

1 Introduction
The phasmatodean genus Ophicrania Kaup, 1871 is classified within the subfamily Platycraninae (Phasmatodea: Phasmatidae) [1], and currently encompasses 19 described species [2]. Recent evidence on the phylogenetic relationships among phasmatodean taxa, based upon the analysis of morphological [3] and molecular [4] data, suggest that Ophicrania belongs to a large clade of mainly Australasian taxa, named Lanceocercata by Bradler [5]. The geographical distribution of the genus is rather wide, and ranges from Malaysia and Indonesia to Papua New Guinea and the Philippines.
A checklist of the Philippine members of Ophicrania was recently provided by Bresseel and Bushell [6], whom reported nine species from the islands of Luzon, Samar and Mindanao. In the present study, an additional distinctive species is described from Panay Island as new for science, along with an emended diagnosis of the genus, and a summary of the Philippine taxa of Platycraninae. This article is a contribution to the series “Systematics of Philippine Phasmatodea” [7–10].
2 Materials and methods
The specimens of Ophicrania studied were dried and pinned. Photographs of the habitus were taken with a Nikon D50 digital camera. To observe the external features of male terminalia, the distal abdominal segments where briefly rehydrated and cleaned in 10% KOH. Morphological observations were carried out using a Zeiss Stemi DV4 stereo light microscope, and drawings were done with the aid of a camera lucida. For the scanning electron microscopy (SEM) study, legs were cleaned in 10% KOH, briefly sonicated, dried in a stove at 60 °C, mounted on an aluminium stub using double stick adhesive, gold-coated in a Balzers MED 010 sputtering device, and observed with a Philips XL20 scanning electron microscope operated at 10 kV. The following letter codens are used to designate the collections: ZSM: Zoologische Staatssammlung München, Germany; JB: personal collection of Joachim Bresseel, Meise, Belgium; MG: personal collection of Marco Gottardo, Rovigo, Italy; MNCN: Museo Nacional de Ciencias Naturales, Madrid, Spain.
3 Systematics
Order Phasmatodea Jacobson and Bianchi, 1902
Family Phasmatidae Gray, 1835
Subfamily Platycraninae Brunner von Wattenwyl, 1893
Genus Ophicrania Kaup, 1871
3.1 Diagnosis of the genus
Members of Ophicrania include medium-sized Platycraninae with a rather slender and smooth body in both sexes. In this genus the prognathous head is distinctly longer than wide, and somewhat cylindrical in cross-section. The vertex is flat or moderately rounded. The antennae are short and filiform. Ocelli are lacking. The ventral surface of the head features a well-developed gula. The prothorax is slightly longer than wide, and is characterized by the presence of pronotal sides (or paranota), and the fusion of the ana- and coxopleurite in the pleural region. The mesothorax is elongated and slender; the mesonotum is rather tectiform and features a slight longitudinal median furrow, and a longitudinal keel on each lateral margin. The genus presently includes macropterous, brachypterous, micropterous, and apterous species. In winged species, the tegmina are small, significantly shorter than the mesonotum. The femora are trapezoidal in cross-section, and feature five carinae. The dorsal carinae, and the medio-ventral carina are rather indistinctly developed in the mid and hind femora. The medio-ventral surface of the femora is armed distally with small spines, whose number varies among the different species. The basal portion of the fore femur is compressed and slightly incurved. The tibiae are almost triangular in cross-section, as the dorsal surface is reduced and the dorsal carinae are close to each other. In the tarsi, the basitarsus in elongated, distinctly longer than combined length of tarsomeres II-III. The male genital region exhibits a short sternite IX, which hardly extends over the abdominal tergite IX. The sternite IX also has a transverse furrow, approximately at the anterior third, which divides the plate into a short anterior portion, and a more elongated posterior portion (or poculum). The tergite X is slightly tectiform. Its hind margin is strongly thickened and shows two symmetrical claspers, which are armed ventrally with well-developed toothlike projections (or thorn pads). The vomer is lacking. In the female genital region, the hind margin of sternite VII exhibits an indistinctly developed preopercular organ. The sternite VIII (the female subgenital plate) is boat-shaped, and lacks a terminal notch on the hind margin. Its length varies among the species: it may be short, when does not extend over the tergite IX; medium, when reaches the hind margin of tergite X; long, when distinctly project beyond the hind margin of tergite X. The ovipositor extends beyond the sternite VIII when the latter is of the short type. In both sexes the cercus is dorso-ventrally flattened, much longer than wide, and with a pointed apex. The egg capsule is elongated (height/length% = 50) and laterally flattened (width/height% = 90–95). The operculum features a peg-like capitulum. The micropylar plate is represented by a small drop-shaped mark at the posterior half of the egg capsule. The median line is present.
