

1 Introduction
The temperature of the upper 300 m of the North Atlantic increased by over 0.4 °C in the two last decades [1]. Correlatively, in the eastern North Atlantic, evidences of northward shifts were brought for fish species [2–4]. At the more local scale of the Bay of Biscay (France), a warming of the sea-surface temperatures was observed [5]. The Bay of Biscay is the subtropical boreal transition subprovince between the Atlantic boreal province and the northern subtropical subprovince, so life may respond rapidly to small shifts in climate. Some first evidences of changes at the aggregate level of the demersal fish community of the continental shelf of the Bay of Biscay have been brought recently [6]. Abundance trends for flatfish species were also reported [7]. The present warming in temperate areas could increase the breeding success for subtropical species. However, the existence of such causal relationships between warming and increase of abundance remains hypothetical, because the biological changes observed concerned species living at the bottom, while bottom-temperature variations have never been analysed on the continental shelf. Hence we do not know if warming spread through the water column. Moreover, fishing effects on species abundance may be a confounding factor when analysing biological changes induced by climate.
Abundance increase of a small bathy-demersal subtropical species, the boarfish Capros aper, was observed from Plymouth westwards (UK) during 9 years, between 1870 and 1879, while this species was one of the rarest fish species in these waters before 1870 and after 1879 [8]. Climate events were advocated to explain this phenomenon. More recently, in the western coast of Portugal, an increase of the abundance of the boarfish between 1980 and 1990 has been reported when the warming began to be observable there [9]. Climate change may then have some impact of this fish population dynamics. Our objectives are to test whether the abundance of boarfish has increased in the Bay of Biscay during the three last decades and whether a warming may be observed also at the bottom (a warming of the sea-surface temperature is already known) during the breeding period to explain the abundance increase.
2 Materials and methods
2.1 Elements of the boarfish biology
Very few information on the biology of boarfish was available. In the Irish Sea, this species were not studied (Minchin, comm. pers.). In the Portuguese waters, some recent works on the boarfish biology were carried out, but mainly on diet [10]. Some aspects of the biology of this species were described in the fish fauna [11]. Boarfish is found in the eastern Atlantic, from western Norway, Skagerrak, Shetlands and western Scotland to Senegal and also in the western Mediterranean Sea. Characteristic morphological traits are the presence of really strong dorsal and anal spines and its deep red colour. Reported sizes are usually 5 to 13 cm up to 16 cm. Longevity and ages are unknown. This is a gregarious species that forms schools, found over rock or coral and sometimes over sandy ground mainly at 100–300-m depth. In the southwestern Ireland waters, where it reproduces in summer (June–August), boarfish feeds on crustaceans. Moreover, boarfish is rarely reported as a prey in the stomach content analyses of the main predators. Actually, boarfish is not found in the stomach contents of hake, Merluccius merluccius, analysed in the Bay of Biscay [12,13]. Capros aper is a prey of Conger conger and of Phycis phycis off the Azores [14]. Diet of these two last predators remains unknown in the Bay of Biscay. More analyses would be needed to identify the existence of potential competitors in the Bay of Biscay. This species was exploited by Portuguese fisheries in the waters of Portugal (Hill, comm. pers.) during the 1960s, but not in the Bay of Biscay.
2.2 Abundance
The data used were collected during 12 autumn groundfish surveys carried out by IFREMER from 15-m up to 450-m depth on the eastern continental shelf of the Bay of Biscay (
2.3 Temperature
Temperature data from the World Data Center A (WDC-A) and temperature data collected by the French Research Institute for the Exploitation of the Sea (IFREMER) and the French Hydrographic and Oceanographic Service of the Navy (SHOM) were used for this study. The temperature measurements were made using a conductivity–temperature–depth (CTD), a bathythermograph (XBT, MBT) and bottle stations, over the period 1967 to 2002 in the zone of the Bay of Biscay between 43°–48°N and east of 5°. The average temperature of two water layers, at 100- and 200-m depth, was calculated for a three-month period (June, July, August), corresponding to the breeding season of Capros aper, using data combined from all of the stations in the zone of study. 867 stations were available for the whole period, on average 20 per year.
2.4 Temporal trends and correlations
To characterize the temporal trend of the boarfish abundance and of the temperature, three models were fitted, a linear, quadratic, and an exponential one. Non-parametric tests of correlation between abundance and temperature were performed on the log transformed datasets: Spearman rank order (R), Gamma (G) and Kendall tau correlation.
