and substrate cover
One growing season after fire, the unburned and burned plots are characterized by different substrate covers, and vegetation structures in response to fire. However, such differences were quickly attenuated two years after fire. Fire destroys initially leaf litter and vegetation, creating bare soil cover. This effect is particularly important in the Mediterranean ecosystems, where litter decomposition may be very slow due to the coriaceous leaf structure of many species, their high concentration in secondary compounds, and the low moisture content of soils [27]. Early in the first growing season after fire, vegetation height and cover are reduced compared to unburned treatment, as in other Mediterranean post-fire grasslands [28]. This drastic structural change increases the amount of light and water availability, and it decreases intra- and inter-specific competition (e.g., [10,29]).
Also, based on contrasting plant assemblages, we found floristic differences in response to fire. Unburned plots are dominated by a low stratum composed of perennial and xerophytic herbs, with a dense vegetation cover that characterizes these xerophytic grasslands. Such a habitat is colonized by small ligneous species and competitive herbs, because of the abandonment of the traditional agro-sylvo-pastoral systems in southern France [30,31].
In burned plots, we found a higher abundance and richness of herbs in response to fire, and an important change in species occurrence and diversity, a pattern classically observed (e.g., [28,29]). The cover and richness of annual herbs increase quickly after fire, as therophyty is an efficient strategy of temporary emergence in fire-prone ecosystems of the Mediterranean Basin [7]. In these communities, therophytes can establish the first year after fire in relation to a reduced competition, but they often disappear five years after wildfire at the latest [10]. Furthermore, fire causes a severe reduction of vegetation cover and height for small ligneous species. However, some of these species, which are top-killed by fire, resprout vigorously (e.g., Euphorbia spinosa) or have a high seedling emergence (e.g., Cistus albidus L., or Thymus vulgaris) [32,33]. Nevertheless, these immediate and profound ecological changes are considerably attenuated during the second growing season after fire because these Mediterranean habitats are particularly resilient and fire-adapted [6,10]. Thus, structural changes constitute only a short-term effect, but one that is of particular importance to the regeneration window of A. nicaeensis, and to other geophytes studied in several fire-prone ecosystems [12,13,19].
4.2 Effects of fire on demographic stages and adult plant performance
A. nicaeensis responds positively to this small fire event, with a significant increase of flowering individuals and seedlings after fire. Several studies in other Mediterranean ecoregions have demonstrated a similar response of geophytes to fire, suggesting that fire increases the short-term availability of plant resources, particularly light and nutrients, and reduced competition with the surrounding vegetation [9,34]. In the Cape Floristic Region, Le Maitre and Brown [12], and Verboom et al. [35] have underlined the positive consequences of fire on the stimulation of geophyte flowering, with direct ecophysiological effects of heat or smoke on seed germination, or through indirect environmental changes. As the density of flowering individuals was positively correlated with a lower vegetation height and a higher bare soil cover in response to fire, our results suggest that flowering is not strictly fire-dependent for this geophyte. In addition, the lack of difference in demographic stage densities and reproductive success during the second growing season after fire between unburned and burned situations highlights the short-term benefit of fire for the dynamic of A. nicaeensis.
One year after fire, A. nicaeensis from burned plots shows a high proportion of seedlings (9.3%), in contrast to unburned plots, where no seedling was observed. Nevertheless, most of the new plantlets disappear the year after, and they probably died or were in dormancy as a bulb bank. The high seedling densities in burned treatment could be explained by the presence of an important seed bank (pers. obs.), higher seed germination, and/or a reduced competition with the surrounding vegetation. Even if the presence of seedlings is very scarce in this xerophytic grassland under unburned situation [22], the germination experiment in laboratory conditions of seeds from A. nicaeensis that occurred in unburned treatment indicates a high germination rate of 88% [20]. Thus, the higher seedling density observed one year after burning could be mostly explained by the indirect effect of fire, which generally reduces aboveground competition and determines a proper regeneration niche sensu Grubb [36] through increase of resource availability. Fire favours light penetration into the soil [37], and a greater fluctuation in soil temperatures with increased oxygen concentrations after the fires may stimulate the germination of several life forms [9]. In general, these deep environmental changes have an influence on seedling emergence higher than the direct influence of fire on seed germination [38,39].
