

1 Introduction
The state of Colima, situated in the western Pacific coastline of Mexico (Fig. 1), is one of the most biodiverse states despite its small size with only 5543 km2 representing 0.3% of the Mexican territory. Colima lies in an area where the Nearctic and Neotropical biogeographical realms meet; as a result, mixed fauna with local endemism is expected to occur in this region. Furthermore, the topology of the region is complex, with a steep altitudinal gradient going from the sea level to abrupt volcanic elevations above 4000 m in altitude [1]. Tropical wet climates and the interaction of local abiotic factors support a rich flora. Fourteen different floristic communities are recognized within this small state [2].
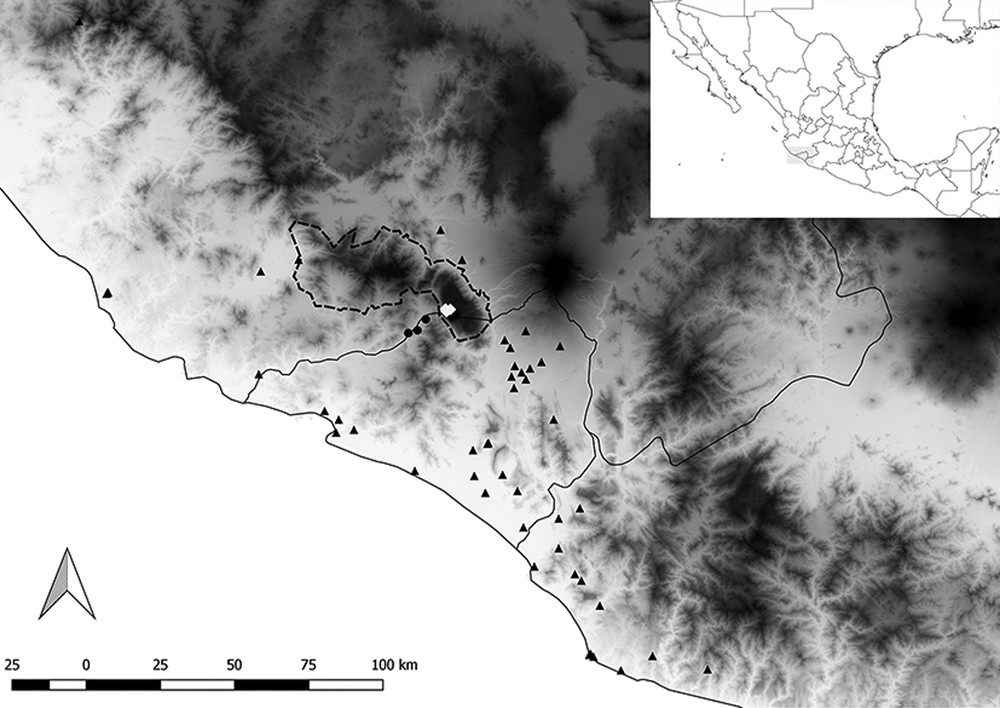
Map of Pacific-central Mexico showing southern Jalisco, Colima, and western Michoacán. The dashed line indicates the boundaries of the BRSM. Georeferenced points recorded by [8]: Centruroides tecomanus (triangles). Centruroides hirsutipalpus (circles), and Centruroides possanii sp. nov. (white diamonds). The topology of this part of Mexico is shown on a grayscale from see level (white) to ca. 4000 m of elevation (black).
Unlike vertebrates and other invertebrates, scorpions have not been surveyed in Colima systematically. Knowing that Colima is highly biodiverse and that scorpions have been poorly studied with appropriate methods like ultraviolet light detection at night [3], the discovery of undescribed species was anticipated.
Historically, records of scorpion species of public health importance have been documented in Colima [4–8]. For instance, Colima holds the highest historical rate of mortality per scorpion envenomation among the states of Mexico in a lapse of 56 years [8], and the rate of morbidity has been steadily increasing over the years following the nationwide trend [8,9]. Although no fatalities have been recorded, for the first seven months of 2019, a total of 4392 envenomation by scorpions have been reported in Colima, making Colima the eighth state with the highest number of scorpion envenomation cases in Mexico [10].
Presently, the scorpion fauna of Colima comprises nine described species included in five genera and three families, additionally, two species are yet to be described (Table 1). In this contribution, we describe a fifth species from the genus Centruroides from Colima, Mexico, Centruroides possanii sp. nov.
List of families, subfamilies, genus, and species recorded in the state of Colima.
| Buthidae | Centruroidinae | Centruroides elegans (Thorell, 1876) |
| Centruroides hirsutipalpus Ponce-Saavedra & Francke, 2009 | ||
| Centruroides infamatus (C.L. Koch, 1844) | ||
| Centruroides tecomanus Hoffmann, 1932 | ||
| Centruroides possanii sp. nov. | ||
| Vaejovidae | Syntropinae | Konetontli ilitchi González-Santillán & Prendini, 2015 |
| Mesomexovis aff. occidentalis | ||
| Thorellius cristimanus (Pocock, 1898) | ||
| Thorellius intrepidus (Thorell, 1876) | ||
| Vaejovinae | Vaejovis janssi Williams, 1980 | |
| Vaejovis monticola Sissom, 1989 | ||
| Vaejovis aff. santibagnezi |
2 Materials and methods
Scorpions were captured by ultraviolet detection at night [3] or rolling rocks and rotten logs during the day. Morphology and nomenclature followed González-Santillán & Prendini [11], except for that of the hemispermatophore, of which followed Monod et al. [12]. The dissection of the hemispermatophore followed Sissom et al. [3] and was cleared by hand. Denticle and granulation of the pedipalp chela fingers followed González-Santillán & Alvarez-Padilla [13], noticing that we include the apical subrow of denticles as part of the total count of denticles, unlike the current nomenclature based on Stahnke & Calos [14]. Lateral ocelli followed Loria & Prendini [15]. Measurements followed Sissom et al. [3] and are reported in the text like means, representing three males and eight females separated by “/”, including the holotype and the paratypes. Sexual dimorphism is indicated when present. All observations and photographs were taken using a Nikon SMZ 1000 dissecting stereomicroscope onto which a Nikon DS-U3 digital camera was mounted. Photographs were captured under LED and ultraviolet illumination. Type specimens will be deposited in the Colección Nacional de Arácnidos (CNAN) of the Instituto de Biología, and vouchers of DNA vouchers at Instituto de Biotecnología (IBt) of the Universidad Nacional Autónoma de México (UNAM). The distribution map was constructed with the program QGIS version 2.14.23-Essen [16]. The altitudinal layers used to construct the elevational model were obtained from Jarvis et al. [17].
