

1 Introduction
Les maladies du bois sont considérées comme très dommageables pour la pérennité du patrimoine viticole. Les parasites responsables de ces maladies provoquent à plus ou moins long terme la mort du cep et peuvent nécessiter un renouvellement des plants pouvant atteindre plus de 10% d'un vignoble [1]. Depuis environ une dizaine d'années on constate une progression inquiétante de ces maladies. Plusieurs observations laissent à penser que ce phénomène n'est probablement qu'en début de cycle : (i) l'interdiction de l'arsénite de soude, seul moyen actuellement connu pour combattre l'esca et le Black Dead Arm (BDA) ou « bras mort noir », à partir de 2001 [2], (ii) l'augmentation annuelle du taux de mortalité de 4 à 5% à partir de la cinquième année dans des parcelles où le traitement par l'arsénite de soude a été arrêté et (iii) le taux élevé de ceps asymptomatiques contaminés dans le vignoble [2].
La suppression de l'arsénite de soude dans tous les pays viticoles du fait de sa toxicité non seulement pour l'environnement mais aussi pour l'homme [3], inquiète d'autant plus les viticulteurs qu'aucune méthode de lutte de remplacement satisfaisante ne leur a été proposée. Ceci met en péril le maintien de l'outil de production et sa longévité et cela à l'échelle mondiale. Par conséquent, les effets de ces maladies du bois seront nombreux à moyen terme : elles entraîneront soit une dépréciation de la qualité des vins suite à un rajeunissement des parcelles, soit une perte de la typicité d'un vin d'une région viticole suite à la non replantation des cépages les plus sensibles.
Le vignoble français regroupe trois principales maladies du bois : l'eutypiose, l'esca et le BDA. Ces maladies de dépérissement sont associées à la présence de différents champignons capables de dégrader les tissus ligneux.
Phaeomoniella chlamydospora, Phaeocremonium aleophilum, Eutypa lata, Fomitiporia mediterranea, Botryosphaeria obtusa, Neofusicoccum parvum et Botryosphaeria stevensii sont les principales espèces isolées en France associées au dépérissement de la vigne.
Ces maladies s'expriment au niveau du bois par la formation de nécroses sectorielles et/ou centrales, par la présence de bandes brunes ou de chancres dans le tronc et les bras et au niveau foliaire par des décolorations et des dessèchements qui peuvent être foudroyants.
La lutte contre les maladies du bois est actuellement engagée. Des études épidémiologiques et des travaux sur l'incidence des pratiques culturales ont débuté. Afin d'améliorer la prophylaxie et la production de matériel sain, des méthodes de diagnostic précoce des maladies sont élaborées actuellement. Des molécules chimiques ou biologiques pouvant substituer l'arsénite de soude sont criblées in vitro et en champs.
Il nous paraît donc important de faire le point sur les connaissances des maladies du bois chez la vigne (Vitis vinifera L.) et plus particulièrement concernant l'esca et le Black Dead Arm, principales maladies impliquées dans ce nouveau désastre viticole [2].
2 Deux principales maladies impliquées dans les maladies du bois
2.1 Esca
2.1.1 Symptomatologie
2.1.1.1 Historique de la maladie.
L'esca, maladie cryptogamique, décrite sous le nom de pourriture à l'avènement de notre ère est plus particulièrement présente dans les vignobles de l'hémisphère Nord. Elle a été décrite tout d'abord sous sa forme apoplectique [4], dont l'origine fut longtemps indéterminée et attribuée à un accident d'ordre physiologique qui était désigné sous le nom de folletage [5,6]. Ce n'est qu'à la fin du XIXème siècle que cette forme fut attribuée à un champignon et de ce fait distinguée du folletage [7]. En 1922, le terme esca a été introduit pour désigner cette maladie caractérisée par la présence d'une pourriture blanche et par la forme apoplectique [8]. L'esca est un terme d'origine languedocienne ou provençale qui signifie amadou (= pourriture blanche). Il était désigné auparavant sous le nom de iska en Grèce ou de yesca en Italie. Entre 1923 et 1931, d'autres formes d'expression ont été décrites sous le nom d'apoplexie lente [9–11]. La première, qui est connue actuellement sous le nom de forme lente de l'esca, est caractérisée par des digitations jaunes (cépages blancs) ou rouges, bordées de jaune (cépages noirs) entre les nervures qui restent vertes. La deuxième se caractérise également par la présence de digitations inter-nervaires, mais se distingue de la forme lente de l'esca par l'absence de liseré jaune dans les premiers stades de l'apparition des symptômes (figure 1). Cette forme lente est caractéristique d'une autre maladie connue sous le nom de forme lente du Black Dead Arm [12]. Ces auteurs décrivent également un autre aspect des symptômes qui est caractérisé par un rabougrissement des rameaux, symptôme caractéristique d'une autre maladie de la vigne, l'eutypiose [13].

Symptômes foliaires associés aux différentes maladies : dépérissement foudroyant (A), symptôme sur cépage blanc (B), symptômes sur cépage noir (C) et coupes transversales d'un tronc présentant des nécroses typiques des maladies du bois (D).
Leaf symptoms linked to the different diseases: apoplectic decline (A), symptom on white vine (B), symptom on red vine (C) and transversal section of a trunk presenting typical grapevine trunk disease necrosis.
Les symptômes sur fruits se traduisent soit par un retard dans leur maturation, soit par leur flétrissement [14]. En Californie [15] et en Italie [16,17], les fruits peuvent présenter des taches brunes violacées à leur surface. Ces symptômes sont en association ou non avec les symptômes foliaires [15].
La forme lente de l'esca est associée à différentes nécroses dans le bois [18] (figure 1). Elle peut être reliée à une pourriture blanche [18], à la présence d'une nécrose brune en position centrale [18] ou à des ponctuations noires (correspondant à des vaisseaux nécrosés) lorsque la maladie n'est pas très évoluée ou avancée [19].
2.1.1.2 Comportement des cépages.
Les enquêtes réalisées dans le vignoble ont permis de classer les cépages selon la sensibilité à l'expression des symptômes foliaires [1]. Par exemple, les cépages Cabernet Sauvignon, Sauvignon blanc, Mourvèdre, Ugni blanc, Cinsault, Trousseau, expriment plus facilement les symptômes de la forme lente de l'esca que le Merlot, le Carignan, le Roussane, le Pinot noir. Il est à noter qu'aucune étude n'a été réalisée pour classer les cépages selon l'importance des nécroses dans le bois. Aucune donnée n'est également disponible sur les espèces autres que Vitis.
Il est également important de signaler que les vignes-mères de porte-greffe ne présentent pas de symptômes foliaires sur la partie herbacée, seules des nécroses dans le bois caractéristiques de l'esca y sont observées.
2.1.1.3 Facteurs environnementaux.
Peu d'informations sont disponibles sur les facteurs environnementaux ayant un rôle majeur dans le développement de la maladie et de son expression.
