

Abridged English version
The palaeoecological study of the French Pyrenees is incomplete in two ways: there are few studies both on mountainous sites and in the central part of the chain.
La Pouretère is an infilled lake located at the limit of mountainous and sub-alpine vegetation belts (1720 m) in the Marcadau valley within the central Pyrenees.
The sedimentary sequence has been analysed for pollen and stomata and in some levels for macroremains. The results are presented in a simplified form as a relative frequency diagram with stomata and macroremains plotted on. Six 14C dating measurements were performed, which support the Lateglacial and Holocene chronology. The new data, in total accordance with previous piedmont regional studies, complete our knowledge of the vegetation history at medium and high elevation.
During the Bölling/Alleröd period, from 13 000±120 BP, trees were absent around the site except Juniperus and Salix shrubs. During the Younger Dryas, Artemisia and the other heliophilous taxa spread out noticeably. At the beginning of the Postglacial period, trees were still absent near the site. Betula was the first to reach La Pouretère surroundings during the Boreal, around 8900 BP. Pollen concentration, pollen clusters and macroremains, as found in another site in a nearby central Pyrenees valley (Estibère), demonstrate that Betula remained the principal altitudinal forest component until 5000 BP. Pinus arrived only around 8300 BP, as inferred by pollen concentration, stomata and macroremains data.
The abundance of Betula is recorded in the central part of the Pyrenees, from the Garonne Valley to the ‘Gave de Pau’ (Ulès, Estibère, La Pouretère). According to modern ecological studies in high latitudinal and altitudinal areas, it suggests that some climatic characteristics differ from those of the following Pinus extension phase: snowfalls were abundant, temperatures were colder at the beginning and at the end of the vegetation period, whereas the summer temperatures were higher. Those characteristics are in accordance with climatic hypothesis inferred from studies upon general orbital parameters changes and upon raininess on the Mediterranean area.
Despite first occurrences of Abies during the Lateglacial (LPAZ Po3) and the Boreal (LPAZ Po8), a regular presence is noticed near 6400 BP (LPAZ Po9). The beginning of the curve is dated 4860±150 BP. It is later than in the eastern part of the Pyrenees, but comparable with data from the Garonne Valley (Barbazan, central Pyrenees) and the Ossau valley (Estarrès, western Pyrenees). The early presence of pollen of Abies in the studied sites suggests the possible existence of refuges for this tree in the northwestern Pyrenean piedmont.
Afterwards, the vegetation changes are strongly influenced by human activities within the valley.
1 Introduction
Dans le versant nord des Pyrénées, d'ouest en est de la chaı̂ne, les étapes de la déglaciation et de la mise en place de la couverture végétale sont de mieux en mieux connues et datées. Néanmoins, excepté dans les Pyrénées orientales, les zones d'altitudes ont été peu étudiées à ce jour. La présente étude vise a combler en partie cette lacune.
Le site montagnard de La Pouretère (1720 m), étudié dans le cadre d'un programme de recherche consacré aux fluctuations de la limite supérieure de la forêt, permet de retracer les étapes de la dynamique de la végétation dans un haut bassin de la partie centrale du versant nord-pyrénéen : la vallée du Marcadau (Parc national des Pyrénées, Hautes-Pyrénées) (Fig. 1).
