

1 Introduction
The scorpion fauna of Southeast Asia remains yet poorly studied. Pioneer work was conducted by Fage [1–3], who described interesting new elements of this fauna [1,4]. Subsequently, some isolated contributions revealed additional new species, such as the monographic work of Couzijn [5] on the genus Heterometrus Ehrenberg, 1828, and the revisions of the genera Chaerilus Simon, 1877 and Isometrus Ehrenberg, 1828 by Kovařik, [6,7]. Unfortunately, Kovařik's descriptions of the new species of Chaerilus and Isometrus are rather insufficient and poorly illustrated. The study of further specimens from the collections of the Muséum national d’histoire naturelle, Paris, allowed more precise diagnoses and proper illustrations of both Chaerilus petrzelkai Kovařik, 2000 and Isometrus (Reddyanus) petrzelkai Kovařik, 2003, both from Vietnam [8,9]. These studies also led to the description of two new species of Chaerilus, C. laoticus Lourenço & Zhu, 2008 from Laos and C. vietnamicus Lourenço & Zhu, 2008 from North Vietnam [8]. Particularly remarkable was the recent description of Troglokhammouanus steineri Lourenço, 2007 from caves in Laos, representing a new genus and species of the enigmatic family Pseudochactidae Gromov, 1998, previously known only from Tajikistan and Uzbekistan [10].
2 The genus Isometrus Ehrenberg, 1828
As previously discussed [9,11], the genus Isometrus was described by Ehrenberg [12] with Buthus (Isometrus) filum Ehrenberg, 1828 [= Isometrus maculatus (DeGeer, 1778)], as type species by monotypy. Vachon [13] proposed the division of the genus Isometrus into two subgenera: Isometrus Ehrenberg and Reddyanus Vachon. The diagnoses of the two subgenera were only proposed in a short key and based on the relative positions of certain trichobothria. Isometrus has trichobothrium db of the fixed finger in a distal position in relation to the trichobothria et and est; the distance between external trichobothria of the femur, e1 and e2, being two to five times the distance between trichobothria e1 and d3 of the femur. In contrast, Reddyanus has trichobothrium db in a basal position to et, situated between et and est; the distance between external trichobothria of the femur, e1 and e2 is always less than twice the distance between e1 and d3.
Vachon [13] placed three species in the subgenus Isometrus and five in the subgenus Reddyanus, leaving five others without a precise assignment. In the catalog of the scorpions of the world, Fet & Lowe [14] placed six species in the subgenus Isometrus and all others (16) in the subgenus Reddyanus. In his revision of the genus, Kovařík [7] placed four species in the subgenus Isometrus and all others (21) in the subgenus Reddyanus. It would therefore seem that the subgenus Reddyanus is much more speciose than the subgenus Isometrus. Kovařík [7] described several new species, including one species, I. (R.) petrzelkai, from Vietnam and Thailand. The type material of the latter (deposited in the personal collection of the author) has not been available for study, but we have examined some specimens of Isometrus from Vietnam, formerly identified by L. Fage as Isometrus vittatus Pocock, 1900, which are conspecific with I. (R.) petrzelkai. This species and Isometrus (Reddyanus) hainanensis Lourenço, Qi & Zhu, 2005 appear to be closely related to the new species described here. Comparison of the coloration patterns of the three species shows significant differences. Subcuticular pigmentation can be very stable within buthid species populations [15,16] and therefore useful as a taxonomic character in the definition of species.
3 Buthid cave scorpions
Troglobitic scorpions (i.e., species which complete their entire biological cycle within caves) are rare across the world. It was not until the late 1960s that the first truly troglobitic scorpions were described from Mexican caves [17]. By the mid-1980s, the number of troglobite species had increased to thirteen [18]. Eleven belong to the families Diplocentridae Karsch, 1880, Euscorpiidae Laurie, 1896, Superstitioniidae Stahnke, 1940, and Vaejovidae Thorell, 1876. The other two belong to the families Troglotayosicidae Lourenço, 1998 and Chaerilidae Pocock, 1893. Subsequently, other troglobitic scorpions have been described, belonging to the families Superstitionidae and Chaerilidae. Exceptions are the recent descriptions of troglobitic species belonging to the families Liochelidae Fet & Bechly 2001, Pseudochactidae Gromov 1998, Urodacidae Pocock 1893 and Buthidae C. L. Koch 1837 [10,19–22].
