

1 Introduction
The genus Hemidactylus Oken is the most speciose gekkonid genus in India, with at least 30 species and the number is steadily rising with exploration of new habitats, examination of museum material and use of molecular data [1–3]. Most of the species recorded from India are distributed south of Narmada valley, especially members of the genus exceeding 90 mm in snout-to-vent length (Mirza, pers. obs.).
Hemidactylus maculatus Duméril & Bibron is a large-sized gecko, which is a member of the “H. prashadi” clade of Bansal and Karanth [4]. The species was thought to be distributed across the Western Ghats from southern Gujarat to Tamil Nadu in addition to reports from Chhattisgarh and Andhra Pradesh [2,5–8]. Agarwal et al. [5] described the population from southern Karnataka and northern Tamil Nadu as H. graniticolus Agarwal, Giri & Bauer, and Mirza & Sanap [2] described the population from southern Tami Nadu as H. acanthopholis. Mirza & Sanap highlight that H. maculatus is a species complex. The population of H. maculatus from the Eastern Ghats was said to differ from true H. maculatus [5], and was further attested by Javed et al. [6].
While conducting herpetofaunal surveys across the central Indian State of Chhattisgarh, we collected specimens of a large Hemidactylus, which resembled superficially H. maculatus. Detailed comparison with the latter and similar large sized geckos, like H. graniticolus and H. acanthopholis, confirmed the distinctiveness of the specimens from Chhattisgarh, which are herein considered as a new species. The new species has appeared in the literature several times as being considered conspecific with H. maculatus, H. subtriedrus Jerdon [9], or was said to be distinct from H. maculatus, but was not formally described likely due the lack of comparative material as well as molecular data [6]. Fresh collection, museum material and available data in the literature as well as molecular data permit the description of a new species herein and further, allow us to add notes on the H. maculatus complex.
2 Material and methods
Specimens in the field were captured by hand and euthanized, followed by fixation in 6% formaldehyde buffer. The specimens were later washed, stored in 70% ethanol, and deposited in the collection of the Bombay Natural History Society, Mumbai, India.
Morphological and meristic data. All measurements were taken following Mirza & Sanap [2] with Mitutoyo™ digital calipers (Mitutoyo Corporation, Kawasaki, Japan) (to the nearest 0.1 mm): snout-vent length (SVL; from the tip of the snout to the vent), trunk length (TRL; distance from the axilla to the groin measured from the posterior edge of the forelimb insertion to the anterior edge of the hind limb insertion), body width (BW; maximum width of the body), crus length (CL; from the base of the heel to the knee); tail length (TL; from the vent to the tip of the tail), tail width (TW; measured at the widest point of the tail); head length (HL; distance between the retroarticular process of the jaw and the snout tip), head width (HW; maximum width of the head), head height (HH; maximum height of the head, from the occiput to the underside of the jaws), forearm length (FL; from the base of the palm to the elbow); ear length (EL; longest dimension of the ear); orbital diameter (OD; greatest diameter of the orbit), nares-to-eye distance (NE; distance between the anteriormost point of the eye and the nostril), snout-to-eye distance (SE; distance between the anteriormost point of the eye and the tip of the snout), eye-to-ear distance (EE; distance from the anterior edge of the ear opening to the posterior corner of the eye), internarial distance (IN; distance between the nares), interorbital distance (IO; shortest distance between the left and right supraciliary scale rows) (Table 1). Meristic counts and external observations of morphology were made using a Leica™ S8APO (Leica Camera, Wetzlar, Germany) dissecting microscope. The images of the specimens were taken with a Canon™ 70D mounted with a Canon™ 100 mm macro illuminated with two external Canon™ 430EX-II flashes (Canon Inc., Tokyo, Japan). The institutional acronyms used in the manuscript are as follows: BNHS, Bombay Natural History Society, Mumbai; CAS, Californian Academy of Sciences, San Francisco; NHM, Natural History Museum London; NCBS, Collection facility, National Centre for Biological Sciences, Bangalore; ZSI, Zoological Survey of India, Kolkata. The Life Science Identifiers (LSID) for this publication are: urn:lsid:zoobank.org:pub:E51AF8B4-CF5C-46DE-95F7-5563D8A8FE52.