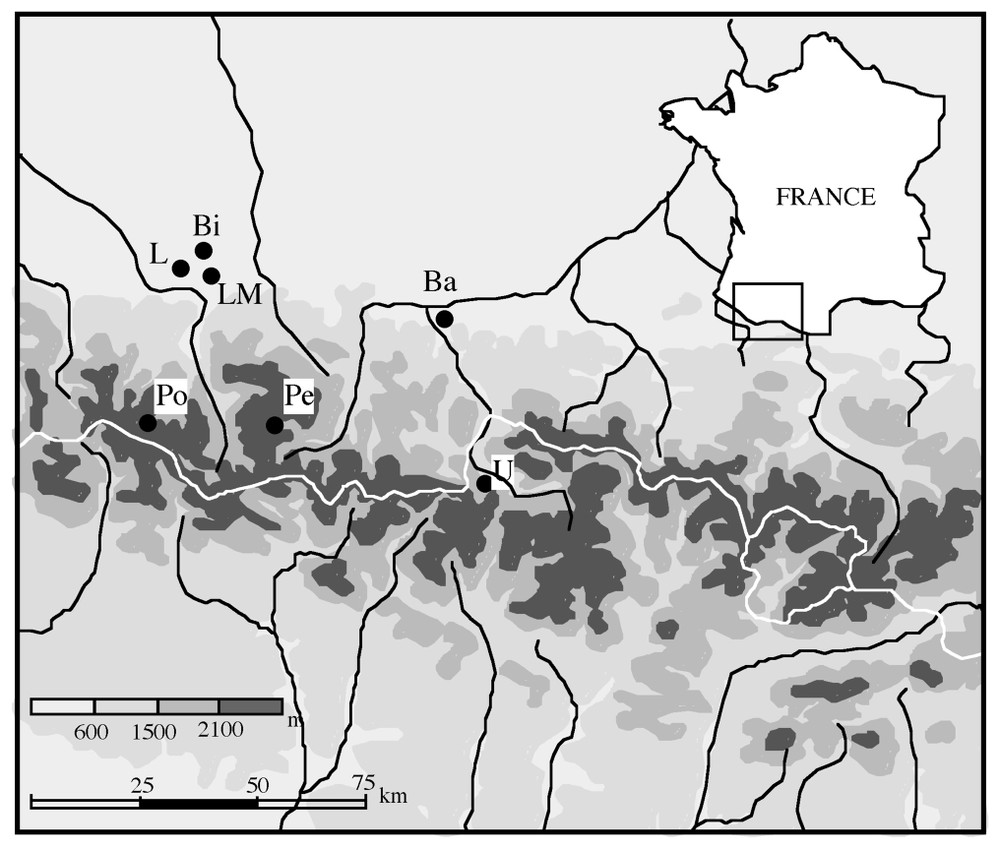
Localisation des sites mentionnés dans le texte. Po : Pouretère, Pe : Pé d'Estibère, Bi : Biscaye, LM : Le Monge, L : Lourdes, Ba : Barbazan, U : Ulès.
Dans les Pyrénées centrales, au niveau des étages montagnard et subalpin, la vallée du Marcadau est exceptionnelle de par le rôle joué à la fois par le pin sylvestre (Pinus sylvestris L.) et par son hybride avec le pin à crochets (P. uncinata Mill ex Mirb) : le pin de Bouget (P. bougetii). Le laquet de la Pouretère occupe une dépression au sein d'une sapinière (Abies alba Mill.), dans laquelle s'observent déjà P. uncinata et P. sylvestris. Le sondage a été effectué dans la partie sud de la tourbière, à environ 2 m du bord externe de la cuvette.
La localisation de notre site d'étude permet d'élucider la dynamique des espèces montagnardes actuelles, Pinus et Abies, dans cette partie ouest des Pyrénées centrales et depuis le retrait des glaciers. À la lumière des travaux palynologiques de David [1,2] dans les Alpes, et des recherches phytosociologiques de Grüber [3] dans les Pyrénées, nous avons été attentifs au rôle joué par le bouleau dans la mise en place de la couverture végétale.