Scorpions of the family Buthidae have also been found in caves. In most cases, however, these proved to be only trogloxenes (i.e., species which occasionally penetrate into caves, but have their biological cycle outside), or troglophilic elements (i.e., species that are regularly found outside caves). Examples are Alayotityus juraguaensis Armas, 1973, Tityus monaguensis González-Sponga, 1974, Tityus magnimanus Pocock, 1897, Tityus demangei Lourenço, 1981, Babycurus johnstonii Pocock, 1896 and more recently Lychas hosei cavernicola Lourenço, 2007 [18,23–25]. In their recent re-analysis of troglobitic scorpions Volschenk & Prendini [22] list only three buthid species. There are also some inadequacies in their analysis, since T. demangei was not considered by Lourenço [23] as a possible troglobitic element, only as a trogloxene. Unequivocal troglobitic characteristics have been observed for two buthid species: Troglotityobuthus gracilis (Fage, 1946) and Troglorhopalurus translucidus Lourenço, Cerqueira Baptista & Giupponi, 2004 [20,21]. The new species describe here can only be considered as a possible troglophile, since it was found both inside and outside the caves.
4 Methods
Illustrations and measurements were produced using a Wild M5 stereomicroscope with a drawing tube (camera lucida) and an ocular micrometer. Measurements follow Stahnke [26] and are given in mm. Trichobothrial notations follow Vachon [27] and morphological terminology mostly follows Hjelle [28].
5 Taxonomic treatment
Buthidae C. L. Koch, 1837
Isometrus Ehrenberg, 1828
Isometrus (Reddyanus) deharvengi sp. n. (Figs. 1 and 2)
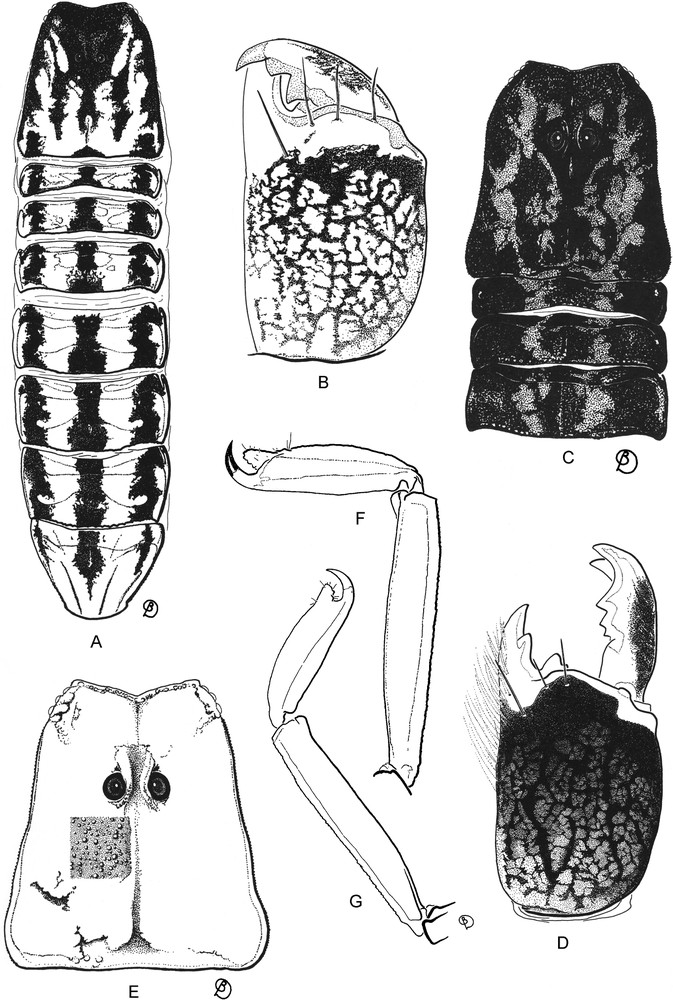
Isometrus (Reddyanus) deharvengi sp. n., male holotype (A,B,E,F) and Isometrus (Reddyanus) petrzelkai male from Vietnam (C,D,G). A to D. Pigmentation pattern on carapace, tergites and chelicera; figures C and D are interpreted from a poorly preserved specimen. E. Carapace; small square shows the tegumental granulation. F–G. Metasomal segment V and telson, lateral aspect.
