

1 Introduction
The taxonomic and phylogenetic studies of terrestrial and freshwater invertebrates inhabiting the North Atlantic islands and archipelagoes (Faeroes, Iceland, Greenland, Svalbard, and others) are of great interest from the biogeographic and paleogeographic points of view [1–4]. It is supposed that the island fauna of Atlantic (including islands of Macaronesia) is rather species-poor, and the percentage of endemic taxa is very low. Probably, the vast majority of invertebrate species have arrived there either from the Americas or from Europe [3,5,6] as the result of long-distance dispersal events. However, the conclusions concerning taxonomic status, phylogenetic affinity, and endemism of the North Atlantic island invertebrates have often been based on the morphology-based systematics only. Given the fact that such phenomena as cryptic speciation, morphological convergence and parallel phenotypic evolution are widespread among invertebrate phyla, it is reasonable to suspect that many biogeographic inferences available from the literature may be at least partly biased. The molecular taxonomic studies of endemic taxa of the North Atlantic are still rare [7–9].
Today, the integrative approach to taxonomy and phylogeny based on joint use of the genetic, morphological, biogeographic and paleogeographic data is acknowledged as giving the most robust results in comparison with purely morphological or exclusively molecular studies. Here, we apply this approach to re-assess the taxonomic position and phylogenetic relationships of three species of the family Lymnaeidae Rafinesque, 1815 (aquatic pulmonate gastropods), which are considered as endemics to Greenland [10,11].
The scientific exploration of the fauna of Greenland, a remote Arctic island, started at the end of the eighteenth century, when the first papers on this subject were published, including Otto Fabricius’ prominent monograph Fauna Groenlandica [12]. As the freshwater Mollusca of Greenland are concerned, their study begins with a quite curious report of “Die breite Mahlermuschel aus Grönland” (i.e. “The broad Unio mussel from Greenland”) published by Schröter [13: 181, pl. IX, fig. 1], a German conchologist. Ironically, the specimen described and illustrated by Schröter originated from the tropics (Tranquebar Island in India) instead of the Arctic, and no species of large freshwater mussels inhabit Greenland [14].
A century later, Westerlund [15] was already able to give a more or less complete zoogeographical and ecological characterization of the Greenland continental malacofauna. In particular, he noted that the fauna includes 13 species of pulmonates, one species of fingernail clams [Pisidium steenbuchi (Møller, 1842)], and no representative of prosobranch snails. Westerlund [15] stated that the land snails of Greenland are mainly of European origin, whereas the aquatic ones demonstrate clear affinity to the Nearctic taxa.
The most prominent and diverse group of aquatic snails in Greenland are lymnaeids (family Lymnaeidae Rafinesque, 1815). Fabricius [12] did not describe any species of freshwater Mollusca, and thus Beck [16] became the first author to propose the scientific names for species of Lymnaeidae from Greenland: Limnaea vahlii Beck, 1837, L. pingelii Beck, 1837, and L. holboellii Beck, 1837. Since the three species were not described properly by Beck [16], most authors consider these names as nomina nuda [17,18]. Møller [19] published the diagnoses of the three species and thereby made the names available under his authorship.
Mørch [14,20] listed nearly 10 nominal taxa of the Greenland Lymnaeidae of species or variety rank. However, he assumed that the actual diversity of the pond snails may be lower, as differences between some taxa are very slight and they are probably synonyms. In 1868, Mørch [14] accepted two species, L. vahlii (with three varieties), and L. holboelli. Westerlund [15] added one more species, L. wormskioldi Mørch, 1857 to this list.
Dall [21] accepted L. vahlii and L. holboelli as two distinct species living in Greenland and went further regarding L. vahlii as a species of rather wide distribution in North America. According to Dall, the range of L. vahlii covers Greenland to Alaska and south to Minnesota and British Columbia. However, Dall [21] had some doubts about validity of L. holboelli and suggested that it may represent ‘merely a depauperate variety’ of L. vahlii.
Baker [22, p. 371] also considered L. vahlii to be a widely distributed species occurring in western Greenland to Alaska, “extending northward from about the 58th degree of north latitude”. Two subspecies of L. vahlii were distinguished by Baker [22]: L. v. pingelii and L. v. arctica (Lea, 1864). However, the anatomical structure of the Greenland Lymnaeidae was still not described, and both Dall [21] and Baker [22] based their conclusions on the examination of shells only.
The first description of the internal anatomy of the Greenland lymnaeids was published by Soós & Schlesch [23]; however, the authors neither discussed the taxonomical, nor the phylogenetic implications of their results. The anatomical study of the Greenland Lymnaea carried out by Hubendick [17] led him to the conclusion that there is a single species of this genus, Lymnaea vahlii, distinct from the North American congeners. Earlier, Mozley [24] provided additional evidence of the species status of L. vahlii based on a biometrical study of shell variation. Hubendick [17] rejected all reports of L. vahlii from localities situated beyond Greenland as being the result of probable confusion with L. arctica and other Nearctic species.
Nevertheless, some authors still accept more than one species of Lymnaeidae in Greenland. Kruglov [11] believes that there are three or four separate species characterized by a relatively short penis sheath. Kruglov and Starobogatov [10, p. 67] separated them into a subgenus of its own, with the following diagnosis: “Whorls weakly or moderately inflated. Cusps of outer lateral teeth unequal. Penis sheath not narrows to praeputium; sarcobellum well developed”.
All above-mentioned papers were based on the examination of shell structure and radular and reproductive anatomy of the snails. However, the phylogenetic value of these characters may be limited due to excessive intra- and interspecific variation, as well as convergence.
Hubendick [17] reported that anatomically, L. vahlii from Greenland is very similar to the South American species Lymnaea (Pectinidens) diaphana King et Broderip, 1832, but explained this fact by parallel evolution. Mozley [24, p. 187] believed that “a casual examination of these Greenland freshwater shells gives little insight into their mutual affinities and degree of relationship to species in other parts of the Sub-Arctic Region”. A recent molecular phylogenetic study has shown that L. diaphana is phylogenetically related to the Nearctic species of the genus Ladislavella B. Dybowski, 1913 [such as L. catascopium (Say, 1817)], but still deserves to be separated in a special genus, Pectinidens Pilsbry, 1911, interpreted as “an archaic relict form derived from the oldest North American stagnicoline ancestors” [25].
Thus, neither the species content, nor the phylogenetic affinities of the West Greenland lymnaeid snails are currently wholly understood. In this study, we address the question how many valid taxa of Lymnaeidae live in West Greenland and what are their phylogenetic relationships with other lymnaeids. The problem of the origin of the Greenland freshwater malacofauna has also been addressed.