Type species Ophicrania striatocollis Kaup, 1871, by original monotypy.
3.2 Description of the new species
Ophicrania conlei, n. sp. (Figs. 1–3; Table 1).

Ophicrania conlei n. sp., holotype male. (A) Habitus, lateral view. (B) Head, pro- and mesothoracic segments, lateral view.

Ophicrania conlei n. sp., holotype male, scanning electron micrographs. (A) Distal portion of the mid femur, lateral view. (B) Dorsolateral surface of the mid femur. (C) Detailed view of the two spines on medio-ventral surface of the mid femur. DS, dorsal surface; PLS, posterior lateral surface; VPC, ventro-posterior carina; MVS, medio-ventral surface.
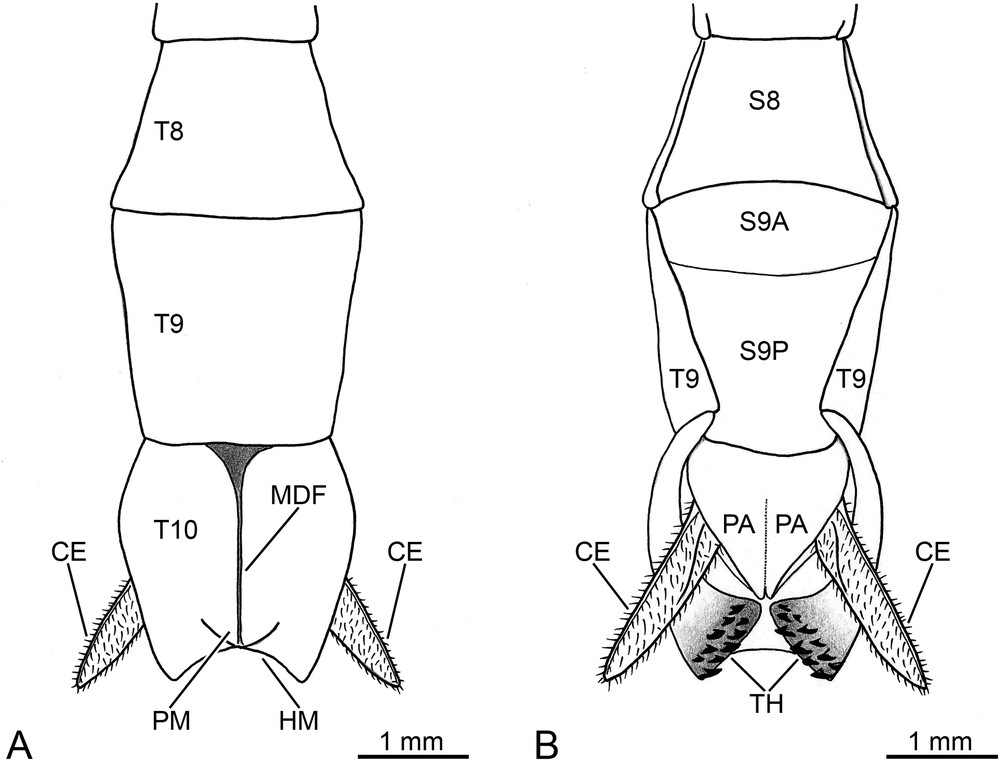
Ophicrania conlei n. sp., holotype male, schematic drawings of terminalia. (A) dorsal view. (B) ventral view. CE, cercus; HM, hind margin of tergite X; MDF, mid-dorsal furrow of tergite X; PA, paraproct; PM, posteromedian mound of tergite X; S8, abdominal sternite 8; S9A, anterior portion of sternite IX; S9P, posterior portion of sternite IX (or poculum); T8-T10, abdominal tergites 8-10; TH, thorns.