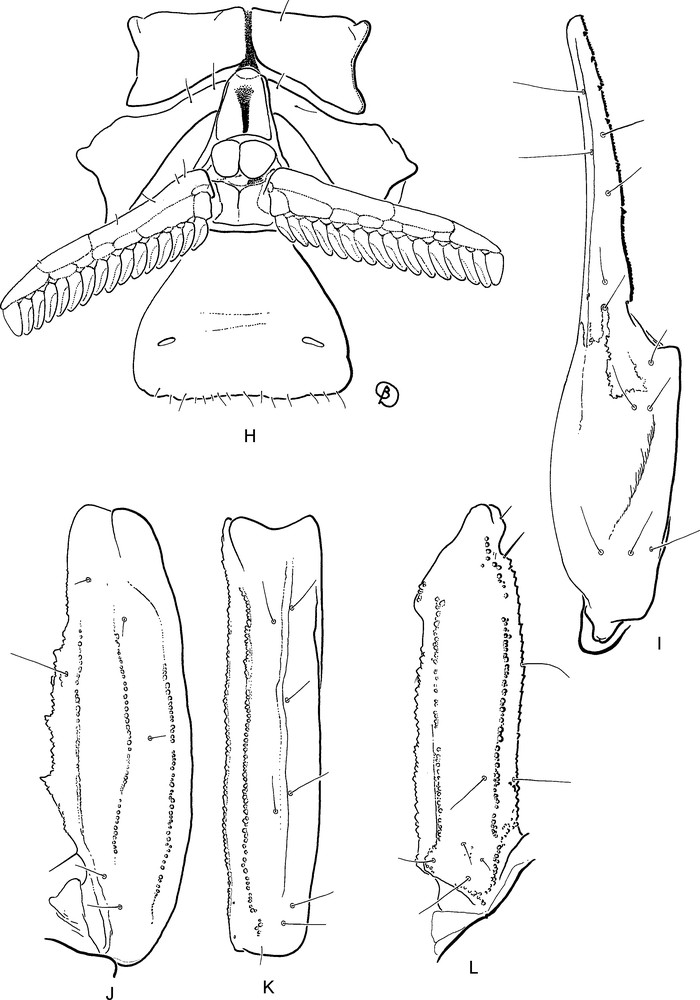
Isometrus (Reddyanus) deharvengi sp. n., male holotype. H. Ventral aspect showing coxapophysis, sternum, genital operculum, pectines and sternite III with spiracles. I–L. Trichobothrial pattern. I. Chela, dorso-external aspect. J–K. Patella, dorsal and external aspects. L. Femur, dorsal aspect.
Type material: Vietnam, Kien Giang, Kien Luong: Hon Chong, Nui Hon Chong hill: Hang Giêng Tiên, in cave (L. Deharveng, Truong Quang Tam & Duong Tien Dung), 8/11/1993, male holotype (VIET-010). Same locality, Nui Cay Ot hill, in litter (L. Deharveng & A. Bedos), 21/08/2003, one juvenile (Vn0308-066). Same locality, Nui Khoe La hill: ‘grotte sud’, in cave (L. Deharveng & A. Bedos), 1/06/2008, one male juvenile (Vn08-030). Same locality, Nui Bai Voi hill, ‘Grotte du Vieux’, in cave (L. Deharveng & A. Bedos), 2/06/2008, one female juvenile (Vn08-041). Same locality, Nui Bai Voi hill, ‘Grotte-Hopital’, in cave (L. Deharveng & A. Bedos), 4/06/2008, one male juvenile (Vn08-078).
Deposited in the collections of the Muséum national d’histoire naturelle, Paris.
Etymology: Patronym in honor of Dr. Louis Deharveng, CNRS/MNHN, one of the collectors of the new species.
5.1 Diagnosis
Isometrus species of moderate to large size, measuring up to 60 mm in adult male. General coloration yellowish to pale yellow in both adults and juveniles; several dark spots present, better marked in juveniles. Carinae and granulations moderately marked. Carapace strongly emarginate, with an open V-shaped angle. Pectines moderately long; pectinal tooth count 16/17 in males and 15/16 in female. Dentate margins of fixed and movable fingers of pedipalp chela with 7–6 almost linear rows of granules. Subaculear tubercle strongly developed and rhomboid, with 6/7 ventral granules.
Relationships: From its general morphology, Isometrus (Reddyanus) deharvengi sp. n. appears to be most closely related to I. (R.) petrzelkai, also described from Vietnam (80–100 km NNE from Saigon), and to I. (R.) hainanensis, described from the island of Hainan. It can be distinguished from these two species by the following characters: (i) a much paler coloration, in particular when compared to I. (R.) petrzelkai, which has a dominant blackish coloration over the carapace and tergites; (ii) the new species has a stronger subaculear tubercle; and (iii) the morphometric values are quite distinct between the new species and I. (R.) petrzelkai in particular (Table 1).