La manifestation des symptômes foliaires est fluctuante d'une année sur l'autre [20,21]. Les étés doux et pluvieux sont plus favorables à l'expression des symptômes de la forme lente alors que des étés plutôt chauds sont plus favorables à l'apoplexie [21]. Une étude des bilans hydriques réalisée pendant trois années consécutives (2004 à 2006) dans le vignoble bordelais montre que les stress hydriques de la vigne contribueraient à inhiber l'expression foliaire [22]. Ces auteurs montrent également que les parcelles à forte réserve utile, dont le sol est caractérisé par une alimentation en eau non limitante, seraient plus atteintes par les maladies du bois.
La manifestation de l'apoplexie est observée selon certaines conditions climatiques, notamment lors de périodes de sécheresse ou de vent fort. Elle est dûe au fait que l'évapotranspiration trop importante durant ces périodes n'est plus compensée par un afflux en eau suffisant car il ne reste qu'une faible quantité de bois fonctionnel.
Certaines connaissances sont également disponibles sur la comparaison des incidences entre les cépages dans différentes régions [23].
Parmi les facteurs biotiques (interventions humaines), il est actuellement difficile de déterminer leur influence sur l'esca.
2.1.2 Champignons associés
2.1.2.1 Historique.
Cette maladie fut initialement attribuée à un seul champignon à la fin du XIXème siècle [7]. La présence de carpophores de Stereum hirsutum (Willd. : Fr.) S.F. Gray et de Phellinus igniarius (L. : Fr.) Pat. sur les ceps dépérissants suggérait au début du XXe siècle, que ces champignons en étaient responsables [24–26]. Rives [26] concevait d'ailleurs que suivant les régions, de telles espèces de champignon lignicole prédominaient plutôt qu'une autre. En 1959, Chiarappa [15] soulignait l'importance d'un autre champignon dans la maladie, Cephalosporium sp., qui, par la suite, a été identifié comme étant Phaeomoniella chlamydospora (W. Gams, Crous, M.J. Wingf. & L. Mugnaï) Crous et W. Gams [27] ; il considérait également S. hirsutum comme un agent saprophyte et mettait en évidence le caractère parasitaire de P. igniarius. A la fin du XXe siècle, l'esca était donc plutôt attribué à un complexe de champignons qui, par leur succession dans le bois conduisait à la formation de la nécrose qui lui est caractéristique [28].
2.1.2.2 Processus menant à la dégradation du bois.
Deux processus menant à la formation de cette nécrose ont été décrits [18,28]. Le premier processus aboutit à la formation d'une nécrose claire et tendre en position centrale et fait intervenir trois champignons : Phaeoacremonium aleophilum (W. Gams, Crous, M.J. Wingf. & L. Mugnaï et P. chlamydospora Crous et W. Gams, gen. nov.), trouvés dans une nécrose brune et dure en position centrale, et Fomitiporia mediterranea Fischer (= Fomitiporia punctata (Fr. ex Karst.) Murrill = Phellinus punctatus (P. Karst.) Pilát), responsable de la pourriture blanche [18,28–30]. Le deuxième processus conduit à la formation d'une nécrose claire et tendre en position sectorielle et implique Eutypa lata (Pers : Fr.) Tul & C. Tul, responsable de la nécrose brune en position sectorielle, et F. mediterranea qui dégrade ce bois brun en une pourriture blanche [18]. Les deux processus peuvent être observés sur la même plante [18].
F. mediterranea est le champignon le plus trouvé dans la pourriture blanche en France [18], en Italie [29], en Espagne [31] et en Allemagne [32]. D'autres champignons sont responsables de la pourriture blanche tel que S. hirsutum dont la présence dans les ceps atteints d'esca est décrite en Hongrie [33], en France [18] et en Autriche [34]. Ces champignons ne semblent pas être présents sur les autres continents. Les microrganismes responsables de la pourriture blanche sur les autres continents sont différents de ceux trouvés en Europe. Fischer [35] signale la présence de Fomitiporia polymorpha en Amérique du Nord, Fomitiporia australiensis et de deux taxons inconnus en Australie, trois taxons inconnus en Afrique du Sud, Inocutis jamaicensis et Fomitiporella vitis en Amérique du Sud, associés respectivement à la Hojà de Malvon [36] et à l'enroulement chlorotique des feuilles [37] (tableau 1).
Implantation géographique des différents champignons impliqués dans les maladies du bois chez la vigne.
Geographical establishment of the different fungi implied in the grapevine trunk diseases.
| Champignons | Présence dans les vignobles |
| Phaeomoniella chlamydospora | Afrique du Sud [38], Algérie [39], Allemagne [32], Australie [19], Espagne [31], Etats-Unis [42], France [18], Grèce [40] , Iran [41], Italie [21], Turquie [43], Autriche [34]… |
| Phaeoacremonium aleophilum | Algérie [39], Allemagne [32], Espagne [31] Etats-Unis [42], France [18], Grèce [40] , Italie [21], Turquie [43]… |
| Eutypa lata | Europe – Amérique du Nord – Brésil – Afrique du Sud – Nouvelle-Zélande – Australie [105] |
| Fomitiporia mediterranea | Allemagne [32], Espagne [31], France [18] Italie [29] |
| Stereum hirsutum | Autriche [34], Espagne [31], France [18], Hongrie [33], Italie [29] |
| Botryosphaeria obtusa | Chili [44], Espagne [141], France [12], Italie [45,46], Liban [47] … |
2.1.2.3 Tests de pathogénie.
Des expérimentations sur la pathogénie ont montré que les champignons sont capables de provoquer les symptômes dans le bois. P. chlamydospora, inoculé dans des boutures, provoque des ponctuations noires, identiques à celles observées dans le vignoble [18,48,49]. Suite à des inoculations artificielles de boutures, P. aleophilum peut provoquer des nécroses sectorielles dans le bois [18].
Concernant F. mediterranea, les tests réalisés sur du bois sain, prélevé sur le tronc de ceps montrent qu'il dégrade le bois en une pourriture blanche, nécrose caractéristique de l'esca. Il en est de même pour S. hirsutum, ce champignon peut également provoquer, contrairement à F. mediterranea, de telles nécroses lorsqu'il est inoculé dans les tiges ligneuses de boutures [18]. Aucune expérimentation n'a été réalisée jusqu'à aujourd'hui pour valider les différents processus menant à la dégradation du bois caractéristique de l'esca.
Peu de travaux expérimentaux sont disponibles sur les champignons qui pourraient être impliqués dans l'expression des symptômes foliaires (forme lente). Sparapano et al. [50] a reproduit des symptômes foliaires similaires à ceux de l'esca en inoculant les différents champignons seuls ou en association. Feliciano et al. [51] ont reproduit dans le vignoble les symptômes de la forme lente de l'esca deux ans après l'inoculation de P. aleophilum ou de P. chlamydospora sur Thompson Seedless. Cependant, ces expérimentations n'ont pas rempli pleinement le postulat de Koch.