3 Results
3.1 Systematics
Buthidae C.L. Koch, 1837.
Centruroidinae Kraus, 1955.
CentruroidesMarx, 1890.
Centruroides possanii sp. nov.
Zoobank: urn:lsid:zoobank.org:pub:AB1DA3A3-E723-4E50-A671-4C9AAD97FD1A.

Two adult, live females of Centruroides possanii sp. nov. from Los Saucos, Colima, showing the melanic variation within the population with paired blotches of infuscation in each sternite either covering the 80–90% or 30–40% of the surface.

Habitus of Centruroides possanii sp. nov. A, B, male; C, D, female; A, C, dorsal aspect; B, D, ventral aspect. Scale bar: 5 mm.
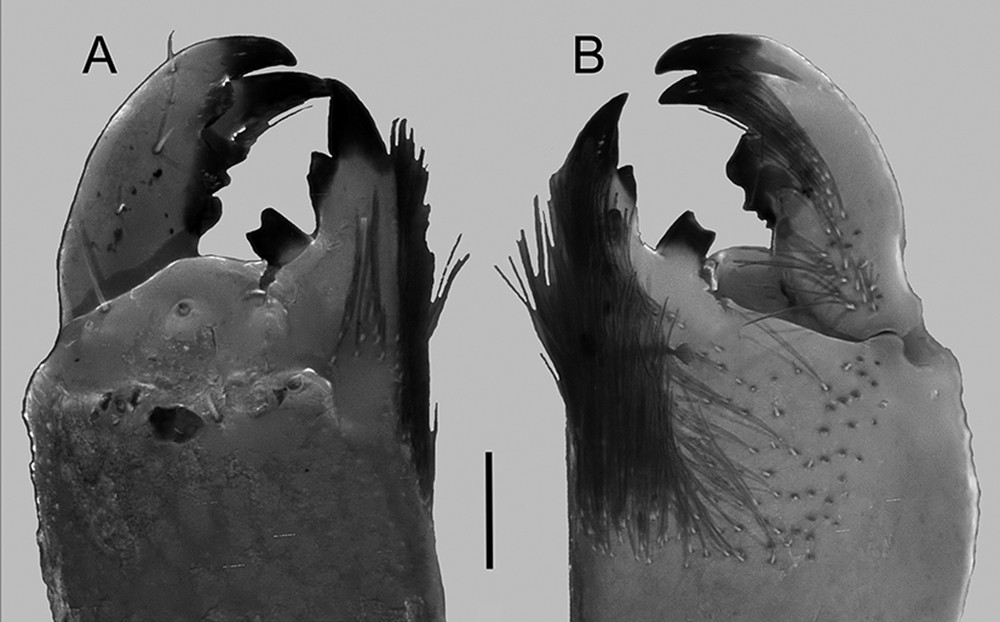
Sinistral chelicera of adult male of Centruroides possanii sp. nov. from Los Saucos, Colima. A, Dorsal aspect; B, ventral aspect. Scale bar: 0.2 mm.

Carapace of adult male of Centruroides possanii sp. nov. from Los Saucos, Colima. Scale bar: 1.0 mm.

Lateral ocelli of adult female of Centruroides possanii sp. nov. from Los Saucos. Colima showing Type 4A configuration. Abbreviations: ALMa, anterolateral major ocellus; MLMa, mediolateral major ocellus; ADMi, anterodorsal minor ocellus; PMLa, posterolateral major ocellus. Scale bar: 0.1 mm.
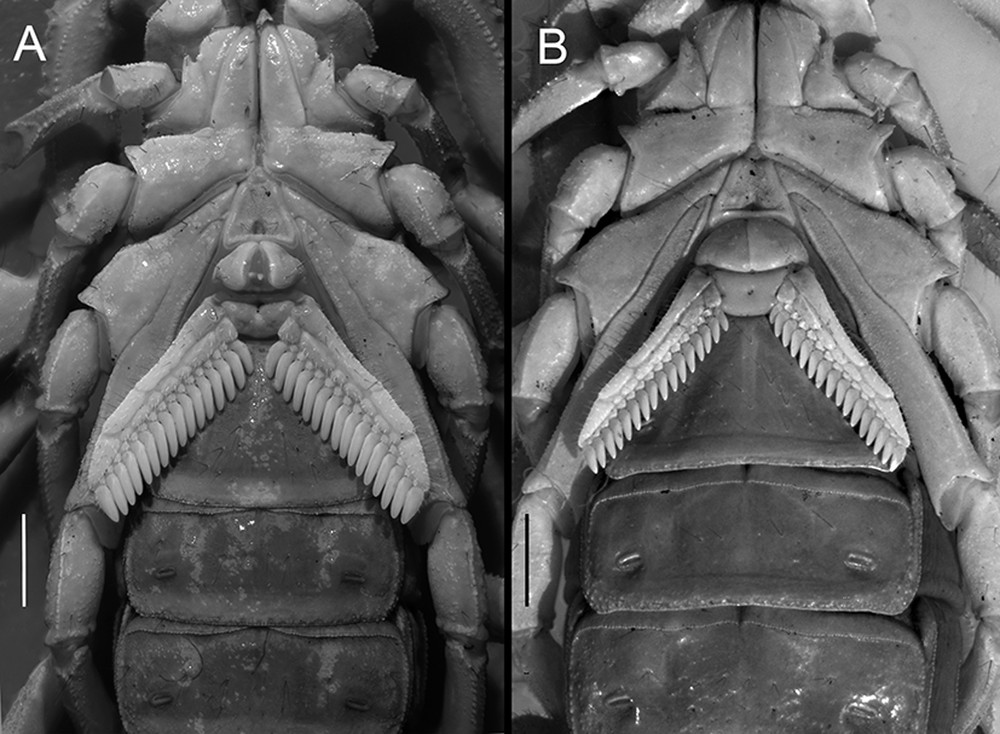
Coxosternal region. pectines and sternites III–V of Centruroides possanii sp. nov. from Los Saucos. Colima: A, male; B, female. Scale bars: 1.5 mm.