Morphometric data for Ophicrania conlei n. sp. from Panay island, Philippines.
| Parameter | Male holotype |
| (ZSM) | |
| Measurement (mm) | |
| Body length | 54.3 |
| Antenna length | 16.6 |
| Head length | 3.0 |
| Pronotum length (PL) | 2.6 |
| Mesonotum length (MOL) | 8.0 |
| Metanotum length (MAL) | 6.4 |
| Median segment length (MSL) | 3.2 |
| Tegmina length | 5.8 |
| Hind wing length | 26.3 |
| Cercus length | 2.0 |
| Fore femur length | 17.1 |
| Fore tibia length | 17.8 |
| Mid femur length | 10.2 |
| Mid tibia length | 9.4 |
| Hind femur length | 11.9 |
| Hind tibia length | 12.3 |
| Morphometric ratios | |
| MOL divided by PL | 3.08 |
| MAL divided by MOL | 0.80 |
| MSL divided by MAL | 0.50 |
Type material: Holotype ♂: Philippines, Panay Island, Mt. Madja-as, Culasi Antique, X.2008, leg. N. Mohagan, ex coll. O. Conle (ZSM).
Comparative material: Ophicrania nigrotaeniatus (Redtenbacher, 1908): ♂, Philippines, Valle de Bulasan (MNCN) (photographs examined). Ophicrania sagittarius Bresseel and Bushell, 2009: ♂ paratype, Philippines, Mindanao Island, Mt. Apo, 1300 m, 27.III-10.IV.2006, leg. R. Cabale (MG). Ophicrania stygius (Westwood, 1859): ♂, Philippines, Luzon, Infanta, VII.2009, leg. Bresseel & Van Dingenen (JB). Ophicrania viridinervis (Stål, 1875): ♂, Philippines, Luzon, Infanta, VII.2009, leg. Bresseel & Van Dingenen (JB). Photographs of type material of the remaining species of Ophicrania have been examined at the Phasmida Species File Online [2].
Diagnosis: a medium-sized species of Ophicrania, characterized by the short antennae, rather slender femora, well developed wings featuring the anal region of the male hind wing divided into an inner whitish area, and an outer brownish area. The body is slender, and the integument is weakly setose and slightly glossy (Fig. 1A).
Morphology: the head (Fig. 1B) is stubby in structure, and moderately narrows towards base. Its colour is mainly pale green, with dark green interocular space. The dorsal surface of head features a rather flat vertex. The gena is approximately one and a half times broader than the greatest diameter of the compound eye. The antenna is evenly dark brown, shorter than fore femur, and composed of 24 antennomeres covered with short sensilla and setae. The scape is rectangular and dorsoventrally flattened, two times longer than wide. The pedicel is slightly wider than long, cylindrical, about half the length of scape. The first flagellomere is about twice as long as the pedicel. The compound eye is light brown, circular and prominent. The pentamerous maxillary palpus, and the trimerous labial palpus are brown. The gula is much longer than wide.
The prothorax (Fig. 1B) is narrower and shorter than head. The pronotum is smooth, yellowish, with a dark brown median longitudinal band; it is rather flat, and possesses a faint cross-shaped incision in the center. The anterior and lateral margins of pronotum feature a distinctive edge; moreover, both anterolateral corners are equipped with an impressed ellipsoid-like glandular area, which supports the opening of the prothoracic exocrine gland. The posterior margin of the pronotum is simple, very moderately rounded. The pronotal sides (or paranota) are well developed, and cover the posterior two-thirds of the pleural region. The prothoracic coxopleurite is yellowish and rather rectangular. The prothoracic basisternite is cup-shaped, evenly yellowish. The mesothorax (Fig. 1A-B) is moderately elongated and slightly constricted towards the posterior third. The mesonotum is tectiform, yellowish, with a longitudinal dark brown median furrow delimited laterally by faint keels; its surface is finely rugulose, with some very small granules. The lateral margin of the mesonotum is charaterized by the presence of a distinct longitudinal keel. The mesothoracic episternum is narrow, approximately as long as the mesonotum; the epimeron is sharply triangular, one-third the length of the episternum; the two sclerites, which are separated by a faint pleural suture, are light brown in colour. The mesothoracic basisternite is elongated, slightly rugulose, also light brown; the furcasternite is much shorther, and moderately widens posteriorly. The metathorax is parallel-sided, longer and broader than the mesothorax. The metanotum is shorter than mesonotum; its surface is yellowish white and smooth. The metathoracic pleurite is light brown, and consists of a large rectangular episternum, and a narrow epimeron, which is displaced on the dorsal surface of the metathorax, alongside the median segment (or first abdominal tergite). The metathoracic sternite is represented by an elongated basisternite, and a short furcasternite, which regularly narrows posteriorly.