Morphological and meristic data for type specimens of Hemidactylus kangerensis sp. nov.
| Holotype | Paratypes | |||||
| Specimens numbers | BNHS 2484 | BNHS 2485 | BNHS 2486 | BNHS 2487 | BNHS 2488 | BNHS 2489 |
| Sex | Male | Male | Male | Female | Female | Female |
| SVL | 95.0 | 89.0 | 74.0 | 95.0 | 85.0 | 64.0 |
| TRL | 38.0 | 36.0 | 29.7 | 42.3 | 32.8 | 24.7 |
| BW | 22.2 | 20.7 | 16.1 | 26.0 | 21.6 | 12.3 |
| CL | 15.6 | 14.4 | 11.7 | 14.7 | 12.5 | 11.4 |
| TL | 115.0 | 151.0 | 28.5a | 163.0 | 93.6 | 77.5 |
| TW | 12.6 | 11.6 | 8.3 | 15.4 | 12.8 | 7.4 |
| HL | 25.5 | 22.8 | 17.2 | 24.7 | 18.3 | 15.9 |
| HW | 21.0 | 19.9 | 14.7 | 20.9 | 17.5 | 12.4 |
| HH | 12.7 | 12.4 | 8.7 | 12.8 | 9.0 | 6.3 |
| FL | 13.0 | 12.1 | 9.0 | 12.3 | 11.1 | 8.4 |
| OD | 5.8 | 5.4 | 4.3 | 5.6 | 4.8 | 4.3 |
| NE | 9.6 | 8.6 | 2.0 | 8.7 | 7.6 | 5.4 |
| SE | 12.0 | 10.9 | 8.6 | 11.0 | 9.6 | 7.0 |
| EE | 8.6 | 7.2 | 5.0 | 8.1 | 7.2 | 3.9 |
| EL | 2.7 | 2.4 | 1.7 | 2.4 | 1.9 | 2.1 |
| IN | 2.7 | 2.7 | 2.1 | 2.6 | 2.3 | 1.9 |
| IO | 9.3 | 8.5 | 7.1 | 9.5 | 6.7 | 5.7 |
| Lamellae | ||||||
| L manus | 10–12–12–13–12 | 10–10–11–11–11 | 10–11–12–12–11 | 10–12–12–12–10 | 10–11–11–11–11 | 9–11–12–12–11 |
| R manus | 10–12–12–12–11 | 10–11–12–12–12 | 9–11–11–10–11 | 9–12–12–12–11 | 8–11–12–11–11 | 11–12–12–12–9 |
| L pes | 9–13–13–12–11 | 9–11–12–12–12 | 9–12–12–12–12 | 9–13–13–13–12 | 8–12–12–12–11 | 8–12–12–13–11 |
| R pes | 10–12–12–11–11 | 9–12–14–13–11 | 9–12–12–12–11 | 9–13–13–13–12 | 8–12–12–12–11 | 9–12–12–12–11 |
| Supralabials | ||||||
| L | 12 | 12 | 13 | 11 | 12 | 12 |
| R | 12 | 13 | 12 | 11 | 11 | 13 |
| Infralabials | ||||||
| L | 9 | 9 | 11 | 9 | 10 | 10 |
| R | 9 | 10 | 10 | 10 | 9 | 10 |
| Pores L/R | 18/19 | 21/21 | 18/18 | |||
| Diastema of non–pored scales | 4 | 4 | 4 |
a denotes broken or regenerated tail.