2 Méthode
2.1 Analyse pollinique
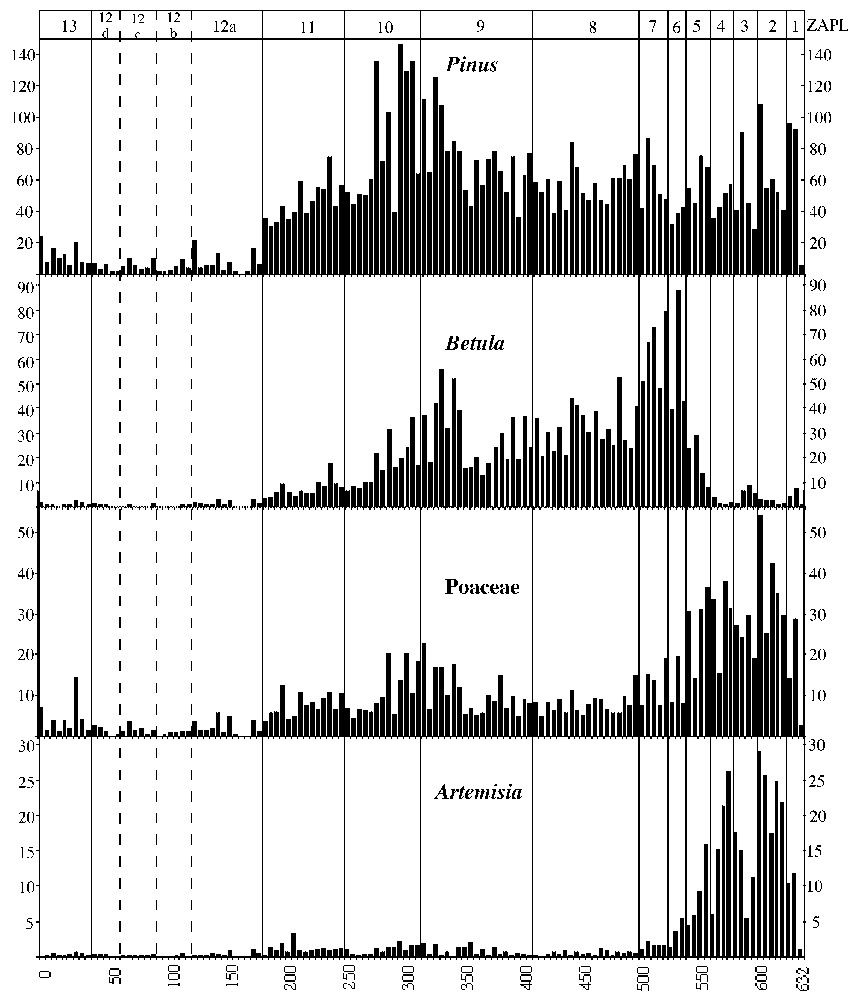
La séquence a été prélevée à l'aide d'un carottier russe [4]. Le traitement chimique des échantillons comprend classiquement des phases d'élimination de la matière minérale (HF), puis de la matière organique. Dans chaque niveau, au minimum 300 grains de pollen de végétaux terrestres ont été comptés et/ou 20 taxa déterminés [5]. Après analyse, chaque lame a été intégralement observée pour chercher les taxa rares. Le diagramme pollinique en fréquence relatives est établi grâce à un programme mis au point au laboratoire (Fig. 2). Les fréquences relatives sont calculées à partir d'une somme de base excluant les pollens des plantes aquatiques et les spores. Les fréquences inférieures à 1 % sont représentées par un point. Les assemblages polliniques caractérisant les zones d'assemblages polliniques locales (ZAPL) sont présentés sur la droite du diagramme (Fig. 2).

Diagramme pollinique simplifié en fréquences relatives. La Pouretère. Les macrorestes sont figurés par une lettre sur la courbe du taxon (B pour bois, A pour aiguille, G pour graine et St pour stomate), les amas polliniques par une étoile. Les courbes en grisé sont multipliées par 10. Les rectangles noirs indiquent l'épaisseur des niveaux datés. Lithologie : Ia–b : limono-sableux organique, II : limono-sableux, III : tourbe fibreuse.
2.2 Macrorestes
Certains macrorestes ont été prélevés pendant l'échantillonnage pollinique et déterminés. Leur présence est signalée près de la courbe pollinique du taxon.
Certains macrorestes végétaux ont été sélectionnés pour des datations AMS.
2.3 Stomates et amas polliniques
Les stomates de gymnospermes ont été recherchés, déterminés et comptés dans les lames d'analyse pollinique. Par ailleurs, nous avons recherché et compté les amas polliniques [6]. Notre analyse préalable du rapport actuel végétation/pluie pollinique nous a montré, en effet, l'existence d'une relation forte entre l'abondance des amas et la présence locale du taxon correspondant [7]. Stomates et amas sont notés près de la courbe pollinique du taxon concerné (Fig. 2).