Sinistral femur of adult male of Centruroides possanii sp. nov. from Los Saucos. Colima: A, dorsal; B, prolateral; C, ventral. D, retrolateral. Scale bar: 1.0 mm.

Sinistral patella of adult male of Centruroides possanii sp. nov. from Los Saucos. Colima: A, dorsal; B, prolateral; C, ventral. D, retrolateral. Scale bar: 1.0 mm.

Sinistral pedipalp chela of adult male of Centruroides possanii sp. nov. from Los Saucos, Colima: A, dorsal; B, prolateral; C, ventral. D, retrolateral. Scale bar: 1.0 mm.
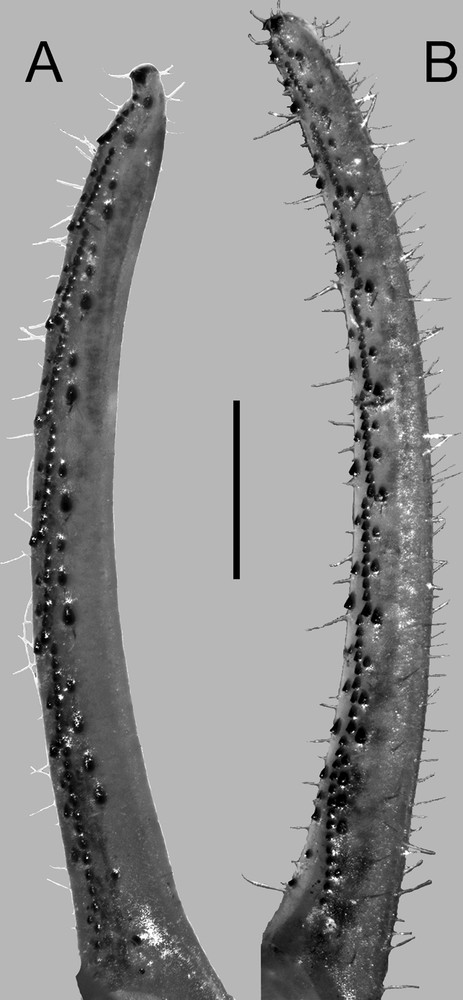
Sinistral pedipalp A. fixed and B. movable fingers of female Centruroides possanii sp. nov. from Los Saucos, Colima. Scale bar: 1.0 mm.

Sinistral basitarsus and telotarsus of adult males of Centruroides possanii sp. nov. from Los Saucos. Scale bar: 0.5 mm.

Sinistral hemispermatophore of Centruroides possanii sp. nov. from Los Saucos: A, pre-dissected and B, post-dissected, in lateral aspects. Abbreviations: bc, basal carina; bh, basal hook; dc, distal carina; f, flagellum; sd, sperm duct. Scale bars: 0.2 mm.
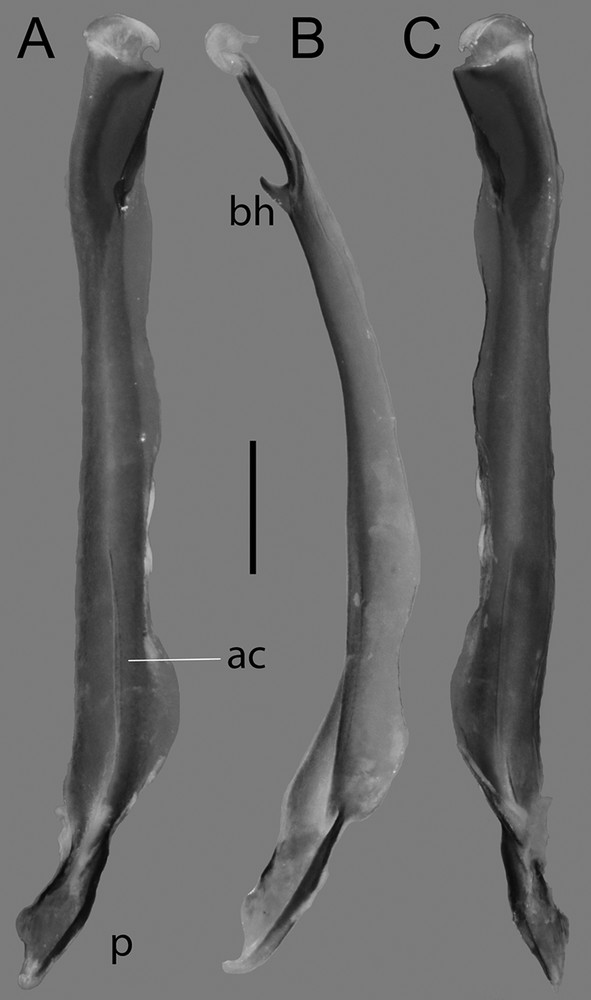
Sinistral hemispermatophore of Centruroides possanii sp. nov. from Los Saucos. A. Lateral. B. Anterior. C. Contra-lateral aspects. Abbreviations: ac, axial carina; bh, basal hook; p, pedicel.

Metasoma of adult males of Centruroides possanii sp. nov. from Los Saucos: A, dorsal; B, lateral. C; ventral aspects. Scale bar: 2.0 mm.