Concernant les symptômes sur fruits (taches violacées en surface), Sparapano et al. [50] et Gubler et al. [52] montrent le caractère pathogène de P. chlamydospora et de P. aleophilum sur les baies de raisin blessées de manière artificielle.
2.1.2.4 Caractéristiques et cycles biologiques des champignons.
Phaeomoniella chlamydospora
W. Gams, Crous, M. J. Wingf. & L. Mugnaï Crous et W. Gams
Ce champignon est présent dans la majorité des vignobles (tableau 1). Outre l'esca, ce champignon est associé à la maladie de Petri, affection des jeunes plantations [53] qui est signalée en Afrique du Sud, Europe, Etats-Unis et en Australie. Il est également impliqué dans un dépérissement chez Actinidia sinensis [54]. Il appartient à la Classe des Ascomycètes de la famille des Herpotrichiaceae [55]. Sa forme sexuée n'est pas connue. La description de ce champignon est signalée par Crous et al. [56] et Crous et Gams [27]. Ses conditions de culture (température, pH) sont décrites par Whiting et al. [57] et Valtaud et al. [58]. P. chlamydospora a été classé parmi les champignons de bleuissement car il ne dégrade pas les parois cellulaires [28]. Les enzymes produites et identifiées sont des polygalacturonases, des polyméthylgalacturonases [59], des β-glucosidases et des endo-β 1,4 glucanases. Des polysaccharides extracellulaires ont été également identifiés [60] ainsi que neuf autres métabolites dont certains sont considérés comme étant des toxines potentielles (p-hydroxybenzaldéhyde) [61]. P. chlamydospora inhibe la formation du cal [62], provoque une forte mortalité des greffés-soudés s'il est inoculé au niveau de la soudure [63] et diminue la vigueur des plants [64]. Aucun élément n'est apporté au sujet des déterminants du pouvoir pathogène.
P. chlamydospora est un champignon à dissémination aérienne qui pénètre par les plaies de taille pendant la période hivernale lors de périodes douces et pluvieuses [65–67] jusqu'au moment des pleurs [65] et après la période des pleurs. Sa dissémination peut également se dérouler pendant la période végétative de la vigne [65]. Les différentes plaies réalisées lors des opérations en vert (ébourgeonnage, effeuillage, échardage, rognage, éclaircissage) peuvent constituer des voies potentielles de sa pénétration dans la plante comme l'ont montré les tests d'inoculation. La source d'inoculum de P. chlamydospora sous forme de pycnides est localisée sur les plaies de taille âgées [67,68], dans des zones protégées (craquelures dans le bois) [69] ou encore sous l'écorce dans les zones où les vaisseaux sont exposés suite à des blessures [68]. Les pycnides contiennent des pycniospores dont le pouvoir germinatif a été montré par Rooney et Gubler [70]. Une abondante sporulation peut être également observée au niveau des craquelures et des crevasses qui fournissent un environnement humide favorable à l'émergence du champignon [69]. Sa présence a également été montrée à la surface de ses différents organes suggérant son caractère épiphyte [71], mais actuellement aucune information ne permet de dire si, ce sont les surfaces qui sont polluées lors de sa dissémination ou alors s'il s'agit du champignon qui se développe à la surface des organes. Le champignon peut également se propager par les bois de porte-greffes ou de greffons en pépinières [63,65,72–74]. Son origine dans les tissus ligneux des sarments n'est pas entièrement connue. Trois hypothèses peuvent expliquer cela : soit le mycélium présent dans la souche va progresser dans les rameaux, soit les spores sont transportées par la sève jusqu'au niveau des rameaux, ou encore les rameaux ou les sarments peuvent être contaminés. Le champignon a été mis en évidence dans les pieds-mères de bois de porte-greffe [75]. Des conidies ont pu être observées dans les vaisseaux de jeunes plantes inoculées artificiellement. Whiteman et al. [76] supposent que chez les vignes-mères de porte-greffes infectées, les spores sont transportées par la sève à différents niveaux dans le sarment. Cependant, des analyses microbiologiques réalisées sur toute la longueur de sarments de porte-greffe montrent que P. chlamydospora n'est isolé que dans la partie proche de la tête de saule [63]. Sa présence sur la surface du matériel de propagation (greffons et porte-greffe) a été montrée au moyen des techniques PCR [77] et pourrait ainsi constituer une source d'inoculum non négligeable pour les contaminations observées en pépinières. En pépinières, des analyses de détection par PCR ont montré sa présence dans les bains de réhydratation [74,75,78,79], sur les outils de greffage [75,78,79], dans le substrat lors de la stratification en pot [78], dans l'eau [80] ou encore dans la sciure [74]. Le champignon est également trouvé dans les greffes-boutures après la stratification [81]. Une des phases au cours desquelles ont lieu les contaminations lors de l'élaboration des plants a été identifiée, il s'agit de la stratification. Le champignon pénètre préférentiellement dans les greffes-boutures par les plaies situées à la base du plant (talon, plaies d'éborgnage) pour les plants ayant subi la stratification à l'eau [63] ou par celles localisées dans sa partie haute pour les greffes-boutures ayant subi la stratification à la sciure [82]. Avant la commercialisation des plants, le champignon peut être non seulement présent dans les tissus ligneux [83–85], mais aussi en surface. De nombreux travaux ont également porté sur la recherche du champignon dans le sol. Aucune étude réalisée n'a permis de le détecter par des techniques classiques [86]. Des techniques moléculaires l'ont détecté dans le sol au niveau des vignes-mères de porte-greffes. [74,87]. Gubler et al. [88] montrent sa capacité à survivre plusieurs mois dans le sol (tests de laboratoire) et Fourié et Halleen [73] l'ont trouvé dans les racines. Cependant, aucune relation n'a été montrée entre sa présence dans le sol et la contamination de la plante. Très peu d'études ont été effectuées sur sa propagation. Les études sur sa variabilité génétique montrent que sa population ne présente pas de biodiversité génétique importante [89–91]. Aucune information n'est apportée sur la distance à laquelle le champignon peut se disséminer à partir de la source d'inoculum ou encore s'il existe des pratiques culturales qui peuvent favoriser sa propagation.