2.4 Datations
Dans le texte, les âges 14C conventionnels sont exprimés par rapport au calendrier radiocarbone en années BP. Les mesures et les âges calibrés en années BP sont présentés dans le Tableau 1.
Datations 14C. L'âge calibré à 2σ a été obtenu par le logiciel CALIB rev. 4.3 de l'université de Washington (M. Stuiver et al., [11])
| Échantillon | Site | Prof. | Matériel daté | Âges 14C BP | ± | Âge calibré BP |
| UGRA 539 | Po | 34–35 | tourbe | 330 | ±110 | 545–71 |
| Gif 99234 | Po | 245 | bois | 3660 | ±80 | 4235–3725 |
| UGRA 538 | Po | 305–325 | gyttja | 4860 | ±150 | 5930–5089 |
| Gif 99235 | Po | 463 | bois | 7450 | ±70 | 8387–8058 |
| Gif 99096 | Po | 630–634 | argiles | 13000 | ±120 | 16119–14539 |
| UGRA 540 | PA | 630–640 | gyttja | 9120 | ±180 | 10742–9697 |
3 Résultats et discussion
Les données restituent les évènements régionaux majeurs de l'histoire de la végétation décrits lors des travaux antérieurs effectués en plaine [8–10].
Dès le Tardiglaciaire, on note cependant des particularités liées à l'altitude du site. La date de la base du sondage, 13 000±120 BP est peu informative au regard des événements polliniques, mais nous l'avons conservée comme base de notre échelle chronologique après confrontation avec les données précédemment publiées [8–10]. La chronozone Bölling–Alleröd (ZAPL Po1–Po2–Po3) est caractérisée par les taux peu élevés et les faibles concentrations polliniques de Pinus et de Betula (Fig. 3). Ils suggèrent que la place du pin dans le paysage local est faible, et l'on peut même considérer qu'ils reflètent sans doute en partie les peuplements de plaine. Toutefois, la présence locale du pin ne saurait être ignorée, du fait de la présence d'amas polliniques. Leur dispersion limitée renforce en effet cette hypothèse. La couverture végétale paraı̂t donc faiblement boisée, dominées par les Poacées et des taxons à affinité steppique, comme les Chénopodiacées et Artemisia. De plus, Juniperus, et quelques Salix, constituent sans doute une strate arbustive éparse. La ZAPL Po3 semble contemporaine de l'optimum Alleröd. Elle correspond, à la fois en fréquences relatives et en concentrations polliniques, à un net recul des Artemisia et, dans une moindre mesure, des Poacées.
Concentrations polliniques de taxa choisis exprimées en 103 grains cm−3 selon des profondeurs en cm.
Postérieurement, la LPAZ Po4 correspond au Dryas récent, enregistré plus nettement que dans les données de plaine [10]. On observe une nette reprise des herbacées et en particulier des steppiques, ainsi qu'un recul de Betula, de Pinus et de Juniperus. À l'altitude du site, la fin de cet épisode est marquée par le recul des herbacées et par un développement de Juniperus et de Salix antérieur au développement de Betula. L'amélioration climatique soulignée par ces changements de la végétation se traduit également par un enrichissement des sédiments en matière organique (transition entre unité Ia et Ib). Ses concentrations polliniques restant très faibles, il semble que Betula n'ait pas encore atteint l'altitude du site. La date du début de la courbe de Corylus dans un sondage proche de la même vallée, situé à l'étage montagnard (Le Paradis, 1619 m) [7] est de 9120±180 BP. Après calibration [11], cette date est similaire à celles obtenues pour le même événement, à basse altitude, dans la tourbière du Monge [10].