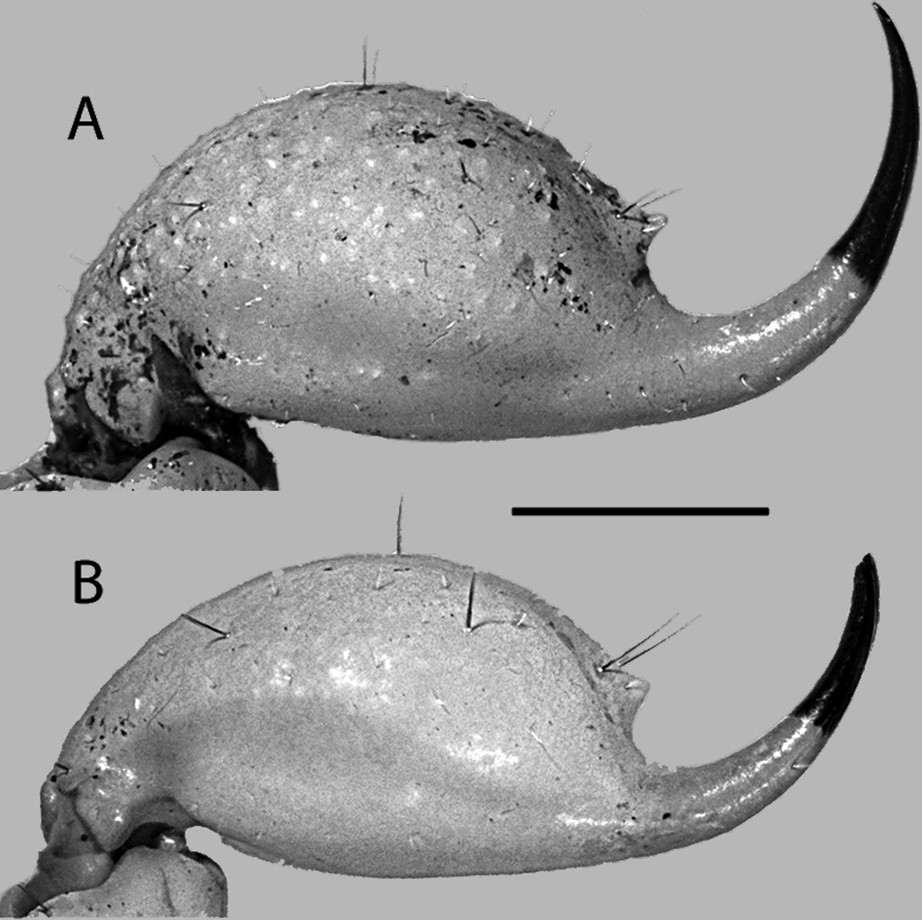
Telson lateral aspect of A. adult female and B. adult male of Centruroides possanii sp. nov. from Los Saucos. Scale bar 2.0 mm.
Measurements (mm) of adult Centruroides possanii sp. nov. The specimens will be deposited in the Colección Nacional de Arácnidos, Instituto de Biología, Universidad Nacional (CNAN).
| Catalog no. | T1360 | T1362 | T1362 | T1361 | T1361 | T1362 | T1362 | T1362 | T1362 | T1362 | T1362 | |
| Type | Hol. | Paratypes | ||||||||||
| Sex | ♂ | ♂ | ♂ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | |
| Carapace | length | 3.50 | 3.30 | 3.50 | 4.05 | 4.20 | 4.05 | 4.00 | 4.45 | 4.10 | 4.05 | 4.20 |
| anterior width | 2.50 | 2.50 | 2.50 | 2.90 | 2.90 | 3.10 | 3.00 | 3.05 | 3.00 | 3.00 | 3.00 | |
| posterior width | 3.75 | 3.65 | 3.80 | 4.35 | 4.60 | 4.75 | 4.75 | 4.95 | 4.60 | 4.70 | 4.70 | |
| Femur | length | 3.50 | 3.50 | 3.65 | 4.00 | 3.95 | 3.90 | 4.00 | 4.15 | 3.95 | 4.00 | 4.05 |
| width | 1.25 | 1.30 | 1.30 | 1.40 | 1.50 | 1.50 | 1.50 | 1.60 | 1.50 | 1.50 | 1.55 | |
| height | 1.00 | 1.05 | 1.00 | 1.05 | 1.10 | 1.10 | 1.15 | 1.15 | 1.05 | 1.15 | 1.15 | |
| Patella | length | 3.85 | 3.80 | 3.85 | 4.15 | 4.25 | 4.40 | 4.40 | 4.55 | 4.40 | 4.50 | 4.55 |
| width | 1.10 | 1.15 | 1.15 | 1.35 | 1.20 | 1.15 | 1.20 | 1.20 | 1.20 | 1.25 | 1.20 | |
| height | 1.70 | 1.65 | 1.65 | 1.50 | 1.95 | 1.90 | 1.95 | 2.10 | 1.90 | 2.00 | 1.95 | |
| Chela | length | 6.00 | 5.93 | 6.07 | 6.67 | 6.67 | 7.00 | 6.67 | 7.00 | 6.67 | 6.93 | 6.93 |
| Manus | width | 1.85 | 1.70 | 1.70 | 1.80 | 1.90 | 1.90 | 1.85 | 2.05 | 1.90 | 1.85 | 1.90 |
| Manus | height | 1.90 | 1.75 | 1.90 | 1.85 | 2.00 | 1.90 | 1.75 | 2.10 | 1.90 | 2.00 | 1.90 |
| Manus | length | 2.65 | 2.58 | 2.57 | 2.67 | 2.67 | 2.90 | 2.42 | 3.00 | 2.57 | 2.78 | 2.73 |
| Fixed finger | length | 3.35 | 3.35 | 3.50 | 4.00 | 4.00 | 4.10 | 4.25 | 4.00 | 4.10 | 4.15 | 4.20 |
| Mov. finger | length | 3.85 | 3.85 | 4.00 | 4.70 | 4.60 | 4.60 | 4.80 | 4.65 | 4.60 | 4.75 | 4.75 |
| Coxa II | length | 2.05 | 2.05 | 2.00 | 2.30 | 2.35 | 2.55 | 2.55 | 2.55 | 2.55 | 2.40 | 2.50 |
| Coxa IV | length | 4.50 | 4.35 | 4.10 | 5.25 | 5.40 | 5.35 | 5.10 | 5.40 | 5.35 | 5.00 | 5.00 |
| Sternum | length | 1.30 | 1.30 | 1.10 | 1.30 | 1.50 | 1.50 | 1.35 | 1.35 | 1.50 | 1.30 | 1.30 |
| anterior width | 1.00 | 0.95 | 0.95 | 0.95 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | |
| posterior width | 1.30 | 1.30 | 1.25 | 1.35 | 1.50 | 1.55 | 1.55 | 1.45 | 1.55 | 1.50 | 1.55 | |
| Mesosoma | length | 11.90 | 11.90 | 10.40 | 14.55 | 13.80 | 16.75 | 15.15 | 15.05 | 13.90 | 15.85 | 15.00 |
| Metasoma | length | 19.25 | 19.25 | 19.00 | 19.30 | 18.60 | 19.45 | 19.30 | 19.70 | 19.05 | 19.05 | 19.00 |
