

1 Introduction
As already mentioned in previous papers, the family Chaerilidae, with its single genus Chaerilus, remains poorly-known among extant scorpions. The genus Chaerilus Simon, 1877 was created to accommodate Chaerilus variegatus Simon 1877, a species originally described from Java. Several other new species were later described from the Indonesian islands, and nearby geographic regions such as Malaysia and Singapore. The majority of these species were described based on very limited material and on weak and/or unclear diagnostic characters. Moreover, in most cases these descriptions were not properly illustrated or not even illustrated at all. This confused situation has lead to several cases of misidentification by different authors [1–6].
More recently, attempts to clarify the taxonomic status of several species have been more or less successful. Kovařík [7] confirmed the identity of one population in Southern Vietnam (Indochina) previously misidentified by Fage [4–6], as Chaerilus celebensis Pocock, 1894, Chaerilus rectimanus Pocock, 1899 and/or Chaerilus variegatus. Kovařík [7] described this population as Chaerilus petrzelkai. Subsequently, this species was confirmed in Vietnam, and properly redescribed and illustrated by Lourenço and Zhu [8]. In the same way, Chaerilus celebensis was redescribed and confirmed to be an endemic element of the Celebes (Sulawesi Island). New associated species have also been described, namely Chaerilus philippinus Lourenço and Ythier, 2008 from the North of Luzon in the Philippines [9], Chaerilus laoticus Lourenço and Zhu, 2008 from Laos and Chaerilus vietnamicus Lourenço and Zhu, 2008 from the North of Vietnam [8], Chaerilus telnovi Lourenço, 2009 and Chaerilus spinatus Lourenço and Duhem, 2010 from the Moluccas Islands [10,11] and finally Chaerilus thai Lourenço, Sun and Zhu, 2010 from South of Thailand [12].
Although some progress has been made in the study of these tropical species of Chaerilus, the status of some species described long ago, still remains unclear. This was true of C. variegatus and C. borneensis Simon, 1880, described by Simon [13,14], from Java and Borneo and considered to be synonymous since the time of Kraepelin [1]. In a recent note, however, the necessary justification was proposed to establish that both species are valid and distinct [15].
Until now, only two Chaerilus species have been discovered in Vietnam: Chaerilus petrzelkai from the South region of the country and Chaerilus vietnamicus from the North range [8]. An additional new species is described here from the South of Vietnam.
2 A division of the genus Chaerilus in species-groups
In the Catalog of Scorpions of the World [16], 21 species have been listed in the genus Chaerilus Simon, 1877. In his attempt to revise the genus, Kovařík [7] defined 18 species as valid. During the last decade, however, numerous additions to the species list of Chaerilus have been published, and again the total number of valid species has proved to be a matter of disagreement among authors [17]. In two recent syntheses proposed by Di and Zhu [18] and Di et al. [19], respectively 27 and 28 valid species were listed. The difference between these numbers comes from the fact that, in the first list the poorly known Indian species Chaerilus dibangvalleycus Bastawade, 2006 was not recognized. In both these publications, however, the authors continued to ignore the validity of Chaerilus borneensis. The validity of this species was finally re-established by Lourenço et al. [15]. Even more recently, other new species have been added to the genus Chaerilus, namely, Chaerilus mainlingensis Di and Zhu, 2009 from China, Chaerilus annapurna Lourenço and Duhem, 2010 from Nepal, Chaerilus thai from Thailand and Chaerilus spinatus from Halmahera Island (Moluccas), Indonesia [20]. This raises the total number of known valid species to 33.
It is obvious that the species of the genus Chaerilus do not form a homogeneous group, but are composed of at least two main sub-groups. Even so, no author has suggested a possible division of the genus in species-groups. Contrary to the idea of genus and subgenus which should be used as compulsory categories, that of the species-group remains informal. In this publication, two species-groups are proposed: the ‘variegatus species-group’, and the ‘truncatus species group’.