2 Material and methods
2.1 Snail collection and taxonomic determination
The primary material for this study was collected between 5 and 21 July 2014 by two of the authors (M.V. and I.N.) in several localities situated on the western coast of Greenland. The sampled habitats were concentrated in three areas (Fig. 1):
- • vicinities of Kangerlussuak Settlement (approximately 67°00′N; 50°41′W). The lymnaeids were sampled from 11 waterbodies belonging to three different types (see below);
- • Nuuk Area. We inspected six waterbodies situated in Nuuk (the capital of Greenland) or in mountain ridges surrounding the town. The lymnaeids were found in a single waterbody – a small shallow lake lying in the center of Nuuk (64°11′15′′N; 51°43′14′′W);
- • vicinities of Narsarsuaq settlement. More than 10 waterbodies of various types were inspected. Lymnaeids were found only in few lakes situated near Quarsiarsuq settlement (61°08′09′′N; 45°32′17′′W). In other waterbodies snails were represented by the planorbid species Gyraulus arcticus (Møller, 1842) only, or were absent;
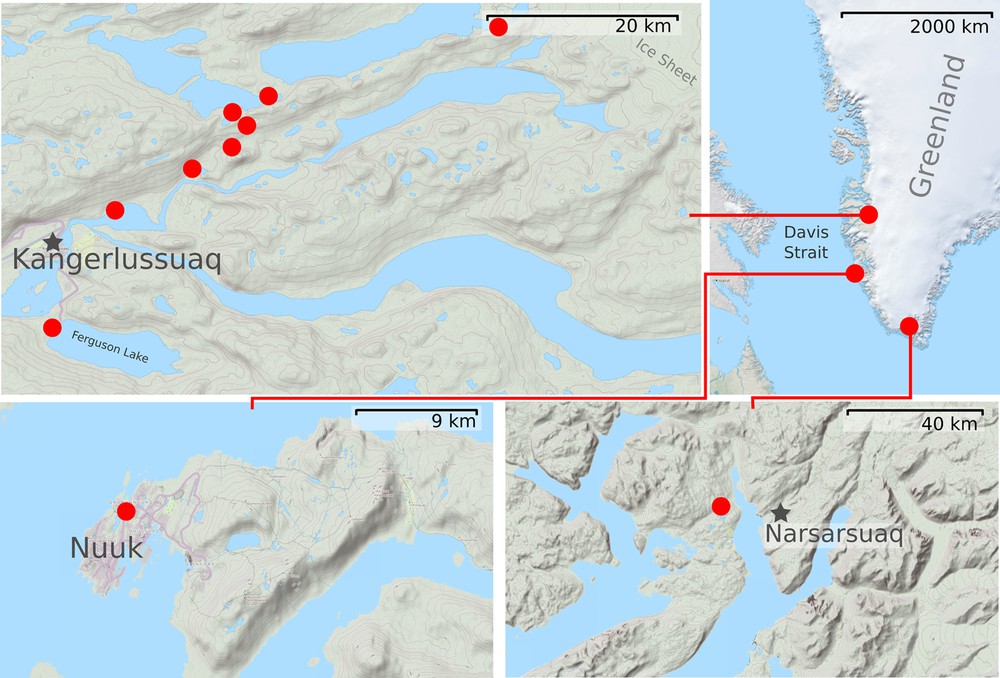
Map of Greenland, showing the sampling areas.
In the vicinities of Kangerlussuak, three types of waterbodies could be discerned (Fig. 2):
- • large mountain lakes with heavy wave action and stony substrate. Lake Tasersuatsiaq (Ferguson) is a typical representative of this group (see Fig. 2A);
- • small tundra lakes with soft bottom substrate and a narrow belt of vegetation. This is the most abundant type of lake in the area, most of them are nameless (see Fig. 2B);
- • floodplain pools. Very shallow, with no vegetation and presumably non-permanent (see Fig. 2C). We inspected several such pools situated in the floodplain of Qinnguata Kuussua River outflowing from the Russell Glacier.

Photos of some of the sampled waterbodies of Greenland. A. Tasersuatsiaq (Ferguson) Lake near Kangerlussuak. B. A nameless small lake, 5 km east from Kangerlussuak. C. A pool in the floodplain of Qinnguata Kuussua River, 1 km east from Kangerlussuak. D. A lake in Nuuk Town. Photos: M. Vinarski.
Snails were picked up by hand from the substrate in the shallow zones of lakes or from vegetation. Also, a sieve was used to collect mollusks from the bottom. The animals sampled were immediately put into plastic tubes filled with 96% ethanol. In total, more than 1500 specimens were collected (most of them were empty shells abundantly sampled for conchometric study). The bulk of the sample is kept in the collection of the Laboratory of Macroecology and Biogeography of Invertebrates, Saint-Petersburg State University, Russia. The shells and tissues of individuals used for genetic studies are in the collection of Senckenberg Natural History Collections Dresden (Museum of Zoology), Germany (SNSD hereafter). Some voucher specimens were given to the collections of the Zoological Institute of the Russian Academy of Sciences, Saint-Petersburg, and Vienna Museum of Natural History, Austria (NHMW hereafter).
The type series of three nominal Møller's [19] species of Greenland Lymnaeidae are kept in the Natural History Museum of Denmark (Copenhagen, ZMUC hereafter) and were examined by us prior to the field work in West Greenland. Several samples of Greenland lymnaeids kept in NHMW were also examined.
The type specimens of Limnaea vahlii Møller, 1842 are kept in ZMUC under the numbers GAS-106, 107, 108, 109, and 110. There are several hundreds of syntypes in ZMUC and six others are housed in the Stockholm Museum of Natural History, Sweden (No. SMNH 3842). One of the syntypes of L. vahlii was previously illustrated by Schiøtte and Warén [18, their fig. 7]. The type locality of L. vahlii is not known exactly. According to Schiøtte and Warén [18], the syntypes were collected in ponds of southwestern Greenland, without exact localisation of these waterbodies.
The type series of Limnaea holboelli Møller, 1842 is kept in ZMUC under the numbers GAS-54, 55, and 56, and includes 27 syntypes collected somewhere in West Greenland. One of the syntypes was illustrated by Schiøtte and Warén [18, their fig. 8].
The type series of Limnaea pingeli Møller, 1842 (ZMUC No. GAS-83) includes five syntypes. The type locality of this species is “a small pond between the mountains at Godthaab” [18, p. 12]. Godthaab is the former name for Nuuk (see above).
Shells of lymnaeids were measured following the standard scheme [11,26] by means of calipers of the ocular-micrometer of a stereoscopic microscope with an accuracy of 0.1 mm. Six measurements were taken from each shell: shell height and width, spire height, body whorl height, height and width of the aperture. The number of whorls of each shell was also counted. The measurements were then analyzed statistically by means of the multivariate statistical algorithms. Discriminant analysis and analysis of principal components (PCA) were realized with the STATISTICA 6.0 software (StatSoft Inc., USA).
The application of different multivariate techniques to the studies of variation in shells of freshwater gastropods is usually complicated by inevitable size differences among specimens in large samples. There are several ways how to set aside this factor and to analyze the “pure” diversity in shape [27]. In this study, we used PCA to extract several principal components, the first of which (PC1) is typically tightly correlated with absolute measurements of shells and thus is considered as the composite “size” variable [28,29]. Then, we carried out the discriminant analysis based on the values of PCs 2–4, which are presumably less correlated with absolute dimensions and reflect mainly differences in shape [29].
Dissections of snails were made following the standard procedure [11]. We examined the taxonomically significant parts of the male and female reproductive systems, including the copulatory apparatus, prostate, bursa copulatrix, and its duct. The index of the copulatory apparatus (ICA hereafter) was calculated for each dissected individual as the ratio between of the lengths of its praeputium and penis sheath (see Fig. 7C and [30] for details).

The copulatory organs (A–F) and fragments of the female reproductive system (G, H) of the Nearctic Ladislavella. A, B. L. vahlii. A nameless small lake, 5 km east from Kangerlussuak. C, D, G. L. vahlii. Tasersuatsiaq (Ferguson) Lake. E. L. vahlii. A lake in Nuuk Town. F, H. L. elodes. USA, Illinois, swampy area off Larue-Pine Hills Rd, Larue-Pine Hills National Landmark. Scale bars 1 mm. (C) shows the structural parts of the copulatory organ and the scheme of their measurements (pp–praeputium; ps–penis sheath; sd–spermiduct).
In total, 176 shells (including syntypes) were measured and 64 snails were dissected (Table 1). Unfortunately, many of the type specimens of Møller's taxa are damaged (broken shell apices) and thus were unavailable for measurement. The juvenile and subadult shells were excluded from morphometric analysis as well. Therefore, the overall number of type specimens used in this study is relatively low compared to the total number (see Table 1).