3 Results and discussion
The abundance of Capros aper actually significantly increased from about 7 to 1500 individuals per haul (Fig. 1). The best-fitted model is the exponential one with a r-squared value of 0.89 against 0.6 and 0.79, respectively, for the linear and quadratic models. The observed increase has really occurred in the late nineties. The relative abundance of boarfish increased from 0.27% in 1973, 15 species were then more abundant than boarfish, to 16.43% in 2000 with only two species more abundant than boarfish (Table 1). Capros aper is now one of the dominant species in the fish community. The main depth range of its distribution (100–300 m) encompasses the area sampled each year. We have then computed the mean abundance per haul from 200-m up to 450-m depth for each year except for the years 1973 and 1992. Despite highest abundances at the end of the time series in 1998, 1999 and 2001 (Fig. 2), the increasing trend is not significant on such depth according to the Spearman test (
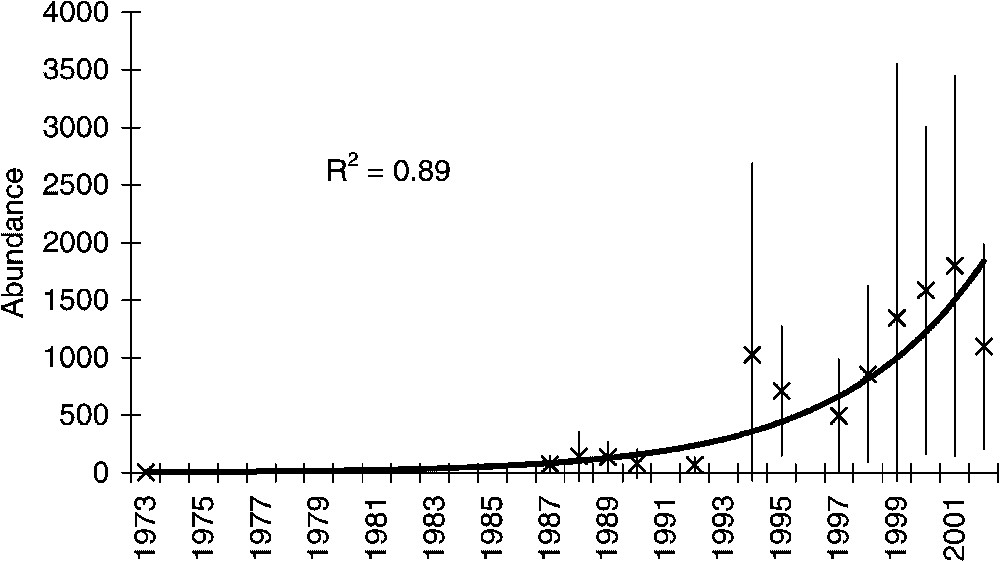
Mean abundance of boarfish (Capros aper) per haul and confidence interval (5%) in the Bay of Biscay from 1973 to 2002 between 15-m depth up to 200-m depth and exponential trend.
Temporal variations of the abundance rank and relative abundance (%) of the boarfish (Capros aper) in the fish community of the Bay of Biscay between 1973 and 2002
| Year | Rank | % |
| 1973 | 16 | 0.27 |
| 1987 | 6 | 1.17 |
| 1988 | 6 | 2.11 |
| 1989 | 4 | 2.95 |
| 1990 | 9 | 1.19 |
| 1992 | 8 | 1.16 |
| 1994 | 3 | 4.88 |
| 1995 | 4 | 6.71 |
| 1997 | 5 | 4.25 |
| 1998 | 4 | 8.41 |
| 1999 | 3 | 7.34 |
| 2000 | 3 | 16.43 |
| 2001 | 3 | 15.96 |
| 2002 | 3 | 5.9 |
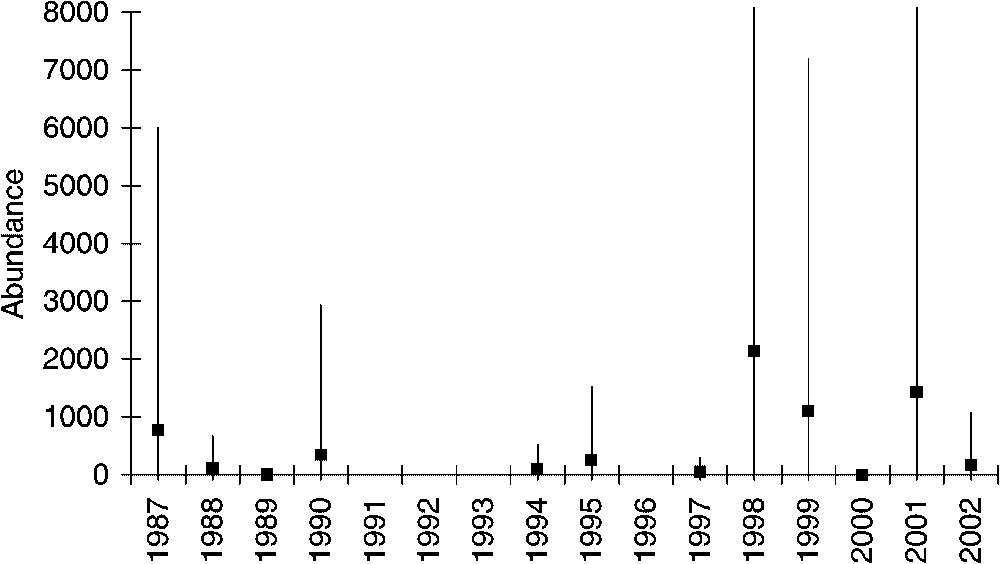
Mean abundance of boarfish (Capros aper) per haul and confidence interval (5%) in the Bay of Biscay from 1987 to 2002 between 200-m depth up to 450-m depth.
An increase in the temperatures during the breeding season was observed (Fig. 3). This warming is coherent with the surface signal and is present over the whole column of water. This tendency was observed to be independent of the quality of the data and of the field of study as signals are coherent (data not shown) with those observed in other zones of the Bay of Biscay and also in winter. For the 200-m depth layer, the best fitted model is the quadratic one with an r-squared value of 0.67 against 0.65 and 0.66, respectively, for the linear and exponential models. For the 100-m depth layer, the r-squared value is 0.61 for the quadratic, linear and exponential models. The temperature gradient over the whole period is around 0.5 °C. As for the observed abundances, the observed temperature increase has really occurred in the late nineties. The correlation between abundance and temperature at 100 and 200 m were statistically significant for all the three tests (Table 2).