W. Gams, Crous, M.J. Wingf. & L. Mugnaï
Il est présent dans la majorité des vignobles (tableau 1). Sa description a été réalisée par Crous et al. [56] et Mostert et al. [92] et ses conditions de culture par Whiting et al. [57] et Valtaud et al. [58]. Il appartient à la Classe des Ascomycètes et à l'Ordre des Calosphaeriales [92]. La forme sexuée a été obtenue in vitro par le croisement de souches compatibles sur milieu de culture [92] ou par incubation en conditions humides de souches atteintes d'esca [93]. Des périthèces du genre Togninia sont obtenus au bout de 3–4 semaines d'incubation et la comparaison avec la morphologie et les séquences ITS publiées pour d'autres espèces de Togninia démontre que la forme teléomorphe de P. aleophilum est Togninia minima (Tul. & C. Tul) Berl [93,94]. Cette forme a été décrite pour la première fois en 1992 par Hausner et al. [95] sur différents hôtes [Aulne, Saule, Prunus pennsylvanica], puis a été observée dans le vignoble californien [96]. Ce champignon est aussi rencontré sur d'autres plantes-hôtes notamment l'Actinidia sinensis [54] et l'olivier [97]. Ce champignon a été classé parmi les champignons de la pourriture molle car il se développe à l'intérieur des parois secondaires en y provoquant la formation de cavités [28]. Les enzymes produites et identifiées sont des xylanases, des β-glucosidases et des endo-β 1,4 glucanases [28] ainsi que sept autres métabolites identifiés comme étant des toxines potentielles (naphthalenones) [61,98,99]. Contrairement à P. chlamydospora, P. aleophilum n'inhibe pas la formation du cal au niveau de la soudure greffon/porte-greffe [62]. Les déterminants du pouvoir pathogène n'ont pas été identifiés mais son rôle important dans la progression des nécroses dans le bois a été montré [28].
P. aleophilum est un champignon à dissémination aérienne [65] qui contamine les plaies de taille plus particulièrement pendant la période végétative. Il est également capable de contaminer les plaies réalisées lors des opérations en vert comme le montrent les tests d'inoculation et ses spores sont surtout disséminées pendant la période végétative de la vigne [65,67]. La source d'inoculum (sous forme mycélienne) est située sur le tronc ou les bras excoriés [65] et les périthèces sont situés dans des craquelures ou des crevasses du cep [96]. Ce champignon peut également se propager par les bois de porte-greffes ou de greffons en pépinières [63,65,81], mais son origine dans les sarments n'est pas connue. La présence de conidies a été observée dans les vaisseaux [100] et des analyses microbiologiques réalisées sur toute la longueur de sarments de porte-greffes montrent qu'il est trouvé uniquement dans la partie proche de la tête de saule [63]. Sa présence sur la surface du matériel de propagation (greffons et porte-greffes) a été montrée au moyen des techniques PCR [77]. Une des phases au cours desquelles ont lieu les contaminations lors de l'élaboration des plants a été identifiée, il s'agit de l'élevage des plants au champ où le champignon infecte les plants uniquement par la partie aérienne [63]. L'origine des contaminations au champ n'est pas connue, deux hypothèses peuvent être faites : soit l'inoculum provient des plants présents en surface, soit du milieu environnant (autres vignes, autres plantes hôtes). Avant la commercialisation des plants, le champignon est non seulement présent dans les tissus ligneux [84,85], mais aussi en surface. P. aleophilum a été trouvé dans le sol au moyen de méthodes utilisant des milieux sélectifs [86] ou par PCR [101]. Mais aucune relation n'a été montrée entre sa présence dans le sol et l'infection des plantes. Les études sur la variabilité génétique de ce champignon montrent que la diversité génétique au sein d'une population est peu importante. Plusieurs haplotypes différents de P. aleophilum sont trouvés au sein d'un vignoble, suggérant plusieurs sources d'inoculum primaire [89–91].
[Pers : Fr.] Tul & C. Tul
Il s'agit du champignon responsable à lui seul de l'Eutypiose [102] et pouvant être associé à l'esca [18]. Il appartient à la Classe des Ascomycètes, à l'Ordre des Diatrypales et à la famille des Diatrypacées [103]. Ses conditions de culture sont décrites par Amborabé et al. [104]. Ce champignon est trouvé sur plus de 80 plantes hôtes [105] et est présent dans les vignobles où le niveau de pluviométrie est supérieure à 250 mm par an [102]. E. lata a été classé parmi les champignons de la pourriture molle car il se développe à l'intérieur des parois secondaires en y provoquant la formation de cavités [28]. Les enzymes produites et identifiées sont des xylanases, des β-glucosidases, des cellulases, des β-1,3 glucanases, des chitinases, des protéases [106], des laccases, des polygalacturonases, ainsi que des pectine-méthyl-estérases. D'autres métabolites toxiques (T) ou non (NT) sont produits par le champignon [107–109] : eutypine (T), eulatinol (T), eulatachromène (T), benzofurane (T), siccayne (NT), eutypinol (NT). Le mode d'action de l'eutypine [110–116] ainsi que des autres métabolites secondaires [116] a été étudié. Il a été mis en évidence que E. lata secrète d'autres métabolites de nature polypeptidique qui induisent des effets toxiques sur les rameaux et les feuilles des vignes [117,118]. Enfin, il a également été montré qu' E. lata synthétise de l'acide abscissique et des polyamines (putrescine, spermidine et spermine) [119]. Les déterminants du pouvoir pathogène sont aussi inconnus, toutefois, Koussa et al. [119] constatent une relation possible entre l'agressivité de deux souches d'E. lata et leur contenu en polyamines et en acide abscissique.
Son cycle biologique a été décrit par Carter [102]. Il s'agit d'un champignon à dissémination aérienne qui se propage toute l'année [120]. Il pénètre par les plaies de taille suite aux précipitations pendant la période hivernale [120,121]. Le champignon peut migrer jusqu'à 2 cm dans les tissus sous-jacents à la plaie de taille [122]. Lors de la période végétative, d'autres voies de contamination, bien que mineures par rapport aux plaies de taille, sont possibles comme les plaies d'ébourgeonnage et d'épamprage [123]. Les études de variabilité génétique suggèrent qu'il se propage uniquement sous sa forme sexuée [124] et se conserve sous forme de périthèces sur les bois. Il ne se propage pas par le matériel végétal de propagation [77].
F. mediterranea M. Fischer (désigné auparavant sous le nom de Phellinus punctatus [P. Karst.] Pilát, et de Fomitiporia punctata [Fr. ex Karst.] Murrill) est présent en Europe [125] (tableau 1). Il appartient à la classe des Basidiomycètes et à l'ordre des Hyménochaetales. Ses conditions de culture et sa description sont données par Fischer [125]. Il est rencontré sur différentes plantes : Vitis vinifera, Corylus avellinus, Olea europaea, Lagerstroemia indica, Actinidia sinensis, Acer negundo. Ce champignon a été classé parmi les champignons de la pourriture blanche car il dégrade complètement les parois cellulaires [28]. Les enzymes produites et identifiées sont des xylanases, des cellulases, des β-1,3 glucanases, des laccases, des péroxydases et des phénoloxydases. D'autres métabolites de type p-hydroxybenzaldéhyde sont produits par le champignon [126].
Peu d'études ont été réalisées sur son cycle biologique. Il se conserve sous forme de basidiomes sur les parties malades de la plante, et se dissémine lorsque les températures sont supérieures à 10 °C et le taux d'hygrométrie est supérieur à 80% [127]. La présence du champignon sur les plaies de taille récentes a été montrée par des outils moléculaires (PCR gigogne). Les études sur sa variabilité génétique [128,129] suggèrent fortement qu'il se propage par les basidiospores dans le vignoble. Les travaux effectués en pépinière n'ont pas permis de détecter sa présence dans le matériel de propagation, ni dans les plants avant leur commercialisation [77].