Samples of lymnaeids from the West Greenland used in morphological analyses.
| Taxon name, sampling site, collection date (in brackets–museum accession number) | No. of measured shells | No. of dissected snails |
| Limnaea vahlii, ca. 1840, West Greenland (ZMUC GAS-106). Syntypes | 4 | – |
| Limnaea vahlii, ca. 1840, West Greenland (ZMUC GAS-109). Syntypes | 3 | – |
| Limnaea vahlii, ca. 1840, Nep Sundet, West Greenland (ZMUC GAS-110). Syntypes | 10 | – |
| Limnaea holboelli, ca. 1840, West Greenland (ZMUC GAS-54). Syntypes | 5 | – |
| Limnaea pingelii, ca. 1840, a small pond between the mountains at Godthaab (ZMUC GAS-83). Syntypes | 2 | – |
| L. vahlii, 06.07.2014, a nameless small lake, 5 km E from Kangerlussuak | 37 | 10 |
| L. vahlii, 08.07.2014, Tasersuatsiaq (Ferguson) Lake near Kangerlussuak | 18 | 16 |
| L. vahlii, 09.07.2014, a nameless lake, 8 km E from Kangerlussuak | 47 | 10 |
| L. vahlii, 13.07.2014, a small lake in Nuuk Town | 11 | 12 |
| L. vahlii, 05.07.2014, a pool in the floodplain of the Quinnguata Kuussua River near Kangerlussak | 39 | 16 |
| In total | 176 | 64 |
2.2 Molecular techniques, sequence alignment, phylogenetic analyses, and species delimitation
For outgroup comparison in the molecular genetic analyses we included sequences of Palearctic specimens of the species Aplexa hypnorum (Linnaeus, 1758) from the freshwater gastropod family Physidae and from the lymnaeid species Galba truncatula (O.F. Müller, 1774), Lymnaea stagnalis (Linnaeus, 1758), L. taurica (Clessin, 1880), Stagnicola corvus (Gmelin, 1791), S. palustris (O.F. Müller, 1774) and S. fuscus (C. Pfeiffer, 1821). We included own sequences of Ladislavella terebra (Westerlund, 1885), L. elodes (Say, 1821), and L. catascopium in the in-group. In addition, we used ITS2 sequences of L. elodes, L. catascopium, L. terebra [= Catascopia occulta (Jackiewicz, 1959)] and L. emarginata (Say, 1821) available from GenBank.
Tissue samples taken from the foot were fixed in 100% ethanol or 100% isopropyl alcohol. They were registered in the tissue collection of the SNSD with both a new tissue voucher number and the collection number in the mollusk collection of SNSD and stored at –80 °C.
For the molecular genetic analyses, we obtained sequence data of the nuclear ITS2 spacer (partial, with 28S ribosomal RNA gene, partial sequence, 280 bp in A. hypnorum, up to 506 bp in L. taurica, the length of the ITS2 spacer varies within the genera and families) and a 329 bp fragment of the cyt-b gene as mitochondrial marker. For primers and protocols of DNA extraction, Polymerase Chain Reaction (PCR), purification of PCR products and DNA sequencing, see Schniebs et al. [31].
All specimens used in the molecular genetic study are listed in Appendix 1. All DNA-sequences have been placed in the European Nucleotide Archive (ENA, see http://www.ebi.ac.uk/ena/).
The alignment of the cyt-b gene dataset was performed using the ClustalW algorithm implemented in MEGA6 [32]. ITS2 sequences were also aligned using the ClustalW algorithm of MEGA6 [32], because of the lack of any analogues with known secondary structure in the ITS2 database [33]. To eliminate highly variable fragments from the ITS2 alignment, we used GBlocks v. 0.91b [34,35] using options for less stringent selection enabling inclusion of gap positions, smaller final blocks and less strict flanking positions. The resulting length of the ITS2 alignment was 468 bp (85% of the original 545 positions). Each alignment was collapsed into unique haplotypes (see Appendix 1) using an online FASTA sequence toolbox (FaBox 1.41; [36]). Absent sites were treated as missing data. A partition homogeneity test was applied in PAUP v. 4b10 to confirm the congruence of phylogenetic signals among sequence data sets [37,38]. This test reveals significant conflict (P = 0.01), and we therefore analyzed each gene data set separately. Each codon of the cyt-b gene was treated as a separate partition.
All phylogenetic analyses were performed at the San Diego Supercomputer Center via the CIPRES Science Gateway [39]. Maximum likelihood (ML) analyses were conducted using RAxML v. 8.2.6 HPC Black Box [40]. A unique GTR model was applied for each partition with corrections for gamma distribution. Nodal support values were estimated using an automatic, rapid bootstrapping algorithm according to the developer's recommendation [41], and the majority-rule consensus tree was constructed from the independent searches. Bayesian inference (BI) analyses were performed in MrBayes v. 3.2.6 [42]. The best models of sequence evolution for each partition based on the corrected Akaike Information Criterion (AICc) of MEGA6 [32] are presented in Table 2. Two runs, each with three heated (temperature = 0.1) and one cold Markov chain, were conducted for 30 million generations. Trees were sampled every 1000th generation. After completion of the Markov Chain Monte Carlo (MCMC) analysis, the first 15% of trees were discarded as burn-in (pre-convergence part), and the majority-rule consensus tree was calculated from the remaining trees. Convergence of the MCMC chains to a stationary distribution was checked visually based on the plotted posterior estimates using an MCMC trace analysis tool (Tracer v. 1.6; [43]). The effective sample size (ESS) value for each parameter was always recorded as > 10,000.
Models of sequence evolution for each partition based on corrected Akaike Information Criterion (AICc) of MEGA6 (Tamura et al., 2013) that were applied within a Bayesian inference framework.
| Partition | Model | Invariant | Gamma |
| cyt-b | |||
| 1st codon | HKY + G | n/a | 0.38 |
| 2nd codon | T92 + G | n/a | 3.35 |
| 3rd codon | GTR + G + I | 0.54 | 2.36 |
| ITS2 | GTR + G | n/a | 0.82 |
To delimit putative species-level units, we used a molecular approach based on the concept of Molecular Operational Taxonomic Units (MOTUs thereafter) [44,45]. MOTUs were separated using the Poisson Tree Processes (PTP) model to infer putative species boundaries on a phylogenetic input tree inferred from a ML analysis of the cyt-b haplotype sequences [46]. We used a Bayesian implementation of the PTP model for species delimitation through an online bPTP server (http://species.h-its.org/ptp) with 500,000 MCMC generations and 15% burn-in [46]. The outgroup haplotypes were removed from the input tree using an appropriate option of the server.
2.3 Statistical biogeographic modelling
To test the null hypothesis of equal evolutionary rate throughout the tree (a global molecular clock model), we performed the likelihood ratio test (LRT) [32,47] based on the consensus phylogenetic tree inferred from ML analyses of the cyt-b haplotypes. The null hypothesis was rejected at a 5% significance level (P = 0). Moreover, the external molecular clock rates appear to be hardly applicable to the Lymnaeidae, because of the possibility of ultrafast molecular evolution in several taxa [9]. The fossil-calibrated phylogenetic reconstructions for lymnaeid snails are still lacking. For biogeographic reconstructions, we therefore used an uncalibrated ultrametric tree of the cyt-b haplotype sequences, which was obtained using BEAST v. 1.8.4 with a lognormal relaxed clock algorithm and the Yule speciation process as the tree prior [48]. Calculations were performed at the San Diego Supercomputer Center through the CIPRES Science Gateway [39]. We specified the same substitution models to the three-codon positions as in the MrBayes analyses, but have used the simple HKY model instead GTR. Two replicate searches were conducted, each with 30 million generations. The log files were checked visually with Tracer v. 1.6 for an assessment of the convergence of the MCMC chains and the effective sample size of the parameters [43]. Almost all ESS values were recorded > 1000; the posterior distributions were similar to the prior distributions. The resulting tree files from two independent analyses were compiled with LogCombiner v. 1.8.4 [48]. The first 15% of trees were discarded as an appropriate burn-in (pre-convergence part). The maximum clade credibility tree was obtained using TreeAnnotator v. 1.8.4 [48]. We tested ancestral area patterns using three different approaches, i.e., Statistical Dispersal-Vicariance Analysis (S-DIVA), Dispersal-Extinction Cladogenesis (Lagrange configurator, DEC), and Statistical Dispersal-Extinction Cladogenesis (a nonparametric ‘Bayes–Lagrange’ approach to biogeographic inference, S-DEC) implemented in RASP v. 3.2 [49]. For an ancestral area reconstruction, we used the set of 18,002 uncalibrated trees that were combined from two runs of BEAST v. 1.8.4 with an appropriate burn-in (see above). As a condensed tree, we used the user-specified consensus tree, which was obtained based on this set of trees using TreeAnnotator v. 1.8.4 (see above). Only Ladislavella taxa were used in the analyses, all the other taxa were removed from input trees using the appropriate option of RASP v. 3.2. We coded three distribution areas as follows: Greenland (A), mainland North America (B), and Eurasia (C). The S-DIVA models were calculated with the following parameters: max areas = 3; allow reconstruction with max reconstructions = 100 and max reconstructions for final tree = 1000; allow extinctions. The DEC and S-DEC analyses were run with default settings and max areas = 3. In addition to the evaluations obtained from each analysis separately, we used the generalized results of all three modeling approaches that were combined using an algorithm implemented in RASP v. 3.2.