Molecular methods and analysis: Genomic DNA was extracted from the tail tissue of the paratype male BNHS 2486 using Qiagen DNeasy™ tissue kit, following protocols specified by manufactures. Partial mitochondrial cytochrome b gene was amplified using primer Cytb1 (5′-CCATCCAACATCTCAGCATGATGAAA-3′) and Cytb2 (5′-ACTGTAGCCCCTCAGAATGATATTTGTCCTCA-3′) published by Kocher et al. [10]. Polymerase Chain Reaction (PCR) reaction totaled 10 μL with H2O comprised of 4 μL, 4 μL of Qiagen™ Taq PCR Master Mix, 0.5 μL of each primer and 2 μL of DNA template. The reaction was carried out with an Eppendorf Mastercycler Nexus GSX1. The thermocycle profiles used for amplification were as follows: 94 °C for 15 min, (94 °C for 50 s, annealing temperature 48 °C for 50 s, 72 °C for 2 min) × 35 cycles, 72 °C for 15 min, hold at 4 °C. The PCR product was cleaned, and sequenced with a 3730 DNA Analyzer after cleaning. The sequence was cleaned manually in Mega7 [11]. The sequences were aligned in Mega7 using ClustalW [12] with default settings. For optimal partitioning strategy and evolutionary substitution model, the aligned data were analyzed using PartitionFinder v.1.1.1 [13]. The Maximum Likelihood method was implemented to assess phylogenetic relationship with RAxML [14]. The data were partitioned into three codons and GTR + G was used as the sequence substitution model, based on the optimal partitioning scheme suggested by PartitionFinder. Maximum likelihood analysis was run for 1000 non-parametric bootstrap replicates with rapid ML search option. Bayesian Inference was implemented in MyBayes 3.2.2. [15] and was run for 10 million generations and was terminated after the analysis has reached a standard split frequency of 0.01. Twenty-five percent of the trees generated were discarded as burnin. The sequence divergence uncorrected “p-distance” was calculated in Mega7. The sequence for the new species has been deposited with GenBank with accession number KY938009.
3 Systematics
Hemidactylus kangerensissp. nov.Hemidactylus maculatus Sanyal & Dasgupta 1990: 18; Ingle 2003:2
Hemidactylus subtriedrus Sanyal & Dasgupta 1990: 18–19; Javed et al., 2009: 368 (in part)
Hemidactylus cf. maculatus Mahony 2009: 60–61; Javed et al., 2011: 10 (in part)
Hemidactylus sp. Agarwal et al., 2011: 36; Mirza & Sanap 2014: 16 (in part)urn:lsid:zoobank.org:act:BD6A980D-FB2A-4111-B920-96934A61068B

Hemidactylus kangerensis sp. nov. dorsal view of male holotype BNHS 2484. Scale bar: 20 mm.

Hemidactylus kangerensis sp. nov. male holotype BNHS 2484 head, (A) lateral view, (B) dorsal view, (C) ventral view. Scale bar: 5 mm.
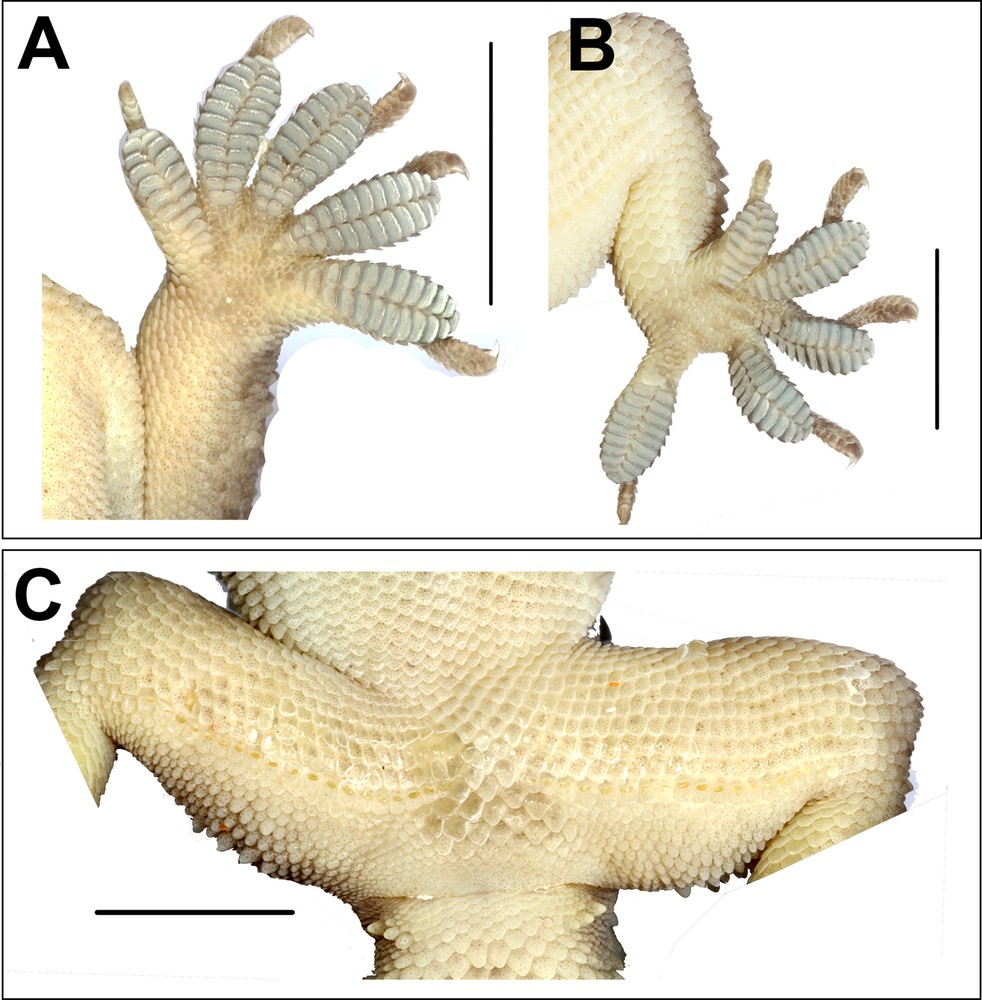
Hemidactylus kangerensis sp. nov. male holotype BNHS 2484, (A) left manus showing lamellae, (B) left pes showing lamellae, (C) ventral view of thigh and cloacal region showing femoral pores. Scale bar: 10 mm.