2.1.3 Interactions hôte-pathogène
2.1.3.1 Impact sur le métabolisme carboné.
Depuis l'interdiction de l'arsénite de sodium, seul produit chimique considéré comme efficace dans la lutte contre l'esca, de nouvelles stratégies de lutte doivent être développées. Ces dernières pourront être optimisées lorsque nous aurons une idée de l'impact de la maladie sur la physiologie de la plante.
Ainsi, depuis quelques années, des études s'intéressent à l'évolution de la physiologie de la plante au cours du développement de la maladie et plus particulièrement sur le métabolisme carboné et le statut hydrique de la plante. L'ensemble des travaux sur le métabolisme carboné a été mené vis-à-vis de la forme lente de l'esca. Petit et al. [130] ont observé une altération de l'appareil photosynthétique associée à l'expression des symptômes foliaires, qui se traduit par une diminution de l'activité photosynthétique et de la transpiration, une perturbation du fonctionnement des photosystèmes et une diminution des concentrations en pigments chlorophylliens. De plus, Petit et al. [130] ont noté un gradient de la perturbation de l'activité photosynthétique à l'échelle de la plante. En effet, les feuilles présentes sur un rameau sans symptômes montrent une photosynthèse peu diminuée alors que celles présentes sur un rameau malade ont une photosynthèse fortement altérée (figure 2). En complément de ces travaux, Christen et al. [131] ont observé que cette modification de la photosynthèse pouvait être détectée 2 mois avant l'expression des symptômes foliaires, chez le Cabernet-Sauvignon. La conséquence directe de cette diminution de la photosynthèse est une chute des réserves carbonées dans les sarments [130]. L'année suivante, les faibles réserves peuvent entraîner une baisse de vigueur de la plante, affectant son développement végétatif et reproducteur. L'ensemble de ces travaux sur le métabolisme carboné a été mené vis-à-vis de la forme lente de l'esca.
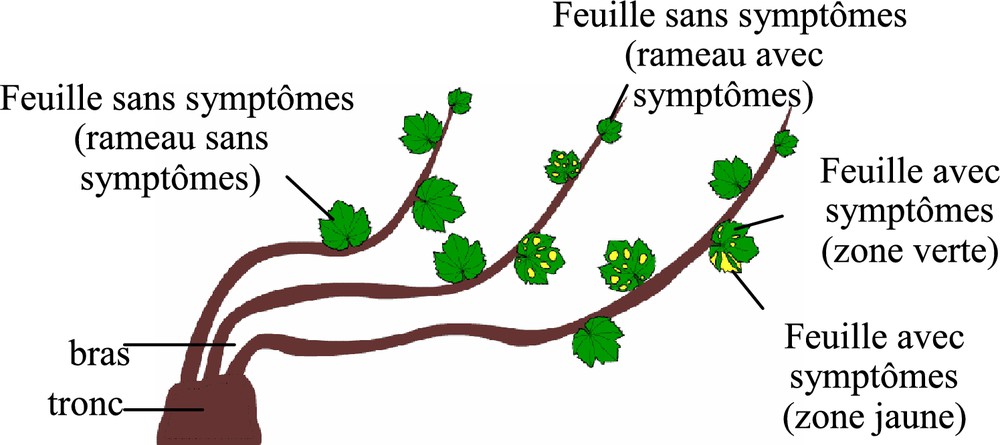
Schéma d'un pied potentiellement infecté par le complexe des maladies du bois.
Diagram of a trunk potentially infected by the wood disease complex.
A l'inverse, la forme foudroyante étant souvent associée à un excès d'eau dans le sol avec des températures élevées [132], l'altération du transport hydrique de la plante liée à des dysfonctionnements du xylème a été étudiée. Les outils disponibles pour ces études sont des mesures de fluorescence chlorophyllienne [131], l'imagerie thermique [133,134] ou la conductance stomatique [135]. Ainsi, Edwards et al. [135] ont observé une diminution de la conductance stomatique entraînant un déficit hydrique chez des plantes infectées par P. chlamydospora. Une étude comparative de la fluorescence chlorophyllienne d'une plante exprimant la maladie (forme lente ou forme foudroyante non précisée) avec celle subissant un stress hydrique a révélé deux fonctionnements différents du photosystème II [131]. Il a donc été conclu que l'expression des symptômes de l'esca ne pouvait pas simplement être corrélée à un stress hydrique, mais qu'elle était plus complexe.
2.1.3.2 Esca, toxine et expression des symptômes.
L'expression des symptômes au niveau des feuilles et des baies n'étant pas reliée à la présence de mycélium des champignons dans le rameau, l'hypothèse qu'ils produisent des molécules agissant à distance a donc été émise [136]. Ainsi, certains champignons produiraient des molécules qui seraient transportées jusqu'aux feuilles et aux baies via la transpiration. Les champignons associés à l'esca produisent en effet une variété de molécules dont certaines ont été identifées. Tabacchi et al. [61] ont décrit de nombreuses molécules chez P. chlamydospora dont l'isosclérone et la scytalone. Ces deux dernières molécules sont également produites par P. aleophilum [137]. Le fait qu'elles soient produites par P. chlamydospora et P. aleophilum justifie qu'elles aient été très étudiées quant à leur implication dans l'expression des symptômes foliaires [137]. Par comparaison, F. mediterranea produit peu de molécules dont les deux principales sont la 6-methoxymelleine et le 2,2-dimethyl-4-oxochoman-6-carbaldehyde [61].
La fonction de ces diverses molécules n'est pas clairement déterminée. En effet, il est difficile de reproduire des symptômes en conditions contrôlées à l'aide de ces molécules.
2.1.3.3 Les réactions de la plante.
En réponse à une agression par des pathogènes, la plante met généralement en place des défenses afin de limiter leur progression. Shigo et Marx [138] expliquent dans leur modèle appelé CODIT comment la plante peut compartimenter les microrganismes. Le modèle est basé sur quatre barrières dont les trois premières servent à restreindre la progression des microrganismes dans le bois présent au temps de la blessure. Elles sont attribuées (i) à l'occlusion des vaisseaux par des thylles et des substances gommeuses, cette première barrière est discontinue et constitue la plus faible des barrières limitant la propagation verticale des microrganismes ; (ii) aux caractères morphologiques des dernières cellules en fin de chaque anneau de croissance ; cette barrière est continue sur toute la longueur de l'arbre et limite l'expansion parallèle aux rayons des microrganismes ; (iii) aux rayons parenchymateux, tissus vivants du bois, qui peuvent déclencher des réactions de défense de la plante, cette troisième barrière est la plus forte des trois mais peut-être discontinue ; elle limite la progression perpendiculaire aux rayons des microrganismes ; la quatrième barrière, la plus forte et la plus importante des barrières, est formée par le cambium après blessure. Elle empêche la progression des champignons dans le nouveau tissu formé.