3 Results
3.1 Conchological examination of the type series
The variability of shell size and proportions among syntypes of Lymnaea vahlii proved to be very high. The shell shape varies from tall and slender turriculate shells to almost ovate specimens with short conical spires (Fig. 3A–D). Most syntypes are brown coloured, fragile, with thin shell walls, and with more or less prominent axial lines on the body whorl surface (see Fig. 3C and D). The aperture proportions are also very variable. Despite the obvious difference among the shells of syntypes, all of them more or less correspond to Møller's [19] original diagnosis, which, however, is very brief and vague.

Shells of three nominal species of Lymnaea described from Greenland. A–C. L. vahlii, syntypes. No. GAS-110. “Nep Sundet, West Greenland”. Collected ca. 1840 by Møller and Holbøll (ZMUC). D. L. vahlii, a syntype. No. GAS-106. “West Greenland”. From the original Møller's collection (ZMUC). E–G. L. holboelli, syntypes. No. GAS-55. “West Greenland”. Collected ca. 1840 by Møller or Holbøll (ZMUC). H. L. pingeli. “Greenland, ex coll. W. Blume” (NHMW). Scale bars: 2 mm.
The extent of conchological variation in the L. holboelli syntypes is comparable with that in the type series of L. vahlii. The shells are diverse in their size and especially in their shape (see Fig. 3E–G). It proved impossible to discriminate the two species based on their shell proportions. Møller [19] believed that L. holboelli shell has a more acute spire and a deeper suture compared to L. vahlii. We should note, though, that these differences appear not to be stable, especially if the overall extent of conchological variation within the type series of L. vahlii is considered. The conchological variables of the two species greatly overlap (Table 3). The two shells of the syntypes of L. pingelii are rather similar to the syntypes of the two above-mentioned species (see [18], their Fig. 9) and, most probably, rather represent an intraspecific form of L. vahlii.
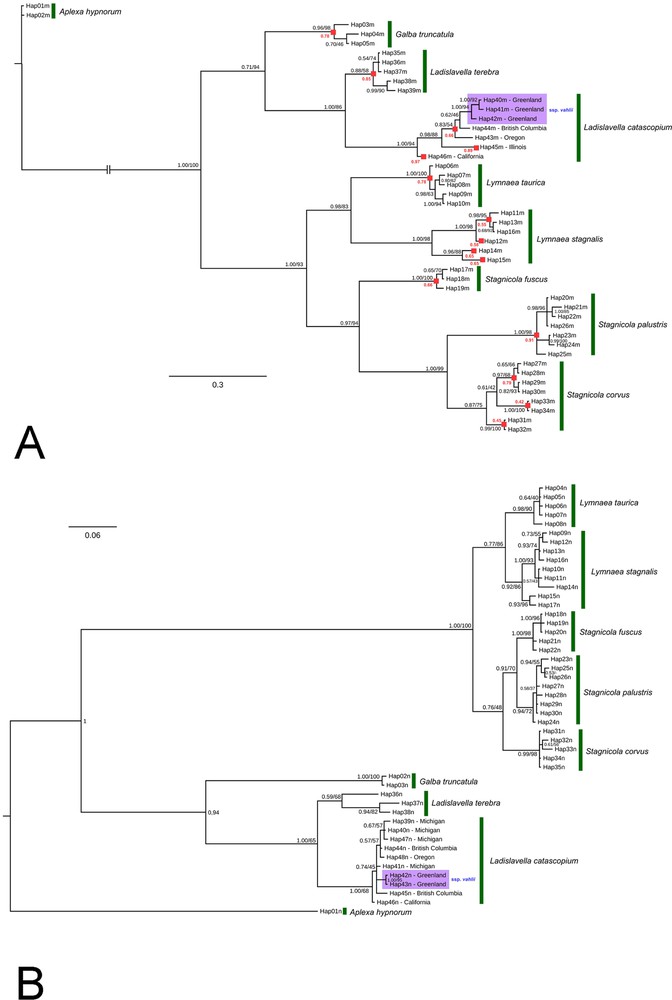
A. Fifty-percent majority-rule consensus phylogenetic tree of the Lymnaeidae recovered from Bayesian analysis of the cyt-b dataset with 44 in-group haplotypes. Two haplotypes of Aplexa hypnorum were used as an outgroup. Black numbers near branches are Bayesian posterior probabilities/ML support values. Red numbers near nodes are probabilities of species-level MOTUs (red squares) based on the highest Bayesian supported solution of the PTP species delimitation model. The violet filling indicates the Greenlandic subspecies of L. catascopium, which is supported well by the mitochondrial sequence data. B. Fifty-percent majority-rule consensus phylogenetic tree of the Lymnaeidae recovered from Bayesian analysis of the ITS2 dataset with 47 in-group haplotypes. A haplotype of Aplexa hypnorum was used as an outgroup. Black numbers near branches are Bayesian posterior probabilities/ML support values. The violet filling indicates the Greenlandic subspecies of L. catascopium, which is supported well by the nuclear sequence data.
3.2 Conchological variation in the Greenland Lymnaeidae collected in 2014
According to our field observations and shell measurements, the intrapopulation variation in the Greenland lymnaeids is very high, and its amplitude corresponds to the extent of shell variation found within the type series of L. vahlii. Nevertheless, one may note that the shells can be sorted into two more or less prominent groups differing in their size and proportions. The first group includes snails with tall and slender high-spired shells, resembling shells of species belonging to the Stagnicola elodes group sensu Burch [50], which are widely distributed in North America (compare Fig. 4A–C and D–E). This group proved to be 100% distinguishable by means of the discriminant analysis (Table 4). The shells of the second group are ovate-conical or almost ovoid, with short spires and strongly inflated body whorls. They look similar to shells of species of the Stagnicola emarginata/catascopium group sensu Burch [50], also of Nearctic origin (compare Fig. 4F–H and I–K). The two forms will be designated hereafter as the ‘narrow’ and ‘broad’, respectively. These can be characterized mainly by differences in two conchological ratios: shell width:shell height and body whorl height:shell height (see Table 2).

Shells of North American and Greenland representatives of Ladislavella. A, B. L. vahlii, ‘narrow’ form. A nameless small lake, 5 km east from Kangerlussuak. C. L. vahlii, ‘narrow’ form. A pool in the floodplain of Qinnguata Kuussua River, 1 km east from Kangerlussuak. D. L. elodes s. str. USA, “lagoon near Aberdeen, South Dakota” (NHMW). E. L. elodes s. str. USA, Michigan, Ann Arbor (MSAM) F–G. L. vahlii, ‘broad’ form. Tasersuatsiaq (Ferguson) Lake. H. L. vahlii, ‘broad’ form. A lake in Nuuk Town. I. L. catascopium. USA, Michigan, Crystal Lake (ZMUC). J–K. L. emarginata. USA, Niagara Falls (NHMW). Scale bars: 2 mm.