Morphometric values (in mm) of male holotype and female paratype of Isometrus (Reddyanus) hainanensis; male holotype of Isometrus (Reddyanus) petrzelkai Kovařík [7]; male holotype of Isometrus (Reddyanus) deharvengi sp. n.
| Isometrus (R.) hainanensis | I. (R.) petrzelkai | I. (R.) deharvengi sp. n. | ||
| Holotype | Paratype | Holotype | Holotype | |
| Total length | 53.4a | 34.2a | 47.0 | 60.7a |
| Carapace | ||||
| Length | 4.9 | 4.6 | 4.3 | 6.1 |
| Anterior width | 2.9 | 2.8 | – | 3.4 |
| Posterior width | 4.6 | 4.6 | 4.0 | 5.4 |
| Metasomal segment I | ||||
| Length | 4.7 | 3.1 | 3.7 | 6.0 |
| Width | 1.8 | 2.0 | 1.4 | 2.1 |
| Metasomal segment V | ||||
| Length | 8.5 | 5.4 | 7.4 | 9.8 |
| Width | 1.3 | 1.4 | 1.5 | 1.7 |
| Depth | 1.5 | 1.6 | – | 1.8 |
| Telson length | 6.4 | |||
| Width | 1.4 | 1.2 | – | 1.7 |
| Depth | 1.5 | 1.4 | – | 1.7 |
| Pedipalp | ||||
| Femur length | 4.9 | 4.0 | 4.2 | 5.8 |
| Femur width | 1.5 | 1.3 | 1.1 | 1.7 |
| Patella length | 5.5 | 4.7 | 4.7 | 6.6 |
| Patella width | 2.0 | 1.9 | 1.7 | 2.2 |
| Chela length | 8.4 | 7.4 | 7.8 | 10.7 |
| Chela width | 2.1 | 1.8 | 1.9 | 2.4 |
| Chela depth | 1.8 | 1.4 | – | 2.0 |
| Movable finger | ||||
| Length | 4.5 | 4.3 | 3.9 | 6.0 |
a The total length does not include the telson. If the length of the telson is added, the total length is 67.1 mm for the male holotype of I. (R.) deharvengi sp. n.
5.2 Description based on male holotype and paratypes. (Morphometric measurements in Table 1)
Coloration. Generally yellowish to pale yellow. Prosoma: Carapace yellowish with an inverted blackish-brown triangle on the anterior margin, extending from lateral eyes to behind median eyes; with five longitudinal blackish-brown stripes; one median running from the median eyes to the posterior margin; two central and two lateral; eyes surrounded by black pigment. Mesosoma: Tergites yellowish with three longitudinal blackish-brown stripes that continue the median and lateral stripes of the carapace. Metasomal segments yellowish, with some minute, diffuse, brownish spots. Vesicle yellowish; aculeus yellowish at base and dark reddish at tip. Venter pale yellow all over. Chelicerae pale yellow with brownish variegated spots; base of fingers pale yellow, rest of fingers yellowish and slightly infuscate, teeth reddish. Pedipalps yellowish, slightly infuscate; chela fingers reddish-brown; rows of granules on dentate margins of fingers dark reddish. Legs pale yellow with diffuse spots.
Morphology. Prosoma: Anterior margin of carapace strongly emarginate, with an open V-shaped angle. Carapace carinae weakly marked; anterior median and posterior median carinae weakly developed; central lateral carinae extending behind lateral eyes. Intercarinal spaces weakly to moderately granular. Median ocular tubercle anterior to the centre of the carapace; median eyes separated by one ocular diameter. Three pairs of lateral eyes. Mesosoma: Tergites I–VI with a median carina; weak on I, weak to moderate on II–VI. Tergite VII pentacarinate, with lateral pairs of carinae moderate to strong; median carinae present in proximal half, moderately developed. Intercarinal spaces moderately granular. Sternites III to V smooth; VI and VII with a weakly marked granulation; spiracles long; sternite VII with four carinae. Pectines moderately long; pectinal teeth count 16-16 in male holotype. Metasomal segments I and II with 10 carinae, crenulate; III–IV with 8 carinae, crenulate. Segment V with five carinae; one posterior spinoid granule on dorsal carinae of segments II–IV. Dorsal furrows of all segments weakly developed, smooth; intercarinal spaces very weakly granular to smooth. Telson very weakly granular, almost smooth with one vestigial ventral carina; subaculear tubercle very strongly marked and rhomboid, with 6/7 granules on the ventral surface. Chelicerae with dentition characteristic of buthids [29]; two small basal teeth on movable finger. Pedipalps: Femur pentacarinate; all carinae crenulate. Patella with nine carinae, crenulate; dorsointernal carinae with 4/5 spinoid granules. Chela with seven/eight weakly crenulated carinae. Intercarinal spaces moderately to weakly granular. Dentate margins on fixed and movable fingers composed of 7/6 linear rows of granules. Trichobothrial pattern type A, orthobothriotaxic [27]; dorsal trichobothria of femur in β (beta) configuration [30]. Legs: ventral aspect of tarsi with a brush-like group of setae. Tibial spurs absent; pedal spurs present and moderately developed on all legs.
Acknowledgements
We are most grateful to Truong Quang Tam from ITB Vietnam who organised the access to study site and to Louis Deharveng (CNRS-MNHN Paris) who provided the material for study, and to Mark Judson and Elise-Anne Leguin, (MNHN Paris) for the revision of the text and for assistance in the preparation of the plates.