Dans le cadre de l'esca, la période de latence entre l'inoculation des champignons et l'expression des symptômes foliaires est de plusieurs années, ce qui suggère un contrôle de la propagation des champignons par la plante suite à l'induction de défenses et/ou la production de molécules spécifiques.
A l'heure actuelle, peu d'études ont été réalisées sur les interactions hôte-parasite dans le cas de l'esca. L'observation des nécroses de ceps de Cabernet-Sauvignon prélevés in natura par des méthodes histochimiques montre que la plante met en place différentes barrières pour limiter la progression des champignons dans le bois [28] : des thylles et des gommes dans les vaisseaux et l'augmentation de composés phénoliques dans les tissus ligneux au niveau de la zone brun rose. Amalfitano et al. [139] constatent une accumulation de trans-resvératrol et de ε-viniférine dans ces mêmes zones sur le cépage Sangiovese. Les formations de thyllose associées à des changements de taux et de composition de composés phénoliques ont été mises en relation avec la sévérité de l'infection dans des racines infectées par P. chlamydospora et plusieurs espèces de Phaeoacremonium [139]. Cette accumulation de composés phénoliques dans les tissus infectés par les pathogènes est probablement régulée par un équilibre entre la production par les cellules végétales et la détoxication par les pathogènes. Sur des boutures inoculées par P. chlamydospora, Larignon [28], Pascoe et Cottral [48] et Troccoli et al. [49] constatent la formation de gommes et de thylles dans les vaisseaux, et la présence de substances non identifiées, probablement de type stilbène [49]. Bruno et al. [140] ont observé une quantité de resvératrol dans les cals infectés par P. chlamydospora ou F. mediterranea par rapport aux taux présents dans des cals non infectés. Contrairement à la théorie courante, ce modèle suggère que le resvératrol est facilement dégradé par les champignons associés à l'esca et donc n'est plus capable de protéger la plante infectée [140].
Au niveau moléculaire, très peu de données sont disponibles quant à la reprogrammation transcriptomique au niveau des tissus infectés par les pathogènes de l'esca. Par conséquent, la caractérisation des interactions hôte-pathogène n'en est qu'à ses débuts.
2.2 Black Dead Arm
2.2.1 Symptomatologie
2.2.1.1 Historique de la maladie.
Le Black Dead Arm connu depuis très longtemps sous le nom d'apoplexie lente [11] est encore souvent confondu avec l'esca car la symptomatologie est très similaire. Il est présent dans différents pays viticoles : Espagne [141], Italie [45,46], France [12], Chili [44] et Liban [47] (tableau 1).
Le terme Black Dead Arm a été introduit par Lehoczky [142] pour distinguer cette maladie du dead arm disease qui est provoqué par Phomopsis viticola Sacc. Ce terme a ensuite été repris par Cristinzio [45] pour désigner les symptômes foliaires qui sont caractérisés par des digitations internervaires sans liseré jaune dans les premiers stades d'apparition des symptômes chez les cépages noirs. Sur les cépages noirs, le suivi des feuilles tout au long de la période végétative montre que les tissus de couleur rouge vineux s'éclaircissaient pour devenir rouge clair, puis présentaient à leur périphérie des jaunissements [12], cette observation suggère à Lecomte et al. (2006) [143] que le faciès BDA etait une phase transitoire de l'expression de la forme lente de l'esca. Des études sur le profil anthocyanique de feuilles atteintes par le BDA ou l'esca à des stades précoces d'expression foliaire montrent qu'ils sont différents, signe de dérèglements du métabolisme foliaire, spécifiques de chaque dépérissement [144]. Ces symptômes sont en relation avec une bande brune dans le bois située sous l'écorce pouvant aller jusqu'au niveau de la soudure, voire du porte-greffe [12]. Cette bande brune peut se former de part et d'autre d'une nécrose sectorielle et se traduire ensuite soit par la présence de taches noires, soit par la formation d'un chancre. Une forme sévère caractérisée par une défoliation des rameaux est également connue [12].
2.2.1.2 Comportement des cépages.
Peu d'informations sont données sur la sensibilité des cépages. Il est connu que les cépages Cabernet Sauvignon, Cabernet franc, Sauvignon blanc sont plus sensibles à ce dépérissement que le Merlot [145].
2.2.1.3 Influence des facteurs environnementaux.
Peu d'informations sont disponibles sur les facteurs environnementaux ayant un rôle majeur dans le développement de la maladie et de son expression. La manifestation des symptômes foliaires est fluctuante d'une année sur l'autre, les étés doux et pluvieux étant favorables à leur extériorisation.
2.2.2 Les champignons associés
2.2.2.1 Historique.
Le BDA a d'abord été associé à Diplodia mutila Fr. (téléomorphe « Botryosphaeria » stevensii Shoemaker) dans le vignoble du Tokay, en Hongrie [142]. Puis, il fut associé à d'autres espèces de Botryosphaeria dans différents vignobles : Diplodia seriata De Not. (téléomorphe « Botryosphaeria » obtusa (Schw.) Shoemaker) en Italie [46], en Espagne [41], en France [146] et au Chili [44] et Botryosphaeria dothidea (B. ribis Gross. & Duggar) en France, ensuite identifié sous le nom de Neofusicoccum parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips comb. nov. [146] (tableau 1). Ces champignons sont également trouvés dans des chancres chez des vignes dépérissantes [147].
2.2.2.2 Processus menant à la dégradation du bois.
Le processus de dégradation du bois n'a pas encore été décrit.
2.2.2.3 Tests de pathogénie.
Les symptômes de BDA sur la partie herbacée semblent avoir été décrits par Cristinzio [45] deux mois après l'inoculation dans le vignoble (B. obtusa) sur le cépage Sangiovese. Cependant, cet auteur apporte peu d'informations sur les méthodes d'inoculation, le stade physiologique de la plante, etc. Rovesti et Montermini [46] ont reproduit un brunissement du cambium et des vaisseaux respectivement sept et douze mois après l'inoculation de « B. obtusa ». Castillo-Pando et al. [148] montrent qu'après inoculation sur plantule, « B. obtusa » provoque un affaiblissement de la végétation conduisant à sa mort. Sur boutures, neuf mois après inoculation, aucun symptôme n'est obtenu sur la partie herbacée, seules des ponctuations noires dans le bois ont pu être reproduites. Auger et al. [44] ont reproduit les symptômes de la bande brune en injectant une suspension du mycélium de « B. obtusa » dans des boutures de V. vinifera var. Red Globe. Rego et al. [149] montrent, par inoculation de boutures cultivées en conditions contrôlées, que les champignons de la famille des Botryosphaeriaceae provoquent des lésions dans les tissus ligneux et des rougissements sur les feuilles un an après l'inoculation des cépages Tempranillo et Castalão. D'autres expérimentations ont été réalisées pour montrer la capacité des Botryosphaeriaceae à provoquer des symptômes dans le bois [146,150,151].