Morphological characteristics of the West Greenland lymnaeids (including syntypes).
| Character or ratio | Syntypes | Specimens collected in 2014 | ||||
| L. holboelli | L. pingelii | L. vahlii | Tasersuatsiaq Lake |
Floodplain of the Quinnguata Kuussua Rivera |
A lake, 8 km E from Kangerlussuak Settlementa |
|
| Shell height (SH), mm | 7.4–11.0 8.8 ± 1.4 |
8.9–9.6 | 7.8–11.4 9.9 ± 1.1 |
13.3–20.3 16.5 ± 1.7 |
16.6–23.6 19.3 ± 1.9 |
18.0–25.6 21.6 ± 1.7 |
| Shell width (SW), mm | 3.2–5.0 3.7 ± 0.7 |
4.6–4.8 | 3.4–5.9 5.0 ± 0.6 |
7.3–10.8 9.0 ± 0.8 |
7.8–10.9 8.9 ± 0.8 |
8.1–10.8 9.6 ± 0.6 |
| Body whorl height (BWH), mm | 5.2–7.9 6.1 ± 1.1 |
6.4–6.6 | 5.7–8.6 7.4 ± 0.8 |
10.0–14.9 12.7 ± 1.2 |
11.3–15.5 13.1 ± 1.1 |
12.5–17.6 15.3 ± 1.0 |
| Aperture height (AH), mm | 4.1–5.4 4.5 ± 0.5 |
4.5–4.8 | 4.0–5.6 4.9 ± 0.4 |
6.3–10.6 8.7 ± 1.0 |
7.0–10.4 8.5 ± 0.8 |
8.1–12.0 10.0 ± 0.7 |
| Aperture width (AW), mm | 2.2–3.4 2.5 ± 0.5 |
2.9 | 2.5–3.6 3.1 ± 0.3 |
5.0–6.9 5.8 ± 0.6 |
5.1–7.6 6.0 ± 0.6 |
5.2–7.4 6.4 ± 0.4 |
| SW/SH | 0.49–0.55 0.51 ± 0.03 |
0.50–0.52 | 0.40–0.58 0.50 ± 0.04 |
0.51–0.59 0.55 ± 0.02 |
0.42–0.51 0.46 ± 0.02 |
0.40–0.49 0.45 ± 0.02 |
| BWH/SH | 0.67–0.72 0.69 ± 0.02 |
0.67–0.74 | 0.65–0.80 0.75 ± 0.04 |
0.73–0.83 0.77 ± 0.03 |
0.65–0.71 0.68 ± 0.02 |
0.67–0.76 0.71 ± 0.02 |
| AH/SH | 0.40–0.45 0.42 ± 0.02 |
0.50–0.51 | 0.41–0.62 0.50 ± 0.06 |
0.40–0.57 0.53 ± 0.04 |
0.40–0.48 0.44 ± 0.02 |
0.41–0.52 0.46 ± 0.03 |
| AW/AH | 0.49–0.53 0.51 ± 0.07 |
0.60–0.64 | 0.58–0.72 0.64 ± 0.03 |
0.61–0.94 0.67 ± 0.07 |
0.61–0.85 0.71 ± 0.05 |
0.56–0.72 0.64 ± 0.04 |
a Above line: limits of variation (min–max); below line: mean values ± standard deviation (σ).
In the vicinities of Kangerlussuaq, snails of the ‘narrow’ form occur almost exclusively in small tundra lakes and floodplain pools, whereas specimens belonging to the ‘broad’ group were found in two large mountain lakes in vicinities of Kangerlussuak settlement: Tasersuatsiaq (Ferguson) and Saningassoq (only eight specimens were collected here). The possible topotypes of L. pingelii sampled in the city of Nuuk also belonged to the ‘broad’ form (see Fig. 4H), although their habitat was a small lake. However, this waterbody lies in a small depression between two mountain gorges, and its bottom was covered by large and little stones (see Fig. 2D). Thus, it is likely that the distribution of both morphotypes is not random, but rather depends on the type of waterbody, but our evidence is insufficient to establish this pattern with certainty.
We found a difference between ‘broad’ and ‘narrow’ forms in the absolute size of their shells. The ‘broad’ shells are smaller and this size difference is statistically significant (Student's t-test: t = 5.864; P = 0.00). Therefore, the direct comparison of the two forms is difficult, unless the size factor of variation is removed. This was achieved with the PCA analysis of conchological variables (see Material and methods). Examination of the axes’ loadings reveals that the first axis of PC1 (explaining 79.0% of the variation) is the ‘size’ variable, since all linear measurements tightly correlate with PC1 values (Table5). The values of the subsequent axes (PCs 2–4) correlate with the number of whorls, spire height, and aperture height; hence, these variables contribute most to the differences between ‘narrow’ and ‘broad’ conchological forms.
Discriminant analysis of shell variation in populations of Greenland Ladislavella from waterbodies of different types (before removing the size component of variation).
| Observed classificationa | % correct | Predicted classification | ||
| ‘Broad’ form (I) | ‘Narrow’ form (II) | ‘Narrow’ form (III) | ||
| ‘Broad’ form (I) | 100.0 | 30 | 0 | 0 |
| ‘Narrow’ form (II) | 90.6 | 1 | 77 | 7 |
| ‘Narrow’ form (III) | 77.5 | 0 | 9 | 31 |
| Total | 90.1 | 31 | 86 | 38 |
a In brackets, the type of waterbody is indicated (see Material and methods).
The discriminant analysis performed after setting aside the first axis still allowed us to distinguish between the ‘broad’ and ‘narrow’ specimens, although the overall exactness of discrimination dropped to 80.0% Table 6. The phenotypic distinctness is not complete due to the existence of ‘intermediate’ specimens. The ‘broad’ and ‘narrow’ individuals form a single ‘cloud’ of points in the multivariate space (Fig. 5) that indicates the absence of the full hiatus between them (see also Table 2). The exactness of the discrimination between ‘narrow’ snails living in floodplain pools and tundra lakes (waterbody types II and III) is rather low (for ‘narrow’ specimens from floodplain pools, it is equal to 57.5%).
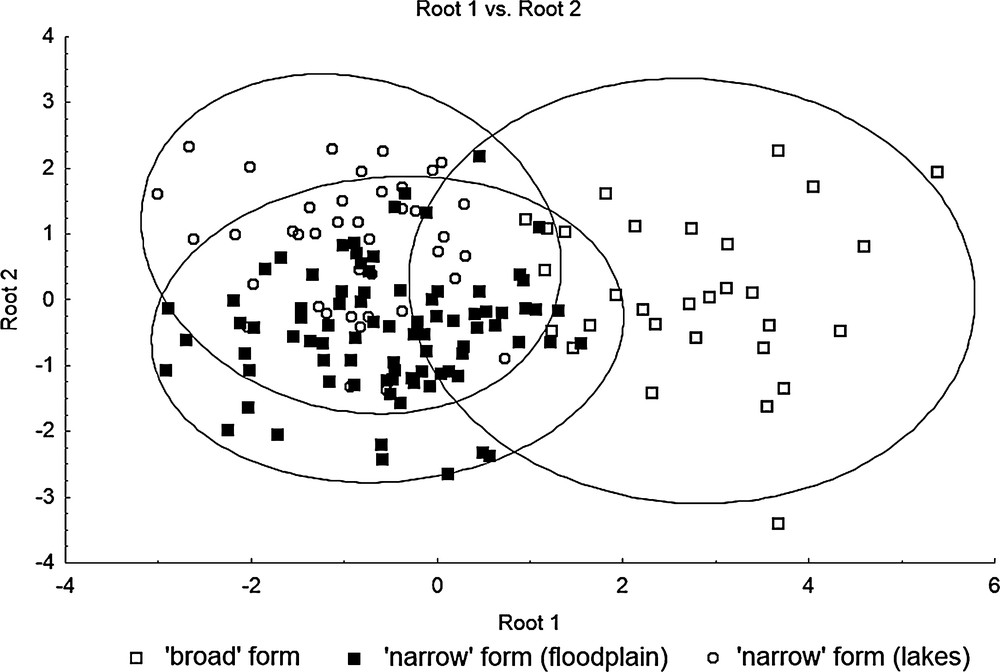
The canonical analysis of variation in conchological traits of the ‘broad’ and ‘narrow’ forms of L. vahlii from West Greenland.
The direct comparison between specimens collected in 2014 and the syntypes of Møller's nominal species was not possible due to large differences in shell size: the syntype shells are roughly two times smaller than the newly sampled individuals (see Table 2). Therefore, the procedure of setting aside the size component of variation was repeated on a united sample consisting of syntypes and non-syntypes. The analysis was successful in eliminating the size variation, since no differences between the type specimens and newly collected shells were found. All syntypes clustered together with the ‘narrow’ specimens sampled in 2014 (Fig. 6).