Ventral view of thigh and cloacal region showing femoral pores, (A) Hemidactylus kangerensis sp. nov. holotype BNHS 2484, (B) H. graniticolus NHM 1946.8.23.72, (C) H. acanthopholis CAS 104311, (D) H. sykesii (= H. maculatus) NHM XXII.20a.

Dorsal scales on trunk, (A) H. kangerensis sp. nov. holotype BNHS 2484, (B) H. granaticolus NHM 1946.8.23.72, (C) H. hunae NHM 1946.8.23.77 (D) H. acanthopholis NHM 1946.8.23.68, (E) H. maculatus NHM 1956.1.11.41.
Holotype BNHS 2484, adult male, in a forest rest house near Kailash caves, Kanger Valley National Park, Bastar District, Chhattisgarh, India (18.841170°, 81.997310°, 464 m). Collected on 2 December 2016 by Harshal Bhosale and Zeeshan Mirza.
Paratypes (five specimens): Two males (BNHS 2485, BNHS 2486); three females (BNHS 2487, BNHS 2488, BNHS 2489) collected from Kanger Valley National Park, Bastar District, Chhattisgarh (18.841170°, 81.997310°, 464 m). Collected on 2 December 2016 by Harshal Bhosale and Zeeshan Mirza.
Referred specimens: ZSI 24151 Katamawas, ZSI 25506, Forest rest house, ZSI 25513, Bhadrimanu, all from Jagdolpur, Bastar district, Chhattisgarh, ZSI 24155 (female), Bastar District, Chhattisgarh, India; NHM 1874.11.11.1 Khamman District, Northern Telangana (earlier Andhra Pradesh).
Diagnosis: A large-sized species of the genus, SVL at least 95 mm. Dorsal scalation on the trunk, granular, intermixed with enlarged, fairly regularly arranged, longitudinal rows of 18–20 strongly keeled, trihedral, tubercles of equal size of dorso-lateral aspect. Two pairs of postmentals, inner pair twice as long and wide as the outer pair. Scales on trunk venter arranged in 28–30 rows. Lamellae divided, covering 80% of the digit. Caudal pholidosis, dorsal aspect, small, juxtaposed, intermixed with large strongly keeled trihedral tubercles, eight or six in number on each segment of the tail, scales sub-equal throughout the original tail. Femoral pores 18–21 on each side separated medially by four non-pored scales. Three subconical post-cloacal spurs, anterior spur larger than the preceding spurs; posterior spur smallest more rounded.
Etymology: The species is named after Kanger Valley National Park, where the type locality is located.
Description of holotype male BNHS 2484 (Fig. 1): Holotype in good condition preserved in a linear manner with a slightly curved tail. Part of the tail regenerated (Fig. 1).