The tegmina (Fig. 1A) are mainly dark brown, with a broad longitudinal pale brown stripe. It is slender, with a small knob-like dorsal eversion on the proximal third; the posterior margin is angularly truncated, and reaches the anterior third of the first abdominal tergite. The hind wing (Fig. 1A) is well developed, and its posterior tip reaches the anterior quarter of abdominal tergite VI. The costal area is pale brown, whilst the remainder of the costal region is dark brown with pale brown longitudinal veins. The anal region features a transparent inner area and a translucent brown distal marginal area.
The legs (Fig. 1A) are green, with the carinae finely setose. The hind legs reach only the posterior margin of abdominal tergite IX. The coxae are unarmed. Trochanters are shorter than the corresponding coxae, and are fused to femora. The fore femur exhibits a pale green basal portion, which is compressed and very moderately incurved as typical of the genus. The dorsal and ventral carinae of fore femur are unarmed, whilst the distinct medio-ventral carina is armed with two very small distal spines. The mid and hind femora show a rather flattened dorsal surface, which lacks distinct dorsal carinae (Fig. 2A-B); the ventro-anterior and ventro-posterior carinae are prominent and unarmed (Fig. 2A); the medio-ventral carina is somewhat indistinct, but the surface is strongly convex, and features two weakly developed distal spines (Fig. 2A,C). The tibiae are unarmed and slender. The five-segmented tarsus is approximately half the length of the corresponding tibia. In the fore tarsi, the basitarsus is longer than combined length of tarsomeres II-IV; it is shorther in the case of the mid and hind tarsi. Tarsomeres I-IV are increasingly shorter. The ventral surface of tarsomeres I-IV exhibits a small euplantula at the distal end. Each euplantula is divided medially by a longitudinal furrow. The pretarsus, which is much shorter than combined length of tarsomeres I-IV, significantly widens towards the distal end, as typical of all phasmatodeans. It lacks an euplantula on the ventral surface, but features a distinct median longitudinal furrow. The pretarsal claws are symmetrical, and lack pectination on the ventral lateral keels. The arolium is rounded and broad.
The abdomen (Fig. 1A) is mainly brown in colour. The median segment is longer than wide, half the length of metanotum. The abdominal tergites II-III exhibit equal length; tergites IV-V are also equal in length, but slightly shorter than previous ones; tergites VI-VIII are progressively shorter; tergite IX is one and a half times the length of VIII. The tergites II-VII are of uniform width, tergite VIII widens towards the posterior end, whilst tergite IX narrows posteriorly. The tergite X (or anal segment) is green (Fig. 1A), tectiform, and features a raised mid-dorsal furrow, which terminates with a moderately prominent posteromedian mound (Fig. 3A). It is shorter than tergite IX, and shows the greatest width at the end of the anterior third, whilst the proximal and distal portions are distinctly narrower. In dorsal view, the hind margin shows a broad semicircular notch (Fig. 3A). The lateral margins are increasingly thickened from base to apex (Fig. 3B). Ventrally, at the level of the posterolateral portion, the thickened margins differentiate into two symmetrical claspers, each equipped with a group of about 10 toothlike projections of various size (Fig. 3B). These thorns are strongly sclerotized, and feature a rather flat and rounded apex orientated towards the ventral mid-line. The paraprocts (Fig. 3B) are rather elongated, and separated medially by a very faint longitudinal furrow. Their outer margin incurves in the posterior half, and progressively tapers to form a pointed apex. The epiproct is indistinctly developed, and does not extend beyond the hind margin of tergite X (not shown). The cercus (Fig. 3A-B) is slightly longer than tergite X; it is laterally flattened, covered with strong setae and short sensilla, and tapers towards a pointed apex. The sternite IX (or subgenital plate) (Fig. 3B) is twice as long as sternite VIII. It is moderately convex, and transversely divided in the anterior quarter. The hind margin of sternite IX is slightly rounded, and indistinctly exceeds the posterior end of the tergite IX.