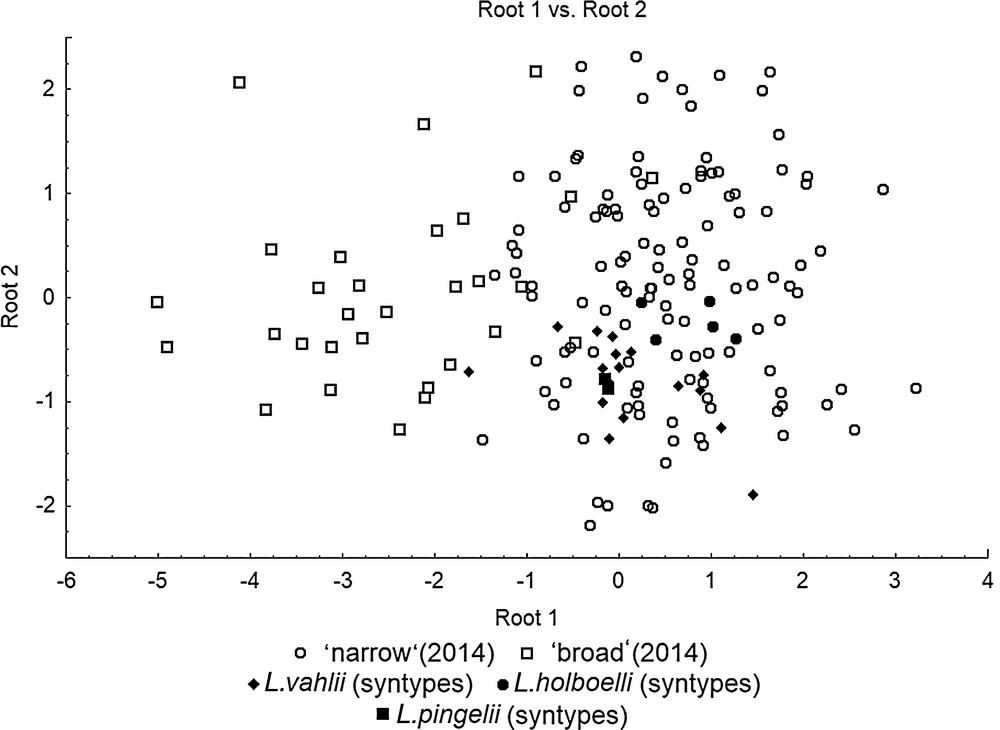
The canonical analysis of variation of lymnaeids from West Greenland, including syntypes of the species described by Møller [19] and the ‘broad’ and ‘narrow’ specimens sampled in 2014.
3.3 Anatomical variation
The structure of the copulatory organ in the Greenland lymnaeids is uniform across populations and waterbody types. All snails have an oblong, usually dark-pigmented praeputium with a more or less prominent distal end (near its border to the penis sheath). The penis sheath is much shorter and narrower than the praeputium, and strongly inflated at its distal end, forming a club-like structure (Fig. 7). The ratio between the lengths of the praeputium and the penis sheath (ICA) varies from 1.26 to 2.32. Though the mean ICA values vary across populations (Table 7), the extent of their variation corresponds to those observed in other lymnaeid species [30] and thus this trait is not effective for discrimination of populations and conchologically defined forms of L. vahlii.
Discriminant analysis of shell variation in populations of Greenland Ladislavella from waterbodies of different types (after the size factor was removed).
| Observed classificationa | % correct | Predicted classification | ||
| ‘Broad’ form (I) |
‘Narrow’ form (II) |
‘Narrow’ form (III) |
||
| ‘Broad form’ (I) |
83.3 | 25 | 5 | 0 |
| ‘Narrow form’ (II) |
89.4 | 1 | 76 | 8 |
| ‘Narrow form’ (III) |
57.5 | 0 | 17 | 23 |
| Total | 80.0 | 26 | 98 | 31 |
a In brackets, the type of waterbody is indicated (see Material and methods).
The bursa (receptaculum seminis) is short and compact, its duct is rather long and very narrow (see Fig. 7G).
Though we had no large samples of other Nearctic species of Ladislavella to carry out a comparative analysis, the examination of the internal structure of several individuals of L. elodes from USA waterbodies revealed no substantial differences in the structure of the copulatory organs and the female reproductive organs (see Fig. 7).
The mantle pigmentation of the Greenland Ladislavella proved to be variable as well (Fig. 8). Individual snails differ primarily in the size of the black patches and in the pattern of their distribution over the mantle surface. The largest black patch was found in a specimen from Lake Tasersuatsiaq, but, as it will be shown below, this difference does not correlate with genetic distance, making it unsuitable as a basis for any phylogenetic or taxonomical inferences.
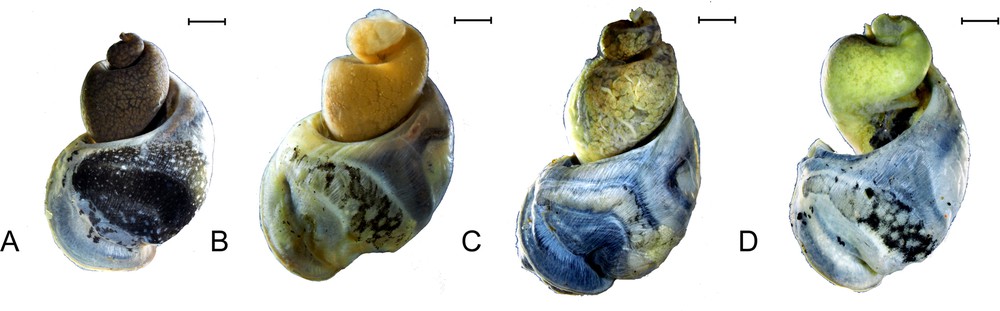
Mantle pigmentation in the West Greenland Ladislavella. A. Tasersuatsiaq (Ferguson) Lake. B. A lake near Quarsiarsuq. C. A nameless small lake, 5 km east from Kangerlussuak. D. A lake in Nuuk Town. Scale bars 1 mm.
3.4 Molecular phylogeny
The hypotheses for the phylogenetic relationships of Ladislavella vahlii recovered from Bayesian analysis of cyt-b and ITS2 sequences are represented in Fig. 9. Both trees demonstrate a similar topology, with two large groups of lymnaeids found: the first one includes taxa of supposedly Neartic origin (Galba, Ladislavella), whereas the second cluster embraces two Old World genera, Lymnaea and Stagnicola. This dichotomy has full Bayesian posterior probabilities/ML support in both cases.
The results of the species delimitation analysis indicate that the Ladislavella catascopium s. l. clade contains three MOTUs, each of which may represent a separate species-level cryptic taxon (see Fig. 9A). The most divergent haplotypes are from California (46m) and from Illinois (45m), whereas the other haplotypes belong to a single MOTU corresponding to Ladislavella catascopium s. str. (Nos. 40m–44m). The Greenland lymnaeids formed a separate cluster within Ladislavella catascopium clade in both trees. Three mitochondrial haplotypes unique for Greenland were recovered during this study (haplotypes 40m, 41m, and 42m in Fig. 9A). Two specimens of L. catascopium s. str. collected in British Columbia shared the same haplotype (44m), which proved to be the most similar genetically to the Greenland snails. The haplotypes 43m, 45m, and 46m were found in snails from the United States, provisionally determined as L. emarginata (see Appendix 1).
Two unique nuclear haplotypes (42n and 43n) were found in the Greenland lymnaeids. These haplotypes have not been revealed in the mainland populations of L. catascopium s.l. (see Fig. 9B). Seven other nuclear haplotypes (Nos. 39-41 and 44-47, see Fig. 9B) are distributed throughout North America, from British Columbia to California. Their distribution, however, is not spatially patterned: the snails sampled in Michigan do not constitute a separate cluster of the cladogram similar to that formed by the Greenland individuals. The same was characteristic of lymnaeids originated from British Columbia (see Fig. 9B).