Temporal variations in the average temperature during the breeding period (June to August) to a depth of 100 m (full squares) and 200 m (crosses) in the zone of the Bay of Biscay between 43°–48°N and east of 5°W from 1973 to 2002.
Non-parametric tests of correlation between the abundance of the boarfish (Capros aper) and the temperature at 200 m depth during the breeding season in the Bay of Biscay between 1973 and 2002 (log transformed data). The results of the tests for temperature at 100 m depth are exactly the same as those for 200 m, they are not shown in the table
| Spearman rank order correlation | |
| Spearman R | 0.629 |
| t(N−2) | 2.914 |
| p | 0.012 |
| Gamma correlation | |
| Gamma | 0.467 |
| Z | 2.425 |
| p | 0.015 |
| Kendall tau correlation | |
| Kendall | 0.467 |
| Z | 2.425 |
| p | 0.015 |
These results confirmed that in the Bay of Biscay warming occurred not only at the surface but also in the whole water column. Warming may not only favour northern immigration of subtropical species, but also may change local abundance. It has been shown for cod populations that recruitment is significantly related to temperature during the reproduction stage [15]. Experimental studies have shown that the reproductive performances of fish are closely linked to temperature [16]. One causal possible explanation is a direct physiological effect on reproduction. Temperature may actually modulate hormone action, affecting especially ovulation and spawning.
Longevity and ages of boarfish are unknown, but usually, small fish species are short-lived, fast growing and early reproducing species. Species with these biological characteristics are considered as having capacity to invade and colonize new areas (r-strategy). Colonisation may be fast if the mortality is low (no predation and no fishing). These observations are not so anecdotal in the global warming context, as it allows us to think that such changes may also concern in the near future northern species of larger size and of commercial interest for fishery and so concern the whole biodiversity and its functions in temperate ecosystems.
Acknowledgments
All data contributors are deeply acknowledged, in particular the IFREMER, the SHOM and the WDC-A. We would like to thank A. Bonnat, M. Fichaut, and C. Maillard for the qualification of all the data and the service of Coriolis, DEL/AO and SISMER for the data digitalization.
Vous devez vous connecter pour continuer.
S'authentifier