| Segment I | length | 3.00 | 3.00 | 3.00 | 2.95 | 2.95 | 3.00 | 3.00 | 3.00 | 3.00 | 3.05 | 3.00 |
| width | 2.10 | 2.05 | 2.05 | 2.40 | 2.55 | 2.55 | 2.55 | 2.60 | 2.55 | 2.50 | 2.50 | |
| height | 1.90 | 1.85 | 1.90 | 2.25 | 2.35 | 2.30 | 2.25 | 2.35 | 2.30 | 2.25 | 2.25 | |
| Segment II | length | 3.55 | 3.55 | 3.55 | 3.60 | 3.40 | 3.55 | 3.50 | 3.50 | 3.55 | 3.50 | 3.50 |
| width | 2.00 | 2.00 | 1.95 | 2.35 | 2.45 | 2.35 | 2.45 | 2.50 | 2.35 | 2.45 | 2.45 | |
| height | 1.85 | 1.80 | 1.85 | 2.05 | 2.20 | 2.25 | 2.15 | 2.50 | 2.25 | 2.20 | 2.25 | |
| Segment III | length | 3.85 | 3.90 | 3.95 | 3.85 | 3.75 | 3.90 | 3.80 | 3.95 | 3.80 | 3.75 | 3.75 |
| width | 2.00 | 2.00 | 1.95 | 2.35 | 2.45 | 2.40 | 2.45 | 2.50 | 2.40 | 2.45 | 2.45 | |
| height | 1.85 | 1.80 | 1.85 | 2.05 | 2.25 | 2.25 | 2.15 | 2.45 | 2.25 | 2.20 | 2.25 | |
| Segment IV | length | 4.10 | 4.20 | 4.00 | 4.20 | 4.00 | 4.25 | 4.25 | 4.25 | 4.15 | 4.15 | 4.20 |
| width | 2.00 | 2.00 | 1.95 | 2.35 | 2.45 | 2.40 | 2.35 | 2.50 | 2.40 | 2.45 | 2.45 | |
| height | 1.75 | 1.80 | 1.85 | 2.05 | 2.25 | 2.25 | 2.15 | 2.25 | 2.25 | 2.20 | 2.25 | |
| Segment V | length | 4.75 | 4.60 | 4.50 | 4.70 | 4.50 | 4.75 | 4.75 | 5.00 | 4.55 | 4.60 | 4.55 |
| width | 2.10 | 2.05 | 1.80 | 2.05 | 2.20 | 2.05 | 2.05 | 2.30 | 2.05 | 2.00 | 2.00 | |
| height | 2.05 | 2.05 | 2.00 | 2.05 | 2.05 | 2.25 | 2.10 | 2.25 | 2.25 | 2.20 | 2.20 | |
| Telson | length | 3.75 | 3.55 | 3.50 | 3.55 | 3.60 | 3.85 | 3.85 | 3.75 | 3.95 | 3.95 | 3.95 |
| Vesicle | length | 2.80 | 2.65 | 2.65 | 2.60 | 2.65 | 2.80 | 2.80 | 2.80 | 2.75 | 2.75 | 2.75 |
| width | 2.00 | 1.75 | 1.75 | 1.80 | 2.00 | 2.10 | 2.05 | 2.00 | 2.10 | 2.00 | 2.05 | |
| height | 1.75 | 1.70 | 1.70 | 1.75 | 1.85 | 1.95 | 1.90 | 1.85 | 1.95 | 1.85 | 1.90 | |
| Aculeus | length | 0.95 | 0.90 | 0.85 | 0.95 | 0.95 | 1.05 | 1.05 | 0.95 | 1.20 | 1.20 | 1.20 |
| Total | length | 38.40 | 38.00 | 36.40 | 41.45 | 40.20 | 44.10 | 42.30 | 42.95 | 41.00 | 42.90 | 42.15 |
Type material: Mexico: Colima: Municipio de Minatitlán: Holotype ♂ (CNAN-T01360) and 2 paratype ♀ (CNAN-T1361) Rancho los Suacos, 11 km NE of Minatitlán, 19°26'12''N 103°57'49''W, 2097 m, 30.ix.2018, oak forest, L.L. Valdez, M. Bermudez, P.E. García J. S. Ramírez, ultraviolet light detection by night; 2 ♂♂ and 6 ♀♀ paratypes (CNAN-T01362), 20.vi.2017, E. Cárcamo, J. Cid, J.C. Fragoso, E. González-Santillán, R. Ibarra, J.M. Jiménez, T. Olamendi, T. Romero, L.L. Valdez.
Etymology: The species name, possanii, honors Dr. Lourival Domingos Possani Postay, for his contributions to the biochemistry, public health importance, evolution, and several aspects of the scorpion biology.