Etymology: the specific name is a patronymic dedicated to Oskar V. Conle (Bosterlang, Germany), in recognition of his contributions to the knowledge of the Phasmatodea.
Geographic distribution: this new species is only known from the type locality, Mount Madja-as, Antique province, Panay Island, Philippines. It marks the first record of the genus Ophicrania from Panay island, and represents the second phasmatodean known to occur on Mount Madja-as, where only the aschiphasmatine Dinophasma maalon Gottardo, 2007 has been previously reported [7].
Remarks: the female and egg of this new species are unknown.
4 Discussion
The most important characters of O. conlei n. sp. can be listed as follow: (a) green head capsule, which is also fairly stubby in structure; (b) short antenna, formed by 24 antennomeres; (c) yellowish pro- and mesothorax, further characterized by a longitudinal dark brown colouration on the nota; (d) tegmina and the costal region of the hind wing with a pale brown external lateral colouration; (e) anal region of the hind wing with a transparent inner area, and a translucent brown distal marginal area; (f) unarmed dorso-anterior carina of fore femur; (g) mid and hind femora with the dorsal surface flattened; (h) medio-ventral surface of mid and hind femora with only two weakly developed distal spines; (i) shape of the tergite X, particularly of its hind margin, which features a broad semicircular notch; (j) claspers of tergite X equipped with a total of approximately 20 toothlike projections on the ventral surface.
Among the Philippine members of Ophicrania, a green colouration on the head of the male is also found in O. viridinervis (Stål, 1875). However, in this species only the interocular space features a vivid green patch, whilst the remainder of the dorsal and lateral surfaces of the head are black. Regarding the males of the remaining Philippine species, the head is evenly greenish-blue in the apterous O. sagittarius Bresseel & Bushell, 2009, black and reddish in O. brunni (Redtenbacher, 1908), orange with a black longitudinal stripe in O. nigricornis (Stål, 1877), evenly pale brown in O. nigrotaeniatus (Redtenbacher, 1908), black with yellow interocular space in O. stygius (Westwood, 1859) and O. vittipennis (Stål, 1875). The colour pattern of the head appears to be a distinctive and constant character in each specie; therefore it is highly informative for their correct recognition. O. conlei n. sp. and O. viridinervis also share the stubby structure of the head capsule, which is an uncommon character state in this genus. In fact, within Ophicrania the head is most commonly strongly elongate-oblong, a feature widespread also in other genera of Platycraninae.
The yellowish colouration of the pro- and mesothorax is apparently a distinctive character of O. conlei n. sp., whilst the presence of a median longitudinal dark band on the pro- and mesonotum of the male is a feature also found in several other Philippine species, for instance O. nigricornis, O. nigrotaeniatus, O. stygius, O. vittipennis, and again O. viridinervis. This dark-colored band may be more or less developed depending on the species, and probably is subject to intraspecific variation.
The presence of a pale brown outer stripe on the tegmina and costal region of the hind wing, is a character distinctly developed also in the male of O. nigrotaeniatus, and could be a possible synapomorphy of these two species. Another important wing feature is represented by the bicoloured anal region, which might be considered an autapomorphy of O. conlei n. sp., as it does not occurs in the other known species of Ophicrania.
The absence of spine-like serrations on the dorso-anterior carina of fore femur, along with the weakly developed spination on the medio-ventral surface of the mid and hind femora, are characteristics shared by most species of this genus, with the notable exception of O. nigroplagiatus (Redtenbacher, 1908) and O. nigrotaeniatus, where leg spination is more developed.
The structure of the male abdominal segment X exhibits mostly specializations of O. conlei n. sp. (character states J and K), not found in the remaining species.