2.2.2.4 Caractéristiques et cycles biologiques des champignons
Botryosphaeria obtusa, B. stevensii et N. parvum
Ces champignons sont présents dans la majorité des vignobles (tableau 1). Ils sont rencontrés sur différentes plantes : Vitis spp, arbres fruitiers, … et peuvent provoquer un grand nombre de dépérissements [152]. Ils appartiennent à la Classe des Ascomycètes de la famille des Botryosphaeriacées. La description de ces champignons et leurs conditions de culture sont décrites par différents auteurs [153–157]. Peu d'informations sont données sur le type de pourriture qu'ils provoquent dans le bois et sur leur équipement enzymatique. B. obtusa secrète du tyrosol, des mélléines, des dérivés de la mélléine et du 4-hydroxybenzaldéhyde [158,159]. Cependant, aucune relation entre la pathogénie et le taux de toxines produites n'a été mise en évidence. Aucun élément n'est apporté au sujet des déterminants de leur pouvoir pathogène.
Leur cycle biologique sur vigne est peu connu. Ils se conservent sous forme de pycnides qui sont localisées soit sur le cep (tronc, bras, plaies de taille), soit sur les bois de taille laissés au sol [145]. Ils se disséminent plus particulièrement pendant la période végétative de la vigne [160]. Ils peuvent également se propager par les bois de porte-greffes ou de greffons en pépinières [63]. Des analyses microbiologiques réalisées dans les tissus ligneux de sarments de porte-greffes montrent la présence de « B. obtusa » et de N. parvum sur toute leur longueur [63], suggérant que ces champignons peuvent contaminer le rameau ou le sarment à différents niveaux soit pendant la période végétative, soit pendant la phase de repos avant la récolte du matériel végétal. Ces champignons sont également présents à la surface du matériel de propagation (greffon, porte-greffe) comme le montrent les tests de détection par PCR [77]. Une des phases au cours desquelles se produisent les contaminations lors de l'élaboration des plants a été identifiée : il s'agit de la stratification. La pénétration se fait par les plaies situées au niveau de la future soudure ou celles causées par le débitage [77].
2.2.3 Interactions hôte-pathogène
Aucune étude spécifique caractérisant l'impact du BDA sur différents métabolismes de la plante (métabolisme carboné, hydrique ou encore la mise en place de défenses) n'a été publiée.
Les travaux dédiés à la recherche et l'identification de molécules toxiques produites par les différentes espèces de Botryosphaeria sont très récents et limités. En effet, Martos et al. [161] ont isolé pour la première fois des molécules de nature polysaccharidique. Ces molécules développent une forte toxicité lorsqu'elles sont testées sur le modèle simplifié feuille isolée [161]. Récemment, Djoukeng et al. [159] ont isolé 4 molécules qui sont respectivement la mélléine, les 4- et 7-hydroxymélléine et la 4,7-dihydroxymélléine. Elles ont également montré une forte toxicité lors de tests sur disques foliaires. Djoukeng et al. [159] ont également observé que B. obtusa oxydait le resveratrol en δ-viniferine dans le bois, ce qui indique une forte activité laccase. Ces expériences ayant été menées uniquement en conditions contrôlées, il faut maintenant caractériser la production de ces composés dans des plantes naturellement infectées.
3 Les méthodes de lutte actuelles
3.1 Historique
Les maladies de la souche, préoccupent depuis toujours les vignerons. C'est ainsi qu'à l'avènement de notre ère, les agronomes préconisaient afin d'empêcher “les pourritures” qui étaient une des causes de la mort des ceps, d'éviter ou de protéger les grandes blessures de taille [162,163]. Ils recommandaient également de guérir les souches en train de pourrir soit par curetage, soit par recépage. La constante préoccupation de protéger les plaies de taille ou encore de diminuer leur nombre et leur surface, ou de rechercher une période de taille idéale, se retrouve chez les agronomes d'hier [164–166] et les phytopathologistes d'aujourd'hui. A l'aube du XXème siècle, la lutte à l'égard de l'esca a trouvé son originalité dans la découverte tout à fait fortuite de l'efficacité de l'arsénite de sodium [167,168]. Ce produit était toxique vis-à-vis des champignons associés aux maladies du bois. Il était capable de tuer la plupart d'entre eux dans le xylème comme le montrent les travaux de Larignon et al. [169]. Actuellement, ce produit est interdit dans tous les pays viticoles du fait de sa toxicité pour le viticulteur [3].
3.2 Evaluation de produits chimiques et biologiques
Suite à son retrait en novembre 2001, la recrudescence des problèmes de dépérissement (Esca, Black Dead Arm) et l'apparition de ces problèmes sur des vignes de plus en plus jeunes ont contribué à la reprise des travaux pour trouver un substitut à l'arsénite de sodium. Les recherches menées par différents laboratoires dans le monde ont porté sur l'évaluation de l'efficacité de molécules chimiques et de produits biologiques aussi bien au laboratoire [170–173] que dans le vignoble, appliqués sous différentes formes (sécateur traitant, injection, protection des plaies par badigeonnage et par pulvérisation, pulvérisation sur toute la souche, peinture) [174–176] et à différents stades de la culture de la vigne.
Dans le vignoble, l'évaluation de l'efficacité des matières actives et des produits biologiques appliqués, soit en curatif sur des vignes malades soit en préventif dès l'implantation de la vigne, est basée sur l'observation des symptômes sur la partie herbacée. Des méthodes d'évaluation plus rapides, tout au moins concernant les méthodes préventives (protection des voies de pénétration du champignon dans la plante) ont été mises au point dans différents pays, plus particulièrement pour Eutypa lata, agent responsable de l'Eutypiose. Ces méthodes consistent à inoculer les plaies protégées par les produits à tester par des spores du champignon. L'évaluation de l'efficacité du produit est réalisée par des analyses microbiologiques via les méthodes classiques ou moléculaires [177]. Ces méthodes ont ensuite été reprises pour les autres champignons pénétrant par les plaies de taille : P. chlamydospora, Botryosphaeriaceae [178]. En France, une variante de cette méthode est désormais utilisée pour évaluer rapidement l'efficacité [120] et fait l'objet de la méthode CEB no 155 [version 1990]. Cette méthode a permis de mettre sur le marché en protection des plaies de taille à l'égard d'Eutypa lata [AMM] l'Atemicep (association de cyproconazole et de carbendazime) [179], qui fut retiré du marché suite à sa phytotoxicité, puis en 1996 l'Escudo (association de flusilazole et de carbendazime), qui a été ensuite homologué pour l'esca (mais peu utilisé par les viticulteurs du fait de son fort coût de main d'œuvre car le produit est appliqué par badigeonnage) et retiré du fait de sa toxicité en 2007. Cette méthode ne permet pas cependant de tester des produits comme des mastics ou encore des microrganismes qui pourraient être utilisés dans la lutte biologique. Une amélioration a été apportée en tenant compte de ces inconvénients [180]. Cependant, cette nouvelle méthode CEB no 155 version 2006 doit être améliorée en prenant en compte les conditions climatiques nécessaires à la contamination des plaies lors de l'essai du produit [122].