C'est au Boréal, vers 8900 BP (ZAPL Po6), que les taux, les amas et les concentrations polliniques de Betula permettent d'envisager la présence locale de cet arbre. Ce résultat confirme ceux obtenus dans une autre vallée des Pyrénées centrales, dont les caractéristiques climatiques sont proches de celles de la vallée du Marcadau : le vallon d'Estibère [12]. Dans cette haute vallée, des macrorestes de bouleaux trouvés à 2100 m d'altitude (Pé d'Estibère) dès 9600 BP attestent la présence locale du taxon, qui présente également des taux élevés. Le climat océanique d'abri [13] commun aux deux vallées peut expliquer cette convergence. Le bouleau joue un rôle également prépondérant dans la haute vallée de la Garonne (Ulès, 1650 m, val d'Aran, versant nord pyrénéen espagnol) [14,15], qui présente des caractéristiques climatiques similaires [13]. Ce rôle prépondérant du bouleau en altitude, plusieurs fois souligné dans le massif alpin [2,16–18], semble caractéristique des hautes vallées de la partie centrale des Pyrénées. Ce n'est que vers 8300 BP que Pinus se rapproche de l'altitude de notre site. Sa présence est attestée par une augmentation significative des concentrations polliniques, des concentration absolues de stomates de Gymnospermes et d'amas polliniques de Pinus (ZAPL Po7).
La connaissance des espèces impliquées permettrait de préciser les caractéristiques climatiques locales. Les stomates de pin comptés ne peuvent être déterminés jusqu'à l'espèce, et seules les études régionales présentant des analyses de macrorestes végétaux ou de charbons peuvent situer les espèces en place. D'après ces données, les deux espèces P. uncinata et P. sylvestris sont représentées dans les Pyrénées depuis le Tardiglaciaire. À Belvis (960 m d'altitude, Aude, plateau de Sault, Pyrénées orientales), des charbons de bois de ces espèces ont été datés à 12 270 BP [19]. À la Balma Margineda (960 m d'altitude, Andorre, Pyrénées méditerranéennes), on observe également des charbons des deux espèces, et ce dès 11 000 BP [20]. Il en est de même pour les espèces de bouleau. Au Tardiglaciaire, la détermination des macrorestes et des charbons s'arrête au genre Betula. Ce n'est qu'au début du Postglaciaire que les macrorestes ont pu être déterminés, jusqu'aux types B. pubescens ou B. pendula. Mais les deux espèces sont présentes simultanément (Ulès [14,15], Pé d'Estibère [12]). Les données régionales ne nous permettent donc pas de statuer sur la migration éventuellement différenciée des espèces de ces deux genres, qui nous permettrait de conclure quant aux conditions climatiques locales.
Seule l'opposition Betula–Pinus peut donc nous permettre d'interpréter nos résultats en termes climatiques. Si l'on se réfère aux travaux effectués en Scandinavie dans la zone actuelle de transition à Betula et la forêt à P. sylvestris [21], le caractère climatique déterminant la distribution des deux espèces semble être l'enneigement hivernal. Betula supporte les contraintes d'un enneigement précoce, lourd et long, tandis que ces conditions sont défavorables à Pinus sylvestris. D'après des travaux pyrénéens [22], ces conditions sont également défavorables à P. uncinata, qui est alors remplacé (dans la partie nord-ouest du val d'Aran, dans cet exemple) par une association d'arbres à feuillage caduc (B. pubescens, B. pendula et Sorbus aucuparia). On peut alors supposer que dans les vallées concernées (Marcadau, Estibère, val d'Aran), les conditions climatiques du début du postglaciaire, notamment l'enneigement hivernal, ont été favorables à Betula, qui actuellement présente la plus grande amplitude écologique. Les précipitations neigeuses hivernales importantes, couplées à des températures plus froides, au début et à la fin de la période de végétation, ont pu être défavorables à Pinus, malgré des températures estivales accrues par rapport au Tardiglaciaire. Cette hypothèse est renforcée par les travaux de Kutzbach et Guetter [23], qui concluent, au Préboréal, à une amplitude thermique maximale (radiation solaire en été >8%, radiation hiver <8%) et par les recherches menées en Méditerranée, qui concluent, pour la même période, à une augmentation des précipitations continentales liées au réchauffement des eaux de surface de la mer [24,25]. Ce n'est qu'après 8000 BP que, les conditions climatiques permettant aux peuplements de s'affermir, la distribution locale de la neige est modifiée. Pinus prend alors peu à peu une place prépondérante.