The ‘variegatus species-group’ contains species of small to medium size (usually from 20 to 40 mm in length), presenting a generally yellow to reddish-yellow coloration and a very intense dark variegated pigmentation over the body and appendages. The majority of species in this group are distributed in tropical and subtropical regions of Asia. The following are integrate within it: C. borneensis, C. celebensis, C. laoticus, C. ojangureni Kovarik, 2005, C. petrzelkai, C. philippinus, C. rectimanus, C. robinsoni Hirst, 1911, C. sabinae Lourenço, 1995, C. sejnai Kovarik, 2005, C. spinatus, C. telnovi, C. thai, C. variegatus and C. vietnamicus.
The ‘truncatus species-group’ comprises species of medium to large size (usually from 40 to 75 mm in length), presenting a dark coloration over all, that extends from brownish to almost blackish. Many of these are also distributed in tropical regions, but a significant number inhabit regions of India, Nepal and China that are mesic temperate to cold. The following species are integrated with this group: C. agilis Pocock, 1899, C. annapurna, C. assamensis Kraepelin, 1913, C. cavernicola Pocock, 1894, C. ceylonensis Pocock, 1894, C. chapmani Vachon and Lourenço, 1985, C. conchiformus Zhu, Han Lourenço, 2008, C. dibangvalleycus, C. insignis Pocock, 1894, C. laevimanus Pocock, 1899, C. lehtrarensis Khatoon, 1999, C. mainlingensis, C. pictus, C. tessellatus QI, Zhu and Lourenço, 2005, C. tichyi Kovarik, 2000, C. tricostatus Pocock, 1899, C. truncatus Karsch, 1879, C. tryznai Kovarik, 2000 and C. julietteae sp. n.
3 Methods
Illustrations and measurements were made with the aid of a Wild M5 stereo-microscope which has a drawing tube (camara lucida) and an ocular micrometer. Measurements follow Stahnke [21] and are given in millimeter (mm). Trichobothrial notations follow Vachon [22], and morphological terminology mostly follows Hjelle [23].
4 Taxonomic treatment
Chaerilidae Pocock, 1893.
Chaerilus Simon, 1877.
Chaerilus julietteae sp. n. (Figs. 1 and 2)
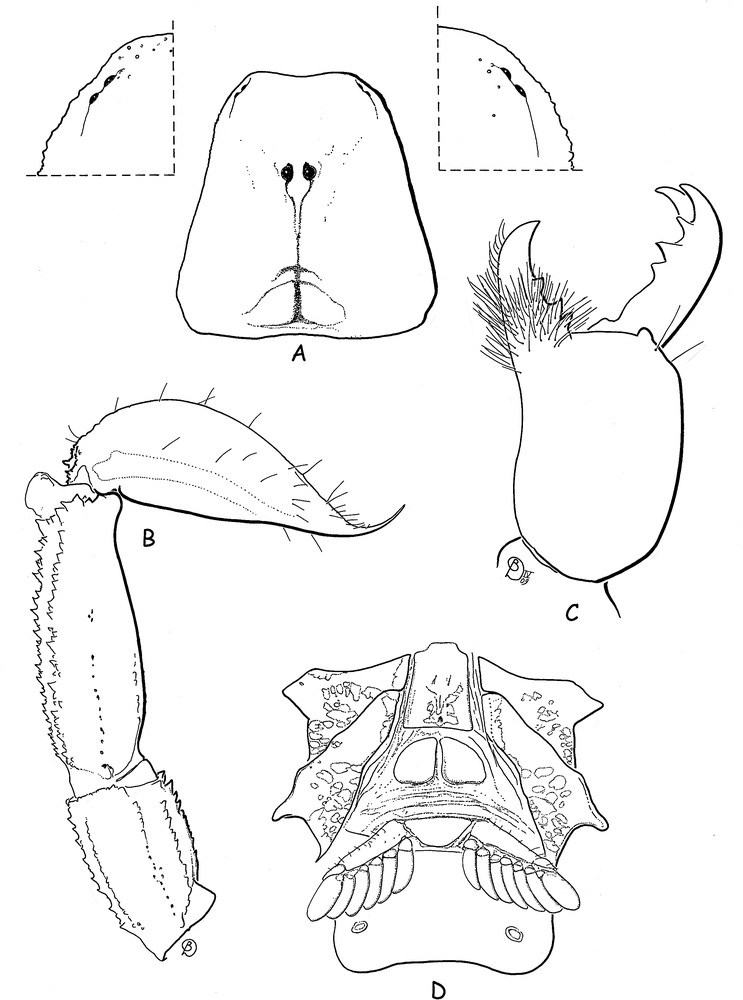
Chaerilus julietteae sp. n. Male holotype. A. Carapace and lateral eyes in detail. B. Metasomal segments IV-V and telson, lateral aspect. C. Chelicera, dorsal aspect. D. Ventral aspect, showing coxapophysis, sternum, genital operculum and pectines.