The evidence presented above suggest that O. conlei n. sp. is apparently most similar to O. viridinervis and O. nigrotaeniatus. From both species, it can be easily recognized by significant features of the integumental coloration, antennae, wings, legs, and terminalia. In particular, O. viridinervis features the antenna composed of 21 antennomeres (in O. conlei n. sp. 24 antennomeres); the hind margin of tergite X lacks a notch (in O. conlei n. sp. it is notched, semicircular), and exhibits small claspers equipped with a total of 10 toothlike projections on the ventral surface (in O. conlei n. sp. the claspers are larger, armed with a total of about 20 toothlike projections). In O. nigrotaeniatus the hind margin of the poculum reaches the posterior two-thirds of tergite IX (in O. conlei n. sp. indistinctly projects beyond tergite IX), and the cercus is strongly elongated, twice as long as tergite X (in O. conlei n. sp. it is short, only moderately longer than tergite X).
Ophicrania, now with 10 described species from the Philippines (Table 2), represents the most speciose genus of Platycraninae reported from the archipelago. Current evidence suggests that all the species are endemics. In fact, the presence of O. nigricornis in Sabah and Sarawak (Malaysian Borneo) is regarded as doubtful [6,11]. The majority of the species can be easily separated on the basis of morphological characters, with the exception of O. palinurus (Westwood, 1859), O. stygius, and O. vittipennis. All three appear to be very closely related, morphologically similar species and are apparently restricted to Luzon Island [6]. Females of O. palinurus and O. stygius are characterized by a strongly elongate-oblong head, frail femora, slender tegminae reaching half-length of median segment, and well developed hind wings reaching the end of abdominal segment IV. Males of O. stygius and O. vittipennis share the same characteristics of the females concerning to the structure of the head and femora, and furthermore feature long hind wings reaching the abdominal tergite VI, a black head with a distinctive yellow spot within the interocular space, a broad longitudinal dark band on the pro- and mesonotum, and the evenly brown anal region of the hind wings. A taxonomic reassessment of these species is required in order to identify a pattern of distinguishing features, and/or the existence of new synonyms.
Checklist of Philippine Platycraninae (Phasmatodea: Phasmatidae).
| Species | Sexes known | Egg known | Philippine records |
| Megacrania alpheus (Westwood, 1859) | ♀ | – | “Philippines” |
| Megacrania batesii Kirby, 1896 | ♂♀ | X | LU, PA, RO |
| Ophicrania brunni (Redtenbacher, 1908) | ♂ | – | SA |
| Ophicrania conlei Gottardo n. sp. | ♂ | – | PA |
| Ophicrania nigricornis (Stål, 1877) | ♂♀ | – | “Philippines” |
| Ophicrania nigroplagiatus (Redtenbacher, 1908) | ♀ | – | LU |
| Ophicrania nigrotaeniatus (Redtenbacher, 1908) | ♂ | – | LU |
| Ophicrania palinurus (Westwood, 1859) | ♀ | – | LU |
| Ophicrania sagittarius Bresseel & Bushell, 2009 | ♂♀ | X | MA |
| Ophicrania stygius (Westwood, 1859) | ♂♀ | – | LU |
| Ophicrania viridinervis (Stål, 1875) | ♂ | – | LU |
| Ophicrania vittipennis (Stål, 1875) | ♂ | – | LU |
The Platycraninae are also represented in the Philippines by the genus Megacrania Kaup, 1871 [12–15] (Table 2), where it is, however, very scarcely documented. Megacrania batesii Kirby, 1896, a species primarily distributed in Australia, New Guinea, and the Solomon Islands, has been reported from the Philippines by Hsiung [14] based on few female nymphs, and by Lit and Eusebio [15] only based on “one specimen collected in 1958 on pandan”. A second species, Megacrania alpheus (Westwood, 1859), known only from the female lectotype (type locality: Sri Lanka, believed to be an erroneous locality datum), is only hypothetically regarded as a Philippine species [14].
The data summarized here emphasize that the current state of knowledge about Philippine Platycraninae is far from complete. The genus Ophicrania appears to have undergone an extensive evolutionary radiation in the Philippines, where several well-differentiated species are distributed on the various islands. Contrarily, the presence of the genus Megacrania requires confirmation, and more material is needed for further assessment.
Disclosure of interest
The author declares that he has no conflicts of interest concerning this article.
Acknowledgements
The author is grateful to his colleagues Joachim Bresseel, who loaned specimens of Ophicrania from his collection; Oskar V. Conle, who has made available the holotype of O. conlei n. sp. for study, and gave useful comments on the manuscript; and Frank H. Hennemann, who has provided photographs of O. nigrotaeniatus.