A large-sized gecko (SVL 95 mm) with a fairly large head (HL/SVL ratio 0.27), head slightly longer than wide (HW/HL ratio 0.82), head depressed (HH/HL ratio 0.50), distinct from the neck (Fig. 2A and B); canthus rostralis slightly inflated; snout short (SE/HW ratio 0.57), obtusely pointed from dorsal view and acutely in lateral view (Fig. 2A); longer than the eye diameter (OD/SE ratio 0.48); scales on the snout subequal, convex, those anterior to the eye and on canthus rostralis, larger than the surrounding scales; eyes large (OD/HL ratio 0.23), pupil vertical with crenulated edges; supraciliaries larger on the anterior edge of the orbit, gradually decreasing in size as they progress towards the posterior portion of the orbit; ear-opening large, sub-oval, obliquely oriented, length less than half of the orbital diameter (EL/OD ratio 0.46) lobules absent; eye-to-ear distance greater than the diameter of the eye (EE/OD ratio 1.48); rostral quadrangle, much wider than deep, divided by a median suture for its entire length; rostral in contact with nasal, first supralabial and internasals; two large and a slightly smaller internasal between nasals; mental triangular, wider (4.4) than long (3.8); two pairs of postmentals, anterior postmental longer (2.6) than wide (1.8); posterior pair of postmental slightly smaller than the anterior pair, wider (2.4) than long (2.1); anterior postmental in contact with the mental, infralabials and posterior pair of postmentals; posterior postmentals less than half the size of the anterior one; anterior postmental slightly smaller than the width of the first infralabial; posterior postmental half the width of the second infralabials; posterior postmental in contact with the anterior postmental, the first as well as the second infralabial (Fig. 2C); scales on the throat circular, smaller than the ones on the ventral aspect of the trunk; ten supralabials (to midorbital position) on the left and right sides; 12 supralabials (to the angle of the jaw) on the left as well as on the right side; nine infralabials (to the angle of the jaw) on either side.
Body elongate (TRL/SVL ratio 0.40) and dorsoventrally flattened; lacking distinct ventrolateral furrow; dorsal scalation on the granular trunk intermixed with enlarged, keeled, trihedral tubercles, fairly arranged in 18–20 longitudinal rows; dorsal tubercles on mid-dorsum situated in close proximity, slightly depressed, slightly longer (1.4) than wide (1.3); tubercles on the lateral aspect of the larger trunk, slightly spaced out in comparison with tubercles on mid-dorsum; ventral scales on the trunk, smooth, flat, larger than dorsal scales; mid-body scales across belly 28–30; eighteen (left) and nineteen (right) femoral pores separated at mid-pelvic region by a diastema of four non-pored scales; non-pored scales slightly larger than pored scales (Fig. 3C).
Limbs moderately long, stout; digits dilated, bearing horizontally oriented lamellae on the ventral surface; lamellae on the basal half of digit I of manus and pes undivided, lamellae on rest other digits divided (excluding terminal lamellae); clawed, claw slightly smaller than the length of the lamellar region; forelimbs short (FL/SVL ratio 0.14), slightly shorter than hind limbs (CL/SVL ratio 0.16). Terminal phalanx of all digits curved, arising angularly from the distal portion of the expanded lamellar pad, free portion of phalanx of all digits half to more than half long as the dilated portion. Lamellae beneath the digits, left manus 10–12–12–13–12 (Fig. 3A), right manus 10–12–12–12–11; left pes 9–13–13–12–11, right pes 10–12–12–11–11 (Fig. 3B). Lamellae not reaching the base of the digit IV of the pes, covering 80% on the digit. Relative lengths of digits: III > V > IV > II > I (left manus), V > II > IV > III > I (left pes).
Tail moderately depressed, oval in cross section, longer than the snout–vent length (TL/SVL ratio 1.21), 42.5 mm of the tail regenerated. Caudal segments distinct; pholidosis of the original tail dorsum with small, juxtaposed scales intermixed with large keeled trihedral tubercles, scales on the regenerated portion of the tail homogenous, slightly smaller compared to the scales on the original tail and lacking tubercles. First tail segment with a whorl of eight large conical, keeled tubercles, second segment onwards, each segment with six tubercles. Ventral aspect with large, broad scales covering about ∼60% of the tail width from the base of the tail to the tip (Fig. 3C). Three subconical post cloacal spurs, anterior spur larger than the preceding spurs; posterior most spur smallest more rounded.