The ‘broad’ and ‘narrow’ conchological forms of L. vahlii outlined above proved to be genetically indistinguishable. For example, specimens from Lake Ferguson (belonging to the ‘broad’ form) shared identical haplotypes (40m and 42n) with snails collected from a nameless small lake near Kangerlussuak, which were classified as ‘narrow’ (see Fig. 9, Appendix 1). On the other hand, the distribution of the haplotypes among specimens of the Greenland Ladislavella seems to be governed partly by geographic distances separating sampled areas. The majority of the Greenland haplotypes recovered during this study appeared in a single area and were not found elsewhere. The haplotypes 40m and 42n are characteristic of the Kangerlussuak area, whereas the haplotype 41m was found in Nuuk, and the haplotype 43m – in vicinities of Narsarsuaq settlement. The only exception is the haplotype 43n, whose bearers were collected in all three sampling areas (see Appendix 1). Anyway, we could not find two separate clades that would correspond to the ‘narrow’ and ‘broad’ groups specimens in both our trees.
The existence of a set of haplotypes (both mitochondrial and nuclear) unique for Greenland as well as its isolated position on phylogenetic trees allowed us to consider the Greenland samples as belonging to a separate subspecies indicated in the trees (see Fig. 9A and B) as Ladislavella catascopium vahlii.
It is interesting also to note that all three alleged species of the L. catasopium group living in the North American mainland (i.e. L. catascopium s. str., L. elodes, and L. emarginata) were characterized by their own set of unique haplotypes, not found in congenerics. Thus, L. catascopium s. str. in our study possess the haplotypes 44m, 39n, 44n and 45n; L. emarginata has two own nuclear haplotypes (40n and 41n), whereas L. elodes demonstrates the widest array of haplotypes unique for this taxon, including three mitochondrial (43m, 45m, and 46m) and three nuclear (46n, 47n, 48n) ones.
3.5 Biogeography
The combined results of our biogeographic modelling (Fig. 10; Table 8) suggest the Nearctic origin of the L. catascopium clade via a vicariance event separating the former continuous Holarctic range of the L. catascopium–L. terebra MRCA (probability = 0.98). The further radiation of L. catascopium s.l. occurred in North America (probability = 0.95). The origin of the Greenlandic subspecies most likely caused by a relatively recent vicariance event, with subsequent radiation within the island (probability of both events = 1.00).
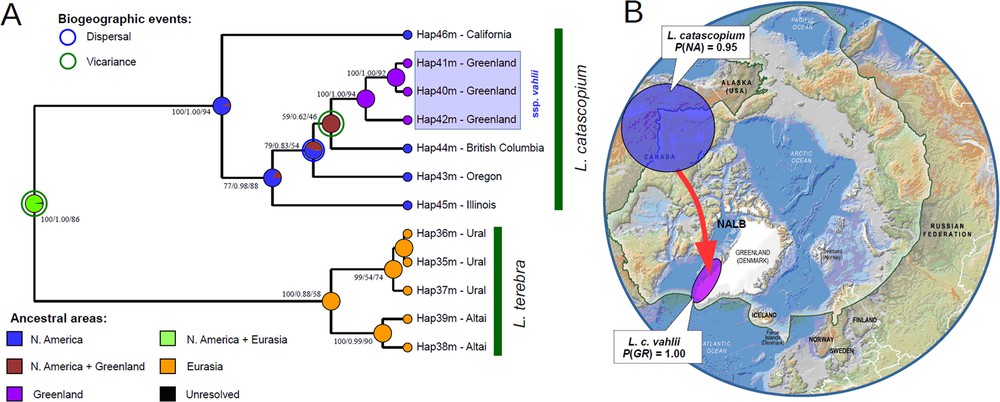
Evolutionary biogeography of the Ladislavella clade. A. Uncalibrated Bayesian phylogeny based on the mitochondrial cyt-b gene fragment with the combined results of ancestral area reconstruction analyses. Black numbers near branches indicate Bayesian posterior probabilities (BEAST)/Bayesian posterior probabilities (MrBayes)/ML bootstrap support values (RAxML). B. Simplified summary of origin of the Greenlandic subspecies L. catascopium vahlii. The red arrow shows the putative expansion event via the western part of the North Atlantic Land Bridge (NALB) obtained from the statistical biogeographic models (see Table 8 for details). Color circles indicate the putative places of origin of the entire species (blue) and the Greenlandic subspecies (violet). The P-values indicate the probability of origin of a clade/subclade within the area: Greenland (GR) and mainland North America (NA). The green boundary indicates the Arctic frontier. The free source of the map: Topographical map of the Arctic, Arctic Council, CAFF Map No. 14, http://library.arcticportal.org/id/eprint/1336.
Evolutionary biogeography of Ladislavella based on the ancestral area reconstruction analyses under three different modeling approaches.
| Clades | The most likely ancestral area | Probability of the most likely ancestral area | |||
| S-DIVA | DEC | S-DEC | Combined results | ||
| L. catascopium + L. terebra clade | North America + Eurasia (vicariance event) | 1.00 | 1.00 | 0.94 | 0.98 |
| L. catascopium clade | North America (intra-area radiation) | 1.00 | 1.00 | 0.84 | 0.95 |
| L. catascopium vahlii + L. catascopium ssp. subclade | Greenland + North America (vicariance event) | 1.00 | 1.00 | 0.99 | 1.00 |
| L. catascopium vahlii | Greenland (intra-area radiation) | 1.00 | 1.00 | 1.00 | 1.00 |
| L. terebra | Eurasia (intra-area radiation) | 1.00 | 1.00 | 1.00 | 1.00 |
4 Discussion
The morphological and molecular data presented above show that there is a single taxon of Lymnaeidae in West Greenland waterbodies characterized by prominent conchological variability that is not accompanied by substantial anatomical differences. Internal groups within the studied sample of the Greenland snails cannot be delineated, except for two phenotypical entities that we call the ‘broad’ and ‘narrow’ forms. These forms, however, appear to be of ecophenotypical origin, and their distribution in Greenland is likely predicted by the type of habitat. The short-spired shells with relatively large body whorls and wide apertures living in mountain lakes probably arise as an adaptation to heavy wave action. Similar intraspecific “lake” forms are known within the Nearctic species L. catascopium [51]. We did not detect genetic differences between ‘broad’ and ‘narrow’ individuals, and snails belonging to two different morphotypes may share the same haplotype. The two forms thus merely represent intraspecific variations connected by phenotypically intermediate specimens.
The three nominal species of Lymnaeidae with type localities situated in Greenland that were described by Møller [19] prove to be indistinguishable from each other. As early as 1868, Otto Mørch also stated that he could not see “any very striking difference between the Greenland forms and [North American lymnaeids such as] L. elodes, Say, L. expansa, Haldem., or L. decidiosa, Say” [14, p. 88]. The results of our study allow us to agree with Mørch. The only difference between the Greenland and North American pond snails mentioned by this author was the difference in the relative thickness of the shell wall: the Greenland lymnaeids have much thinner shells than the mainland ones [14]. We also observed this, but such a difference is clearly not enough to substantiate the species status of the Greenland pond snails.
The very close relationships, if not full identity, between the North American and Greenland lymnaeids were reported also by other authors studying conchological traits of these mollusks [21,22,24].
From the genetic point of view, the West Greenland lymnaeids determined as Lymnaea vahlii, L. pingelii, and L. holboellii are almost indistinguishable from widely distributed in North America species Ladislavella catascopium, L. elodes, and L. emarginata. This genetic similarity may reflect very recent migration events that led to colonization of the island by snails from the North American mainland. These events could not take place until 11 kyr B.P., when the Ice Age ended abruptly in the North Atlantic [3]. Before this, the whole of Greenland was covered with an ice sheet, so it was completely insuitable for freshwater snails.
Given the fact that Greenland is now well isolated from the North American mainland, it would be reasonable to consider the Greenland populations as a subspecies of L. catascopium s. lato, considering the anatomical differences that Hubendick [17] reported to separate this species from other North American stagnicolines. Conchologically, the Greenland snails differ from the mainland ones by much thinner shell walls and smaller size. The penis sheath in L. elodes is a little narrower than the praeputium width in the Greenland Ladislavella (compare Fig. 7F with A–E).