Diagnosis:Centruroides possanii sp. nov. may be distinguished from other species of Centruroides of the North-American clade [18], including the elegans species group [19] by the presence of infuscate carapace with symmetric pattern of markings (Figs. 2 and 3), instead of having four parallel, longitudinal lines of infuscation on the carapace. Centruroides possanii sp. nov. superficially resembles species from the nigrovariatus and infamatus groups [19], but differs from the nigrovariatus by the distinctive metasomal segments I–V width and height that increases distally, but is relatively constant in Centruroides possanii sp. nov. (Table 2). From the infamatus group Centruroides possanii sp. nov. differs by a smaller body length at a range from 38 to 42 mm, whereas members of the infamatus group are 40 to 70 mm total length body of adult male and female [19]. Centruroides possanii sp. nov. is morphologically distinct from the four species that inhabit the state of Colima, Centruroides elegans (Thorell, 1876), Centruroides hirsutipalpus Ponce-Saavedra & Francke, 2009, Centruroides infamatus (C.L. Koch, 1844), and Centruroides tecomanus Hoffmann, 1932, by the following characters: it differs from the first three taxa by the presence of a median depression on the basal piece of the pectines in the female (Figs. 3 and 7B), which is absent in these species. Centruroides possanii sp. nov. differs from C. tecomanus by having the subaculear tubercle smaller and the apex oriented toward the medial portion of the aculeus, instead of oriented toward the apex of the aculeus. Centruroides possanii sp. nov. can be distinguished from the adjacent distributed species, C. hirsutipalpus by having only three setae on the prolateral surface of the trochanter, instead of having more than 20.
Description: The following description is based on the type series and includes adult males and females.
Color and infuscation. Base color yellowish straw to orange-ochraceous. Carapace, dorsal prolateral and retrolateral surfaces of pedipalp trochanter, femur, patella and manus, and tergites, prolateral surface of legs, dorsal and lateral surfaces of metasomal segments first to fourth moderately infuscate, all surfaces of the fifth segment and sternites sixth and seventh faintly infuscate, other surfaces of the body immaculate (Fig. 3). Chelicerae manus base color same as the rest of the body with evidently reticulated infuscation, fingers immaculate. Carapace with anterior submedian and posterior lateral carinae evidently infuscate, ocular tubercle margins evidently infuscate, each median ocellus encircled by an infuscate areola weak anteriorly and broader posteriorly, lateral ocelli evidently infuscate. Tergites with paired longitudinal stripes, comprising two moderate blotches, one on the pretergites and other on the postergites connected by a reticulated pattern of fainter infuscation, with two consistent forms: paired blotches of infuscation in each sternite either covering the 80–90% or 30–40% of the surface (Figs. 2 and 3), other surfaces immaculate. Pectinal teeth whitish. Distal portion of the aculeus reddish distally.
Chelicerae. Manus dorsal surface with sparsely small granules, lustrous, with a transversal subdistal smooth carina with three distal macrosetae, decreasing in size from median to lateral surface. Prolateral and ventral surfaces of the fixed fingers densely cover with macrosetae. Fixed finger prolateral margin basal and median teeth joint into a bicuspid, subdistal tooth similar size than the median, distal tooth blade-like; retrolateral surface with one median tooth (Fig. 4). Movable finger, prolateral margin with two small basal teeth, most of the time, worn out in adults and only sharp and visible in juveniles; median and subdistal tooth similar in size, larger than basal; retrolateral margin with medial and subdistal teeth similar in size. Prolateral and retrolateral distal teeth opposable, forming a fork.
Carapace. Length slightly greater than to posterior width (0.90/0.88). Surface shagreened, with scattered conical granules, all sulci cover with minute granulation or smooth (Figs. 3 and 5). Distal margin with two pairs of macrosetae; emarginated, gently curved, extreme lateral margins curved, lateral ocelli in a lateral frontal position. Four pairs of lateral ocelli congruent with the Type 4A, with PLMa, MLMa ALMa equal size and ADMi vestigial, three lateral ocelli delimited from ADMi by a granular carina (Fig. 6). Superciliary carinae of lateral ocelli granular. Median ocular tubercle raised, situated in the anterior half of the carapace (Figs. 3 and 5); superciliary carinae granular, higher than median ocelli. Anteromedian and posteromedian sulci deep; anterolateral and posterior transverse shallow.
Coxosternal region. Sternum subtriangular, truncated with a sagittiform notch posteriorly (Fig. 7), length shorter than posterior width (0.96/0.92), with two pairs of microsetae. Median sulcus deep with an anterior depression. Coxa IV more than twice the length of coxa II (2.11/2.12). Coxae I–IV ventral surfaces smooth, lustrous with all margins granular. Coxa II, prolateral subproximal margin with a low granular protuberance.
Pedipalps. Femur length (2.76/2.65) times greater than width; all intercarinal surfaces matte (Fig. 8A–D); dorsal prolateral, dorsal retrolateral, ventral prolateral and retrolateral dorsosubmedian carinae complete, irregularly granular; prolateral ventrosubmedian carina with scattered coarse scatter granules and three macrosetae (Fig. 8D); retrolateral ventral and ventral median carinae vestigial, reduced to a short row of granules proximally. Patella length (2.30/2.31) times greater than width and width (1.30/1.27) times greater than femur width; all intercarinal surfaces matte (Fig. 9A–D); dorsal prolateral, dorsal median, dorsal retrolateral, retrolateral median, ventral retrosubmedian, and ventral prolateral carinae complete, finely granular; prolateral ventral and prolateral dorsal process moderate with a proximal spiniform enlarged tubercle on each (Fig. 9A and C), prolateral median carina vestigial with few scatter spiniform granules and a proximal macroseta, prolateral ventral carina vestigial with three to four coarse scattered granules and a proximal macrosetae (Fig. 9D). Chela length (3.43/3.60) times than width, (1.70) times greater than femur length and (1.57/1.55) times than patella length, width (1.36/1.26) times greater than femur width and (1.05/0.99) times than patella width; all dorsal, prolateral and retrolateral carinae complete finely granular (Fig. 10A, B and D), all ventral surface acarinated except for ventral retrolateral carinae finely granular (Fig. 10C); three costate, dorsal carinae extending one to two thirds on the fixed finger (Fig. 10A and D).