Coloration (Figs. 6, 7 & 8A): (in life) Background coloration in shade of light brown. Four distinct broad dark brown bands, each edged with black on its anterior and posterior edges, present between the nape and the caudal constriction. One more band that coincides with the caudal constriction is slightly diffused and does not possess defined edges. Similar bands present on the original part of the tail. Tubercles on dorsum with black tips. Supraciliaries and scales on canthus rostralis yellowish as well as scales in a row posterior to the orbit, appearing as a yellow strip from the nares to the temporal region. Limbs in shade of light brown, tubercles on these slightly darker. Ventrally white. (In preservative, Figs. 1–3.) Coloration much paler than in life. Dark dorsal bands faint, almost of the same color as the overall coloration, only black borders of these broad bands visible. A faint pale stripe between the orbit and nares present. White below.

H. kangerensis sp. nov. holotype BNHS 2484 in life. Photo by Zeeshan Mirza.

H. kangerensis sp. nov. coloration in life (A) paratype female BNHS 2487, (B) paratype male BNHS 2486. Photos by Zeeshan Mirza.

Coloration and habitus of similar looking large Hemidactylus from India, (A) H. kangerensis sp. nov. Photo by Zeeshan Mirza, (B) H. acanthopholis Photo by Deepak C.K., (C) H. graniticolus Photo by Atish Gawai, (D) H. maculatus. Photo by Zeeshan Mirza.
Distribution and Natural History. The type locality lies in a mixed moist deciduous forest and is dominated by Shorea robusta, Tectona grandis, and Bamboo species. During the survey conducted by us, all adult specimens were found in old houses, whereas the juveniles were found on termite mounds and tree trunks. During monsoons, even adults were seen on trees (S.M.M. Javed and Sunny Patil personal communication). Based on published records, museum specimens and our collection, it is certain that the new species occurs in two Indian states across the northern Eastern Ghats (Fig. 9): Chhattisgarh (Kanger Valley National Park 18.841170°, 81.997310°; Jagdalpur 19.065835°, 82.02634°; Sukma 18.390813°, 81.653934°), TELANGANA (Khamman 17.290373°, 80.26238°).
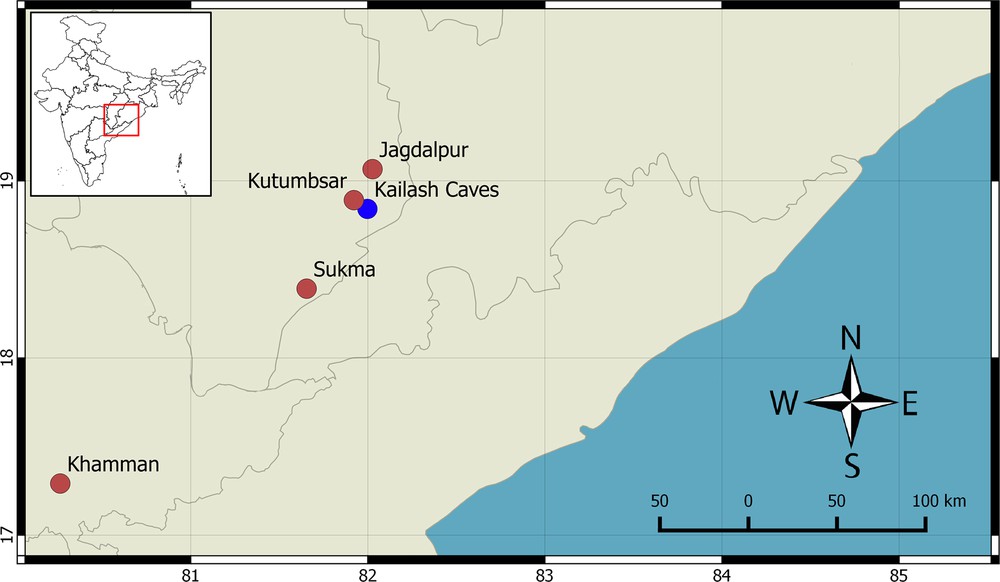
Map showing distribution of H. kangerensis sp. nov., type locality indicated by blue circle and other localities by brick red circles.
Suggested common name: Kanger valley rock gecko.
Variation: The paratypes agree with the description of the holotype in all respect. Detail morphometric and meristic data are presented in Table 1. A variation is seen in the number of lamellae on each digit, the number of labial scales, and the numbers of pores, which range in the type series from 18–21. Posterior postmental pair, less than half the width of the anterior pair; however, in the male specimen BNHS 2485, the posterior postmental is more than half the width of the anterior postmental.