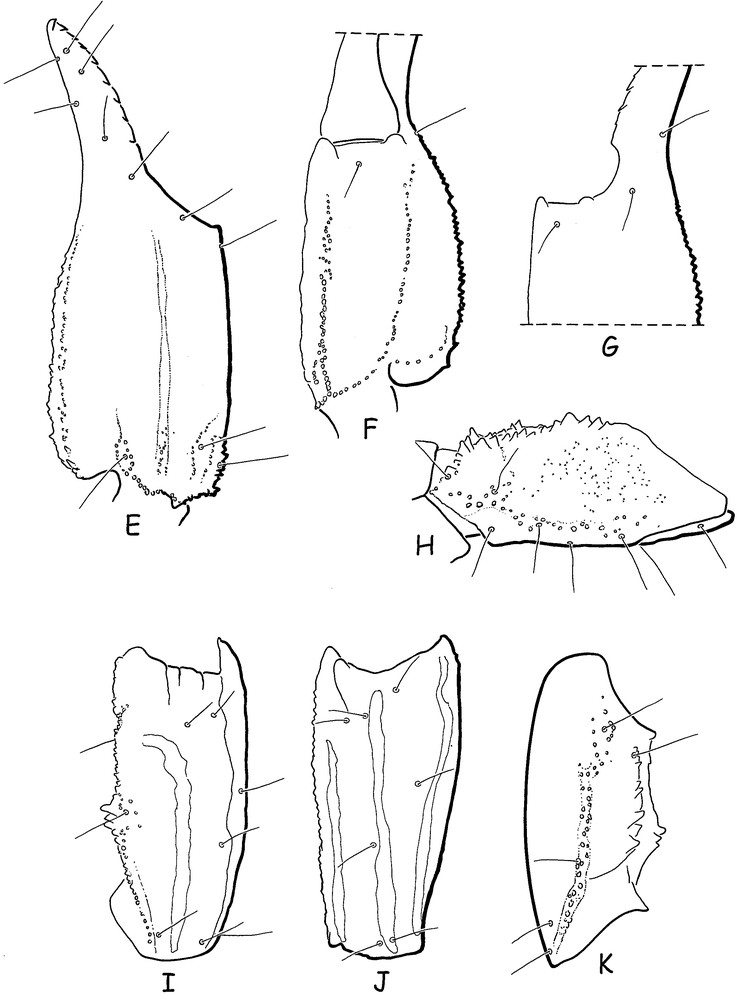
Chaerilus julietteae sp. n. Male holotype. Trichobothrial pattern. E-G. Chela, dorso-external, ventral and internal aspects. H. Femur, dorsal aspect. I-K. Patella, dorsal, external and ventral aspects.
Diagnosis: species of moderate size in relation to the other species of the genus which reach 46 mm in total length. Carapace strongly narrowed toward the anterior edge in males, less narrowed in females; almost acarinate and weakly granulated to smooth; anterior margin straight; furrows shallow in males, moderately deep in females. Metasomal carinae strongly marked; ventral carinae reduced on segment I, moderately marked on segment II; latero-ventral and ventral carinae on segments IV-V composed of strong spinoid granules; other carinae with moderately marked spinoid granules. Dentate margins of fixed and movable fingers of pedipalp chela with 10-10 rows of granules. Pectinal teeth count 7-6 in male, 5-5 in female. Genital operculum plates have a sub-oval shape. Trichobothriotaxy of type B, orthobothriotaxic.
Relationships: Chaerilus julietteae sp. n., shows morphological structures similar to those of Chaerilus pictus described from Bangladesh. The new species can, however, be readily distinguished from it by the following features: (i) the shape and structure of the carapace, (ii) a small number of rows of granules in chela fingers, (iii) the larger number of pectinal teeth, (iv) metasomal carinae armed with very strong spinoid granules.