Pedipalp fixed and movable fingers. Notches, lobes, and gap when fingers closed moderate (♂) or weak (♀), profusely setose (Fig. 10B and D), dentate margin with rows of serrate, median oblique and flanking accessory prolateral and retrolateral denticles (Fig. 11A and B). Fixed finger median row comprising eight imbricate, oblique denticle subrows, with five denticles in position I, eight in position II–VII, and 17 or 18 in position VIII; flanked by eight prolateral and eight pairs of retrolateral enlarged denticles, and by a two-denticle retrolateral accessory subrows separated equidistally along each median subrow, and by one prolateral accessory denticle placed midway along each median subrow; distal subrow without accessory denticles (Fig. 11A). Distal retrolateral denticle with two proximal, linear, tapered distally macrosetae longer than distal denticle. Movable finger median row comprising eight (♂) or eight to nine median denticle subrows (♀), with three denticles in position I, eight to nine in positions II–VII (♂) or II–VIII (♀) and 13–15 in position IX (♀); flanked by eight retrolateral and eight prolateral subrows of denticles (♂) or nine retrolateral and nine prolateral subrows of denticles (♀) and by two retrolateral and two prolateral accessory denticles in positions V–VII, separated equidistally along each median subrows (Fig. 10B); position I without retrolateral accessory or prolateral accessory denticles, position II–IV with one retrolateral accessory and one prolateral accessory denticle, position VIII with five to six retrolateral and five to six prolateral accessory denticles (♂), position IX with three prolateral and tree retrolateral accessory denticles (♀). Distal retrolateral denticle with two pairs of modified macrosetae, one pair proximal linear tapered distally and the other pair lateral shaped like half of a spatula. Trichobothrial pattern Type A, configuration α (Figs. 8–10).
Legs. Basitarsi I–IV: dorsal, prolateral dorsal, prolateral ventral, retrolateral ventral and retrolateral dorsal carinae with dense rows of microsetae; retromedian and dorsal surfaces with accessory rows and clusters of microsetae. Telotarsi I–IV, with irregularly clustered prolateral ventral and retrolateral ventral rows of dense microsetae (Fig. 12). Ungues short and curved.
Genital operculum. Width greater than length, with three pairs of lateral macrosetae; sclerites separate longitudinally. Genital papillae present reduced not protruding posteriorly (♂) (Fig. 7) or absent (♀).
Hemispermatophore. Flagelliform, stalk with flagellum coiled twice on the distal margin of capsular area, relatively short (Fig. 13A). Basal hook strongly sclerotized and elongated, blunt (13B). Sperm duct devoid of folding, elongated, basal and distal carinae strongly sclerotized (Fig. 13B). Stem fusiform with a strong axial carina (Fig. 14). Truncal flexure absent, pedal flexure well-developed (Fig. 14). Pedicel with margins strongly sclerotized.
Pectines. Basal piece, narrowed (♂) without median depression or broadened with deep median depression (♀). Marginal lamella comprising three pieces, median lamellae with six to seven pieces each. Fulcra with 15–16 pieces. Pectinal teeth count 16–18 (♂) 14–16 (♀), relatively short, distal margin of marginal lamella as long (♂) or slightly shorter (♀) than coxa IV margin (Fig. 7).
Tergites. I–VII, intercarinal surfaces shagreened, densely covered with minute and coarse granules mostly on pigmented areas (Fig. 3A and C); dorsal median carina complete, granular, other carinae absent; VII; dorsal submedian and dorsal sublateral carinae granular subparallel.
Sternites. Sternite III–VII, surface smooth, lustrous except lateral surfaces, sparsely granular (♂) or increasing granulation distally (♀); III, with an anteromedian triangular glandular hyaline area, V similar anteromedian hyaline glandular area, but with six to eight associated setae (♂) or anteromedian triangular glandular hyaline area vestigial (♀, Fig. 3B and D); VI, with granular ventrolateral carinae on posterior half of the sternite; VII, ventrolateral carina granular, incomplete on posterior two thirds of the sternite and ventral sublateral carina granular, incomplete on median third of the sternite.
Metasoma. Length (1.7/1.3) times greater than mesosomal length (Table 2); segments I–V length (1.5/1.2, 1.8/1.5, 2.0/1.6, 2.1/1.7, 2.3/2.2) greater than width; segment V width (1.1/1.0) times greater than telson width. Segments I–V all intercarinal surfaces sparsely granular (♂) or densely granular (♀), increasing distally; dorsal lateral, lateral median, ventral lateral, and ventral submedian carinae on segment I–IV complete, finely serrate, costate (Fig. 15); lateral inframedian carina on segment I complete, finely serrate, costate; on segment II vestigial with a row of three or four posterior granules (♂) or less granules (♀); on segments III and IV absent. Segment V, dorsal lateral carina obsolete with few scattered granules (♂) or complete, finely serrate, costate (♀), lateral median and lateral inframedian carinae absent; ventral lateral and ventral median carinae, complete, finely serrate, costate (♀) or flat, weakly serrate (♂). Macrosetae counts on carinae of segments I–V, respectively: dorsal lateral, 0:0:0:0:0; lateral median, 0:0:0:0:0; lateral inframedian, 0:0:0:0:0; ventral lateral, 2:2:2:2:2; ventral sublateral, 0:0:0:0:2; ventral submedian, 2:2:2:2:2.
Telson. Vesicle globose, total length (2.0/1.9) times greater than vesicle width (Table 2); all surface smooth (♂) or granular on ventral surface (♀); all carinae obsolete with two macrosetae each (Fig. 16). Ventrolateral surface with a pair of hyaline oblong glandular areas. Aculeus, moderately elongated and curved laterobasal microserration absent, subaculear tubercle moderate, venom delivery openings slit like, paired.