Comparison:Hemidactylus kangerensissp. nov. differs from most Indian congeners in bearing the following set of differing and non-overlapping characters: SVL 95–121 mm (vs. SVL < 80 mm in H. aquilonius Zug & Mcmahan, H. flaviviridis Rüppell, H. frenatus Schlegel, H. garnotii Duméril & Bibron, H. leschenaultii Duméril & Bibron, H. sataraensis Giri & Bauer, H. gracilis Blanford, H. reticulatus Beddome, H. albofasciatus Grandison & Soman, H. scabriceps Annandale, H. persicus Anderson, H. robustus Heyden, H. turcicus Linnaeus, H. platyurus Schneider, H. anamallensis Günther, H. treutleri Mahony, H. gleadowi Murray, H. parvimaculatus Deraniyagala, H. kushmorensis Murray, H. murrayi Gleadow, H. triedrus Daudin, H. subtriedrus Jerdon, H. gujaratensis Giri, Bauer, Vyas & Patil); dorsum with large, keeled trihedral tubercles in 18–20 fairly regular longitudinal rows (vs. few smooth or rounded tubercles in H. giganteus Stoliczka, H. gujaratensis Giri, Bauer, Vyas & Patil, H. aaronbaueri Giri, H. yajurvedi Murthy, Bauer, Lajmi, Agarwal & Giri, H. hemchandrai Dandge & Tiple); dorsal pattern with fairly distinct bands and dorsal tubercles trihedral (vs. dorsal pattern with spots and dorsal tubercles subtrihedral in H. prashadi). The new species is most similar to H. maculatus, H. graniticolus, H. acanthopholis, and H. hunae Deraniyagala in general appearance and large SVL. Hemidactylus kangerensis sp. nov. differs from these as follows: femoral pores 18–21 separated by diastema of 4 non-pored scales (vs. 23–28 separated by a diastema of 1–3 poreless scales in H. graniticolus, 19–21 pores separated by a diastema of 12–14 poreless scales in H. acanthopholis, 19 pores separated by a diastema of 7 poreless scales in H. maculatus, Fig. 4); tubercles on dorsum enlarged, arranged in fairly regular longitudinal 18–20 rows strongly keeled, trihedral, tubercles (Fig. 5, vs. 16–18 rows of sub-trihedral, weakly keeled tubercles in H. graniticolus and H. hunae); dorsal pholidosis of tail comprised of small, pointed, keeled scales and a series of six or eight large keeled, trihedral tubercles (vs. small, imbricate, striated scales and a series of four enlarged, keeled and weakly striated and flattened tubercles in H. graniticolus and H. hunae). Hemidactylus kangerensis sp. nov. bears broad bark edged bands on its trunk (Fig. 8, vs. shaddle shaped bands on the trunk in H. graniticolus and H. acanthopholis, bands edged with big black spots, three each on the anterior and posterior edges in H. maculatus).
4 Discussion and conclusion
Hemidactylus kangerensis sp. nov. is morphologically similar to H. maculatus, H. graniticolus, and H. acanthopholis, and was found to belong to the Indian clade Tropical Asian clade the genus Hemidactylus [4], and was found to be embedded within the monophyletic H. prashadi group [4,5]. The suggested relationship is based on the external morphology and the partial sequence of the mitochondrial cytochrome b gene. Despite the general similarity in appearance, Hemidactylus kangerensis sp. nov. is geographically isolated in the northern Eastern Ghats from morphologically similar species, H. maculatus occurs in Northern Western Ghats, H. acanthopholis in southern Tamil Nadu, south of Palghat Gap, and H. graniticolus in parts of the southern Eastern Ghats and the Bangalore–Mysore plateau (Fig. 10). Furthermore, the new species shows an uncorrected pairwise sequence divergence of 10% from H. maculatus for cyt b (Table 2, Fig. 11). Phylogenetic relationships recovered from our analysis strongly support the inclusion of the new species in the “H. prashadi” group, but the relationships within the clade received poor support. The lack of samples could be one of the factor for poor support and hence the presented data must be treated merely preliminary.

Map of Peninsular India, showing distribution of large similar looking Hemidactylus. H. maculatus (green circles), H. acanthopholis (yellow circles), H. graniticolus (blue circles), and H. kangerensis sp. nov. (red circles).