Such a level of genetic and morphological difference is roughly comparable to the difference between other subspecies pairs in Lymnaeidae studied genetically, for example, between Lymnaea taurica (Clessin, 1880) and L. t. kazakensis Mozley, 1934 [52].
The extent of intraspecific variation in shell characters in the Greenland pond snails is relatively high (see Electronic Appendix 2), but nevertheless substantially lower than variation in their mainland counterparts. For example, the shell shape in L. emarginata (a synonym of L. catascopia according to [53–55]) in North America may vary from typical “stagnicoline” to Radix-shaped, with very short conical spires and enormously inflated body whorls and very wide apertures [17,50,51]. In the past, such Radix-like stagnicolines of North America were sometimes even described as separate species (for example, Limnaea ampla Mighels, 1843, non Hartmann, 1821; Limnaea angulata Sowerby, 1872).
No such conchological forms are known to occur in Greenland, where the snails retain more or less uniform stagnicoline-shaped shells. Possibly, this indicates that the genetic diversity in the Greenland populations is low compared to that in the mainland ones.
It is known that the Arctic landscapes underwent very dramatic changes in the Pleistocene caused by cyclic fluctuations of alternating long-term periods of cooling and warming. Two main hypotheses have been advanced to explain the origin of contemporary Arctic biota [2]. The “Nunatak hypothesis” assumes that the core of the Arctic biota is formed basically by cold-adapted species that survived the glaciation in refugia. According to the alternative explanation (the “tabula rasa” hypothesis), the Arctic fauna and flora consists exclusively (or almost exclusively) of migrants that colonized this region during the recent postglacial period. The latter hypothesis appears to be more realistic for Greenland continental biota because the island was almost completely covered by an ice sheet during the last glacial maximum (ca. 18 kyr ago; [3]). Apparently, only a small proportion of species was able to survive in refugia. The presence of a separate monophyletic lineage of Ladislavella catascopium in Greenland (see Fig. 10) fits well with the pattern that was discovered in Icelandic populations of another lymnaeid snail, Radix balthica. They belong to the unique Icelandic lineage, which originated via rapid microgeographic radiation of a mid-Holocene founder population under an elevated mutation rate [9]. In contrast, the Icelandic lineage of Radix balthica differs by exceptionally high haplotype diversity that was most likely caused by strong geothermal influence [9,56]. The results of our study are in accordance with the “tabula rasa” hypothesis, since the genetic data suggest that the Western Greenland lymnaeids have only recently arrived in this area from North America. Similar scenarios were proposed for some species of freshwater arthropods and plants of Greenland [2,3].
There is no direct evidence of land bridges connecting the North Atlantic Islands during the last 20 Ma [3]; therefore, we have to look for possible mechanisms of transoceanic dispersal of snails. Three main modes of distribution are able to explain the colonization of the West Greenland Coast by North American invertebrates [1]: by wind, by sea currents (including drifting ice and wood) and zoochory (transportation by birds). The first of these appears very unlikely in the case of lymnaeids, whereas the two latter ones may theoretically be proposed as a real mechanism of freshwater snail migration from mainland to Greenland. The long-distance dispersal events have often been invoked in explanation of the presence of pulmonate snails on remote oceanic islands as well as the mechanisms of transcontinental invasions of exotic species [6,57,58].
The taxonomic implications of our results are as follows. First of all, we need to lump all the diversity of nominal taxa described from Greenland in the past into a single taxon of subspecies rank (see above). In this case, Lymnaea pingelii, L. holboelli, and L. wormskioldi become the full junior synonyms of Ladislavella catascopium vahlii comb.n.
The question of the taxonomic relationships between the mainland Nearctic species L. catascopium s.str. and its congenerics (L. elodes and L. emarginata) in the light of the data obtained during this work remains unresolved. Some authors, especially those followed the strictly morphological approach to species delimitation accepted the three species as separate taxa closely related to each other [17,50; but see 51]. This opinion was accepted by the authors of a recent checklist of the North American freshwater Gastropoda [59]. However, since Remigio and Blair [53] and Bargues et al. [54], the most prevalent idea was to regard L. elodes and L. emarginata as full synonyms of L. catasopium s. lato [55]. Hubendick [17, p. 132] himself cautiously noted that anatomical differences between L. catascopium and L. emarginata are so slight that “there are no features distinctly separating emarginata from catascopium in either the copulatory organs… or the radula”. Having examined anatomically a plethora of North American stagnicoline species, Walter [51] advanced the hypothesis of conspecificity of L. catascopium, L. emarginata, and L. elodes.
The data presented above indicate that the three alleged species of the L. catascopium group possess each its own group of unique haplotypes (both mitochondrial and nuclear). It may mean that there is a cryptic genetic diversity within this species complex that merits a further study. The sample of the mainland specimens of Ladislavella analyzed by us was too small to propose a definitive decision.
5 Conclusions
Our data show that West Greenland is inhabited by the North American Lymnaeid species Ladislavella catascopium. A low degree of morphological and genetic divergence between the Greenland and North American populations indicate the relatively recent island colonization by the pond snails. We assume the Nearctic Ladislavella (= subgenus Walterlymnaea Starobogatov et Budnikova, 1976; see [60] for details) to include a single widely distributed species, L. catascopium s. lato, with two subspecies: L. c. catascopium (inhabiting the North American mainland) and L. c. vahlii (endemic to West Greenland). A third subspecies, L. c. tumrokensis (Kruglov et Starobogatov, 1985) was recently identified by Vinarski et al. [61] in the Kamchatka Peninsula, and represents a group of populations of this species established in northeastern Eurasia. However, the existence of several nominal species of the mainland Ladislavella characterized by unique sets of haplotypes indicated that there might be more than one species of this genus in the waterbodies of Canada and of the United States.
Most likely, L. catascopium s. lato evolved on the North American mainland, where the highest level of phenotypical and, possibly, genetic diversity is observed. Subsequently, individuals are postulated to have dispersed eastward to Greenland and westward to Beringia, developing populations described here as two additional, geographically isolated subspecies (see also [61]).
Disclosure of interest
The authors have not supplied their declaration of competing interest.
Acknowledgements
We would like to express our thanks to Prof. Dr. Uwe Fritz (SNSD) for the financial support for the greatest part of the molecular analyses, Anke Müller (SNSD) for some sequences and the instruction of K.S. to lab work as well as Dr. Dilian Georgiev (University of Plovdiv), Michael Korn (University of Konstanz, Limnological Institute), Dr. André Reimann (SNSD), Dr. Michail O. Son (Odessa Branch Institute of Biology of the Southern Seas), Dr. Vitaliy Anistratenko (Institute of Zoology NAS of Ukraine, Kiev), Dr. Alfried V. Karimov (Omsk State Pedagogical University), Dmitry M. Palatov (Moscow State University), Dr. Michael L. Zettler (Leibnitz Institute for Baltic Sea Research), Reinhard Diercking (Hamburg), Uwe Jueg (Ludwigslust), Dr. Ira Richling (State Museum of Natural History Stuttgart), Andrea Pohl (Dresden), Dr. Nicole Schröder-Rogalla (Munich) and Christa Schniebs (Oelsnitz) for providing materials of European and Siberian snails. Special thanks are given to Dr. Stephanie Clark (Invertebrate Identification, USA) for sending us materials from the Nearctic waterbodies.
We are also deeply obliged to the museum curators whose help was invaluable to us: we thank Mag. A. Eschner (NHMW), Dr. T. Schiøtte (ZMUC), and Dr. A. Warén (SMNH) for their great help during work with collections.
The work of M.V., I.B and I.N. is supported financially by grants of the Russian Fund for Basic Research (projects No. 14-04-01236 and No. 16-05-00854), the President of the Russia Grant Council (No. MD-7660.2016.5 and No. MD-2394.2017.4), and the Russian Ministry of Education and Science (No. 6.1352.2017/4.6 and No. 6.2343.2017/PCh).