Les premières occurrences polliniques régulières d'Abies datent de 6400 BP (Po9), mais les premiers pollens sont observés dans des sédiments du Tardiglaciaire (Po3) et du Boréal (Po8). Le début de la courbe continue est daté à 4860±150 BP. Le remplacement de la pinède par la sapinière se fait assez progressivement, puis l'optimum de la sapinière au niveau du site survient entre 3660 et 2650 BP, ainsi que l'attestent les pourcentages, les macrorestes et les concentrations polliniques. Ce développement est récent au regard de la dynamique du taxon dans les Pyrénées orientales, où la courbe continue du taxon débute vers 8250 BP [26]. Mais il est comparable à celui observé à l'est, dans le piémont de la vallée de la Garonne (Barbazan, Pyrénées centrales) [27], et à l'ouest, dans la basse vallée d'Ossau (Pyrénées occidentales), à Estarrès (356 m d'altitude ; 4860±70 BP, GIF 7071) [28]. La présence de pollen d'Abies au Tardiglaciaire et au Boréal dans le Marcadau ou pendant le Pré-Boréal dans la vallée de la Garonne (Barbazan) et, enfin, plus à l'ouest dans les Pyrénées occidentales (Estarrès) nous conduisent à envisager sur le piémont nord pyrénéen l'existence de refuges würmiens multiples, comparables à celui décrit dans la partie orientale de la chaı̂ne (charbons de bois [19]), ce qui s'oppose à l'hypothèse d'une migration est–ouest de ce taxon.
Conformément aux données régionales, la date 3660±80 BP (ZAPL Po11) correspond au début de la courbe continue de Fagus, qui marque le développement des hêtraies aux altitudes inférieures.
À partir de 2650 BP, les influx polliniques de tous les taxons arboréens chutent de façon drastique. Cette date correspond à l'augmentation de Plantago type lanceolata, de Cerealia, des Ericacées et des Cyperacées. L'ouverture du couvert forestier est confirmée par un meilleur enregistrement des pollen de la chênaie collinéenne (Quercus ZAPL Po12a). Ces événements marquent le début d'une phase de développement des activités humaines en altitude. Ce phénomène est observé dans l'ensemble des Pyrénées [29] à la fin de l'âge du bronze et au début de l'âge du fer. Des monuments mégalithiques datant probablement de cette période ont été retrouvés sur les chemins d'estives actuels dans la vallée du Marcadau [30]. Une approche couplant données polliniques et données archéologiques et/ou historiques, sur le modèle des travaux de Galop [29], nous a permis de montrer l'importance du facteur anthropique dans la dynamique de la végétation de cette période à nos jours [7].
4 Conclusion
Cette étude décrit les étapes de la mise en place de la végétation au niveau de l'étage montagnard actuel d'une vallée des Pyrénées centrales. Elle complète l'étude paléoécologique de la chaı̂ne pyrénéenne, dans sa zone centrale et à moyenne altitude. Elle illustre également l'intérêt des analyses « multi-proxy » dans la reconstitution de la dynamique de la végétation et du climat à l'échelle locale.
Elle souligne enfin la complémentarité d'études de sites localisés dans les différents étages de végétation d'une même région pour une reconstitution précise de l'histoire de la mise en place de ces étages. Alors que les sondages du bassin de Lourdes [8–10] permettent de retracer précisément l'histoire de l'installation des chênaies et des hêtraies, notre étude permet une meilleure caractérisation de la dynamique des sapinières et des pinèdes.
Ici, comme dans le reste des Pyrénées, Abies joue bien un rôle majeur à l'étage montagnard et ce dès 5000 BP. Betula est prépondérant pendant la première partie du postglaciaire, à l'étage montagnard, et peut-être dans l'étage subalpin [7,12], dans les Pyrénées centrales.
Cette observation ouvre de nouvelles perspectives de recherche en ce qui concerne la caractérisation climatique des épisodes tardiglaciaires et holocènes à moyenne et haute altitude sur le versant nord de la chaı̂ne, et elle suggère des convergences avec le domaine alpin.