Uncorrected pairwise sequence divergence based on the partial segment of the mitochondrial cytochrome b gene.
| Species | Pairwise sequence divergence (%) |
| Hemidactylus graniticolus (n = 2) | 14–16 |
| Hemidactylus prashadi (n = 5) | 15–17 |
| Hemidactylus hunae (n = 1) | 16 |
| Hemidactylus lankae (n = 1) | 16 |
| Hemidactylus maculatus (n = 1) | 10 |
| Hemidactylus triedrus (n = 5) | 13–16 |
| Hemidactylus depressus (n = 3) | 15–16 |
| Hemidactylus flaviviridis (n = 2) | 17–18 |

Maximum likelihood tree for members of the H. prashadi group showing relationship of H. kangerensis sp. nov. rooted with H. flaviviridis as an outgroup based on ∼284 bp of the mitochondrial cytochrome b gene with 1000 non-parametric bootstrap replicates. Black solid circles at nodes indicate high bootstrap (> 70) and Bayesian (> 0.90) support.
The status of the population currently considered conspecific with H. maculatus occurring in southern Maharashtra and Kerala require attention, which is out of the scope of the present study. Three specimens from Malabar are available, two in the collection of the Natural History Museum (NHM 1946.8.23.76 male, NHM 1946.8.20.3 juvenile female) and the Bombay Natural History Society (BNHS 1255). Agarwal et al. (2011) referred NHM 1946.8.23.76 as H. graniticolus and the specimens BNHS 1255 (1–3) and NHM 1946.8.20.3 as H. maculatus. Examination of NHM 1946.8.23.76 specimen confirms that it is not conspecific with H. maculatus given that the femoral pores in these specimens are separated by a diastema of three non-pored scales, a character seen in H. graniticolus. On the other hand, BNHS 1255 has a series of three specimens, of which one is a male. The male specimen possesses 17 femoral pores separated by a diastema of eight non-pored scales. The collection locality for all these specimens is Malabar which is a large area and it will be difficult to narrow down to a specific locality based on available data. Fresh specimens from Kerala should be examined to ascertain if they are indeed conspecific with H. graniticolus, H. maculatus or represent yet another undescribed species. Records of the species from Karwar, Karnataka are conspecific with H. prashadi (Harshil Patel, personal communication). Specimens from Odisha and Telangana must be examined to ascertain if they are conspecific with the new species or represent yet another undescribed species.
Discovery of yet another species of a large gecko brings to light the poor nature of reptile documentation and increases the number of Hemidactylus in India to 31 [1]. This is the second new species of Hemidactylus to be described from Chhattisgarh after H. yajurvedi [16]. The Eastern Ghats have long been neglected with regards to its biodiversity as are other hill ranges in the country, but recent explorations have resulted in discovery on interesting reptiles from across India [3,17]. Dedicated surveys across unexplored areas of India, especially outside of biodiversity hotspots, where most of funding and efforts are diverted, will certainly yield more undescribed species of reptiles. Further, a detailed revision of most species that are considered widespread is necessary to elucidate cryptic species which are presently grouped under a single species. Documentation of cryptic species will greatly enhance our understanding of the reptilian diversity of India.
Funding details
Research work conducted by Z.M. was sponsored by Tulika Kedia through the Singinawa Conservation Foundation.
Geolocation information
India.
Acknowledgements
Research work conducted by Z.M. was sponsored by Tulika Kedia through the Singinawa Conservation Foundation. Fieldwork in Chhattisgarh would not have been possible without the support of Sanctuary Asia, Tourism Department of Chhattisgarh, and the Forest department. Special thanks to the PCCF/Wildlife, Forest Department of Chhattisgarh for issuing necessary permits to carry out surveys across the state (Permit letter No. 71/16/4454 & 71/16/4487). Harshil Patel (Gujarat) provided valuable inputs on the manuscript and data from which the manuscript greatly benefitted. Atish Gawai and Deepak C.K. kindly permitted us to use their images. Rahul Khot and Saunak Pal (Bombay National History Society) helped with specimen registration and data of specimens in the BNHS collection. The Herpetology department of California Academy of Sciences is thanked for sharing images of specimens in their collection. Z.M. visited the Natural History Museum, London through a travel grant from the Newby Trust Limited. David Gower and Patrick Campbell (Natural History Museum, London) are acknowledged for hosting Z.M. and granting access to specimens in their collection. The Director of ZSI and Kaushik Deuti granted access to specimens in the collection of ZSI.