Etymologie: patronym in honor of Dr. Juliette Arabi who collected the type material.
Material: Vietnam, Binh Thuan province, Ta Kou, 672 m alt. (under a stone), 13/XII/2006 (leg J. Arabi). Male holotype; one juvenile female paratype. Type material deposited in the Muséum national d’Histoire naturelle, Paris (MNHN-RS-8846-8847).
4.1 Description
Coloration: basically blackish to blackish-brown. Carapace blackish. Tergites with the same colour as the carapace and some diffused confluent yellowish pigmentation. Metasoma: All segments blackish with yellowish spots laterally; carinae dark to blackish. Telson blackish with lateral yellowish stripes; aculeus reddish-yellow. Chelicerae yellowish with blackish variegated spots; fingers blackish with reddish teeth. Pedipalps: blackish; chela fingers with the extremity reddish. Legs blackish-brown with diffused yellowish spots. Venter and sternites yellowish with variegated blackish-brown spots; pectines pale yellow.
Morphology: carapace strongly narrowed anteriorly in males, less narrowed in females; anterior margin straight, almost acarinate and weakly granular to smooth; furrows shallow in males, moderately deep in females. Two pairs of lateral eyes, and a pair of moderate median eyes, about three times the size of the lateral eyes; median eyes anterior to the centre of the carapace. Tergites with only minute granulations in both males and females; carinae obsolete. Sternum pentagonal, longer than wide; genital plates of operculum sub-oval in shape. Pectinal teeth count 7-6 in male holotype, 5-5 in female paratype. Sternites smooth with spiracles small and oval-shaped; carinae absent from VII. Metasoma: Segments I and II wider than long; III as long as wide; segments IV and V longer than wide. All the carinae strongly granular; ventral carinae reduced on I, moderaly marked on II; segments IV and V with latero-ventral and ventral carinae composed of strong spinoid granules. Vesicle very elongated with a pear-like shape, smooth, with a short aculeus. Pedipalps: Femur with five carinae; internal having spinoid granules. Patella with seven carinae; dorsal and dorso-external weakly granular; dorso and basal-internal with spinoid granules. Chela with eight carinae, moderately to strongly granular; ventral median carinae weak. Tegument with few granulations. Finger shorter than manus with 10-10 rows of granulations on the dentate margins. Chelicerae characteristic of the family Chaerilidae [24]. Trichobothriotaxy type B; orthobothriotaxic [22]; femur with 9 trichobothria, patella with 14, and chela with 14. Legs with pedal spurs moderately developed. Tarsi with two rows of spiniform setae.
Morphometric values (in mm) of the male holotype. Total length, 46.2 (including telson). Carapace: length, 7.6; anterior width, 3.9; posterior width, 7.7. Mesosoma length, 14.0. Metasomal segments. I: length, 2.3; width, 3.6; II: length, 2.8; width, 3.2; III: length, 2.9; width, 2.9; IV: length, 3.3; width, 2.5; V: length, 6.2; width, 2.4; depth, 2.3. Telson length, 7.1. Vesicle: width, 2.5; depth, 2.3. Pedipalp: femur length, 6.7, width, 2.7; patella length, 6.8, width, 2.8; chela length, 13.8, width, 4.4, depth, 4.6; movable finger length, 6.9.
Disclosure of interest
The author declares that he has no conflicts of interest concerning this article.
Acknowledgements
The new species was collected during the project ‘Cordillère Annamitique’ coordinated by Louis Deharveng (MNHN) and Truong Quang Tam (ITB, Ho Chi Minh City). The project was funded by the PPF ‘État et structure de la biodiversité actuelle et fossile’ of the Muséum national d’Histoire naturelle of Paris. The field trip to Takou nature reserve was organized by our colleague Dr Luu Hong Truong from CDB-ITB (Ho Chi Minh City).
My thanks go also to Bernard Duhem, (MNHN, Paris) who prepared the illustrations; Elise-Anne Leguin, (MNHN, Paris) for the preparation of the plates and to Prof. John L. Cloudsley-Thompson, London for comments on and revision of the manuscript.