Distribution. Known only from three localities in Colima.
Ecology.Centruroides possanii sp. nov. can be found under rocks, the bark of fallen trees, or debris on the ground. The habitat and habitus of this species indicate that its ecomorphotype may be classified as errant [20] or corticolous [21]. During the night was collected with ultraviolet detection and was observed hunting using the sit-and-wait (ambush) strategy of forage. The altitude of the collection localities ranged from 2013 to 2200. It was collected in sympatry with Vaejovis aff. santibagnezi.
Remarks. Centruroides possanii sp. nov. may be assigned to the infamatus subgroup of Ponce-Saavedra & Francke [19] due to the similar infuscation markings on the carapace characterized by asymmetric variable pattern covering 60 to 80% of the surface and the infuscation of the tergites with two longitudinal stripes of variable breadth separated by a medial narrow strip of base color, lacking infuscation. However, as we explained below, we refrain to assign Centruroides possanii sp. nov. to any of the subgroups of Ponce-Saavedra & Francke.
Additional material examined. MEXICO: Colima: Municipio de Minatitlán: El Terrero, 19°26'35.94''N 103°57'05.67''W, 2200 m, 10.x.2015, J.S. Ramírez, L.L. Valdez, 1 ♂, 1 ♀ juvenil (IBt, UNAM); El Terrero, Mirador el Filete, 19°26’40“N 103°58'1''W, 2137 m, 11.x.2015, J.S. Ramírez, L.L. Valdez, 1 ♀, 1 ♂ juvenil (IBt, UNAM).
On the distribution of Centruroides possanii sp. nov. and its implications for conservation.
The tree localities recorded here, El Terrero, Mirador el Filete, and Los Saucos, are located in the northwestern extreme of Colima in the municipality of Minatitlán (Fig. 1), which are part of the Biosphere Reserve Sierra de Manantlán (BRSM). Physiographically, BRSM belongs to the Sierra Madre del Sur, but it is in direct contact with the Sierra Madre Occidental and the Trans-Mexican Volcanic Belt [22]. The dominant vegetation in these localities is the oak/pine forest and is circa 50 km from the Pacific Ocean, thus the humidity is high and constant throughout the year. The average annual temperature is 14 °C and the annual precipitation is approximately 1700 mm. These localities are on the western part of Cerro Grande (Fig. 1), an isolated, karstic dome extending over an area of 25 by 15 km. This area comprises terrains that originated during the Cretaceous and the heterogeneous landscape is attributed to the subduction of the Pacific plate beneath the Mesoamerican plate [22]. The Cerro Grande mountain exhibits steep slopes along its complete periphery, isolating itself from other mountains and the main mass of sierras that include the BRSM [22]. In contrast, the lower elevation engulfing Cerro Grande is characterized by a tropical climate and vegetation with an annual temperature of 26 °C and annual precipitation of circa 1000 mm [23]. These valleys are composed of young volcanic rocks from the Tertiary and the Quaternary [24]. The combination of all these physiographic and climatic features emphasize the uniqueness of this mountain. For these reasons, we think that it is reasonable to hypothesize that Centruroides possanii sp. nov. is a microendemic to Cerro Grande and that its conservation shall be a priority. Future expeditions in search of this species outside and nearby Cerro Grande shall test the idea of such a restricted distribution.
4 Discussion
Ponce-Saavedra et al. [6] reported five species of Centruroides: C. elegans, C. hirsutipalpus, C. infamatus, C. tecomanus, and Centruroides limpidus (Karsch, 1879) from Colima; we confirm the presence of all species reported, except for C. limpidus. It is unlikely that natural populations of C. limpidus inhabit Colima due to the approximately 200 km of distance between Colima and the central areas of Michoacán reported recently, which represents the closest record of C. limpidus [8].
Ponce-Saavedra et al. [19] recently proposed a scheme of species groups and subgroups to organize the Mexican Centruroides based on the color of the body, different patterns of infuscation on the carapace and mesosomal tergites (Fig. 3), and the number of imbricated subrows of denticles of the cutting edge of the fingers of the pedipalp manus of these scorpions (Fig. 11) [8,25–29]. These groups appear to be non-monophyletic when compared to the North-American clade recovered by Esposito & Prendini [18]. According to Esposito & Prendini, the North-American Centruroides comprises three clades that vaguely resemble the scheme proposed by Ponce-Saavedra et al. [19]. The diagnostic morphological characters proposed by Ponce-Saavedra et al. have yet to be evaluated by phylogenetic analyses. Some of these characters are likely to be homoplastic. For instance, the different patterns of infuscate markings on the carapace used to distinguish the elegans from the infamatus and the nigrovariatus subgroups [19], mapped onto the phylogeny inferred by Esposito and Prendini [18] make these subgroups polyphyletic. Further analyses are in progress to help to resolve the incongruence between the morphological and molecular evidence we are faced with currently within Centruroides.
Locals from the vicinities of the type locality, at Los Saucos, informed us that the sting of Centruroides possanii sp. nov. produces the typical symptoms of moderated intoxication, such as pruritus in the nose, mouth, and throat, and foreign sensation in the throat [8,9]. However, the venom of this species has never been tested experimentally for its potency. Interestingly, four out of six species present in Colima are within the topmost venomous species of Centruroides in Mexico. These species include C. elegans, C. hirsutipalpus, C. infamatus and C. tecomanus with an LD50 between 9.6 and 11.7 μg of venom/20 g of mouse, compared to other 12 species with an LD50 over 12 μg of venom/20 g of mouse [7]. This may account for the record of mortality during 56 consecutive years prior to the introduction of commercial antivenom in Mexico [8].
Acknowledgments
This work was supported by a postdoctoral scholarship from CONACYT, Mexico (grant IN237864) to EG-S. Permits for collecting wild specimens were provided by SEMARNAT: FAUT-0305 to EG-S. We thank Erika García for English editing and two anonymous reviewers for helping improving this manuscript.