Proportions of mature fruits, flower abortion, and fruit predation do not differ between the two treatments and years, as well as the mean number of seeds per fruit, in contrast with other studies that mention a reduced fruit-set in burned treatment as a result of low activity of pollinators [40], although the difference of proportions of mature fruits and flower abortion of plots between the two years could be explained by the contrasted activity of pollinator linked to various climatic conditions. Higher density of clumps and number of individuals per clump in burned treatment are observed one year after fire, indicating a direct and/or indirect effect of fire on asexual reproduction. In addition, the higher total number of seeds produced in the burned plots one year after fire (ca. 131 seeds/m2 vs. 50–55 seeds/m2 in unburned situations) indicates an increase of the sexual reproduction in response to fire, even if the mean number of seeds per fruit remains unchanged between treatments and years.
In the northern part of the Mediterranean Basin, land-use changes favour the fast colonization of oligotrophic habitats by several competitive species that can modify the vegetative production mechanism and the sexual reproduction of several rare Mediterranean herbs [41], due to the considerable shading they cause, whether directly or indirectly through litter [42]. Nevertheless, we could not exclude the combination of direct and indirect effects of fire on the asexual and sexual reproductive mechanisms of this geophyte, since such combined effects were already demonstrated for Cape geophytes [12].
For A. nicaeensis, the number of flowering individuals and clumps, and the mean number of individuals per clumps increase together in the same way. In contrast with other species (e.g., [43]), sexual reproduction of A. nicaeensis seems not to occur at the expense of vegetative survival and growth. Thus, our data suggest a lack of trade-off between vegetative and sexual function [35]. Such a pattern could explain why a high density of flowering individuals is linked to conditions favouring high productivity such as environments induced by fire, as it has already been demonstrated in clonal geophytes [12].
4.3 Importance of fire for Mediterranean geophyte conservation
Despite the frequency of fires in the Mediterranean Basin, their effects on viability and managing of populations of rare and endangered Mediterranean plants are almost totally unknown. A neglected aspect concerning rare Mediterranean geophytes is notably related to the magnitude of the storage effect sensu Higgins et al. [44], i.e. the storage of reproductive potential over generations, which allows bridging unfavourable recruitment periods. Storage organs such as bulbs can indeed buffer the negative effects of environmental variability (climatic events and disturbances) and enhance population persistence over time for the plant species living in such stochastic Mediterranean environments [16]. Fire response traits of this palaeoendemic species, i.e. the resprouting capacity and the propagule-persistence capacity provide also an important trade-off association to persist facing this natural disturbance present since the Tertiary [11].
As for A. nicaeensis, demographic stage densities and adult plant performance of other rare geophytes such as Acis longifolia (J. Gay ex Salisb.) J. Gay ex M.J. Roem. in Corsica [45], or Simethis mattiazzii (Vand.) G. Lòpez & Jarvis in siliceous Provence seem to be influenced positively after moderate fire. Infrequent and reduced fires could provide an adequate interval for the regeneration window of these heliophilous plants (see also [13,46]). In this context, fire could be an important factor in regulating the demographic processes of small populations of rare and threatened geophytes, as indicated by the intermediate disturbance hypothesis [47]. Nevertheless, the use of prescribing fires implies to consider all disturbance parameters, including intensity, severity, frequency and season of fire [4], as well as the nature of the plant communities involved [33].
The enhanced recruitment of A. nicaeensis is probably due to the indirect impact of fire on xerophytic grassland. In this fire-prone habitat, fire severity is mitigated by the presence of species with high capacity for regeneration, such as Cistus albidus, Euphorbia spinosa or Thymus vulgaris [33]. For A. nicaeensis, one of the major causes for population decline is indeed the colonization by competitive species in its microhabitats induced by land-use changes, which lead to a higher probability of extinction linked to the lack of recruitment [48]. Thus, micro-site and vegetation characteristics seem to play a determinant role in the persistence of A. nicaeensis, in particular in driving the seedling density as well as vegetative production mechanisms and sexual reproduction. In this way, our results suggest that reduced and prescribed fires could represent an adequate conservation option to the persistence of rare and threatened geophytes facing the actual drastic land-abandonment in the northern Mediterranean. Future studies should examine the optimal fire return interval for the long-term persistence of these endangered plants [46].
Acknowledgements
This research was funded by the Conservatoire botanique national méditerranéen de Porquerolles.