Les essais menés jusqu'à maintenant par les différents laboratoires dans le monde n'ont pas permis de trouver de méthodes de lutte curatives ou préventives satisfaisantes utilisant des produits chimiques pour lutter contre l'esca, parce que soit les produits testés ne sont pas efficaces, soit leur mode d'application n'est pas réalisable par le viticulteur.
Les recherches portent de plus en plus sur l'effet de micro-organismes utilisés en lutte biologique, notamment les Trichoderma. Ces champignons présentent une activité antagoniste et d'hyperparasitisme à l'égard d'un grand nombre de microrganismes, plus particulièrement ceux du sol et sont utilisés en lutte biologique à l'égard de diverses maladies sur différentes cultures [181]. Leur apport dans le sol a été signalé comme bénéfique à la croissance des plantes [181]. Ils peuvent jouer également un rôle dans l'élicitation des mécanismes de défense de la plante [182]. Ils ont été signalés comme capables de protéger les plaies de taille à l'égard des champignons pathogènes provoquant des pourritures du bois, notamment chez le Quercus rubrum [183] et les arbres fruitiers (maladies du plomb dues à Stereum purpureum) [184]. Smith et al. [185] rapportent également que Trichoderma harzianum est efficace pour protéger les plaies de taille à l'égard de Phialophora melinii chez le chêne rouge. Chez la vigne, Hunt et al. [186] montrent que l'application de Trichoderma sur les plaies de taille réduit fortement (85%) la colonisation du bois par E. lata. Di Marco et al. [187] montrent que l'application d'une souche de T. harzianum sur les plaies de taille empêche le développement de P. chlamydospora. D'autres microrganismes ont été testés uniquement à l'égard de E. lata : Bacillus subtilis [188], Cladosporium herbarum [177], Fusarium lateritium [177] en association ou non avec du bénomyl [189].
Dans les conditions de la pratique, aucun essai n'a pu montrer l'efficacité de tels produits en protection des plaies de taille face à l'esca ou au BDA.
3.3 En pépinières
Des recherches sur les méthodes de lutte sont menées également au niveau de la production des plants en pépinières car les champignons pionniers de l'esca et ceux associés au BDA se propagent par le matériel végétal [63]. Les traitements, qu'ils soient biologiques, chimiques ou physiques, ont été testés à différentes étapes de l'élaboration des plants.
Parmi les traitements effectués sur le matériel de propagation, le traitement à l'eau chaude réduit l'incidence de P. chlamydospora lorsqu'il est effectué à une température de 50 °C pendant 30 min [190,191]. Fourié et Halleen [192] montrent également que des bains de bénomyl, de captane ou de didecylméthylammonium chloride, réduisent sa présence dans les tissus ligneux au niveau de la soudure et à la base du plant. L'utilisation d'autres substances actives comme l'eau de Javel, l'oxyquinoléine, des Trichoderma ne montre pas d'efficacité vis-à-vis des champignons associés aux maladies du bois [193].
Des traitements sont également effectués sur les plants à la sortie de la pépinière. Le traitement à l'eau chaude effectué à une température de 50 °C pendant 45 min montre une bonne efficacité vis-à-vis de P. Chlamydospora et de B. obtusa, mais il ne présente pas d'efficacité à l'égard de P. aleophilum et de N. parvum [193].
La production de plants de bonne qualité sanitaire résultera de la conjugaison de diverses méthodes de lutte ou de l'utilisation de nouveaux process.
Une autre approche a été récemment abordée dans l'objectif est de produire des vignes plus résistantes par application de produits chimiques ou de produits biologiques en pépinières et/ou sur jeunes plantations [194]. Fourié et al. [195] montrent que T. harzianum, appliqué en pépinière, permet un meilleur développement racinaire des plants de vigne et ainsi une meilleure protection à l'égard de P. chlamydospora. Feirreira et al. [196] indiquent que les vignes présentant un meilleur système racinaire résisteraient mieux à des situations de stress et par conséquent seraient plus résistants aux maladies liées aux stress, en particulier la maladie de Petri dont l'agent responsable est P. chlamydospora. Actuellement, aucune expérimentation menée dans le vignoble n'a permis d'affirmer cette hypothèse.
3.4 Incidence des pratiques culturales
La recherche de méthodes culturales limitant l'incidence des maladies du bois a été peu développée. Certains systèmes de taille, notamment la taille guyot-poussard, semble limiter l'esca [197,198]. Contrairement à l'eutypiose [199], la taille tardive ou la protection des plaies de taille par badigeons ne permet pas de limiter l'esca ou le BDA. La restauration des souches par le recépage est efficace pour l'eutypiose [200] mais aucune expérimentation n'a permis de montrer son efficacité vis-à-vis de l'esca et du BDA. Le rôle des sarments laissés au sol, qui peuvent héberger les sources d'inoculum des champignons associés aux maladies du bois, sur la propagation de ces maladies n'est pas connu. Le compostage des sarments permet cependant d'éradiquer les champignons associés aux maladies du bois [201].
4 Conclusion
Cette revue nous montre que l'esca et le BDA sont le fruit d'un complexe de champignons déjà décrit à la fin du XXème siècle. Depuis une dizaine d'années, on observe dans le monde entier une augmentation des symptômes liés à ces champignons. Une absence de lutte efficace et probablement des facteurs environnementaux ne font qu'aggraver cette situation. Toutefois, il est intéressant de noter que certains vignobles jamais traités par l'arsénite de sodium, seul traitement ayant potentiellement une action contre le developpement des maladies du bois, présentent aujourd'hui une recrudescence des symptômes liés à ces maladies. Il n'y a donc pas eu dans ce cas de pression de sélection expliquant l'absence de développement des symptômes malgré la présence des champignons. Pourquoi ces symptômes apparaissent aujourd'hui : changement de la conduite de la vigne, changement climatique, pathogènes non diagnostiqués ? L'impossibilité de reproduire des symptômes avec les champignons connus doit du point de vue phytopathologique ne pas nous faire écarter cette dernière hypothèse. Ainsi, malgré des connaissances sur les champignons associés à la maladie, les interactions hôte-pathogène et la symptomatologie, il manque encore beaucoup de notions sur les réelles causes du développement des symptômes. Sans la compréhension des différents stades de la maladie, il ne paraît pas envisageable pour l'instant de lutter d'une façon durable contre cette infection. Doit-on penser après la pyrale de la vigne en 1825, le début de la crise de l'oïdium en 1845, les premières descriptions du phylloxera en 1863 et la crise du mildiou en 1879 que les maladies dites du bois seront une future crise sanitaire majeure ? Pour conclure on peut rappeler la citation de Charles Nicolle (1866–1936, Prix nobel de physiologie et médecine et membre de l'Académie de Sciences) « Ce qui est nouveau, ce n'est pas la survenue d'une maladie antérieurement inconnue, c'est cette survenue au sein d'un monde qui se croyait définitivement aseptisé, protégé et tranquille. »