

Abridged English version
Today, the theory of the chemostat has been widely studied in the literature (see, for instance, the textbook [14]). The classical model describing the competition of several species
| (1) |
In this work, we propose to replace in (1) the classical functional responses
| (2) |
We introduce the concept of steady-state characteristic
Our main result is the following: assume that the function
This persistence result generalizes previous results where the
1 Introduction
Le chémostat a été introduit il y a plus d'un demi-siècle dans l'étude de la croissance des micro-organismes, et le modèle mathématique ci-dessous :
| (3) |
Un modèle analogue de compétition entre plusieurs organismes dans le même dispositif a été ensuite étudié, soit :
| (4) |
D'autres travaux ont considéré le modèle plus général :
| (5) |
| (6) |
| (7) |
En 1976, Armstrong et McGehee [3,12] ont fait remarquer que la coexistence ne se fait pas nécessairement à l'équilibre et que des oscillations forcées (modèle du chémostat (4) où
En 1989, Arditi et Ginzburg [2], dans le contexte de l'étude de la relation « proie–prédateur » ont proposé de remplacer la réponse fonctionnelle
2 Le modèle « un consommateur – une ressource »
La quantité de ressource disponible à l'instant t est désignée par
| (8) |
Hypothèses H1
- 1.
La fonction
- 2.
- 3.
La fonction
- 4.
Pour tout
Proposition 1
Sous l' Hypothèse H1 , les propriétés suivantes sont vérifiées.
- (i)
Pour tout
- (ii)
La fonction
Preuve
Soit
Soit
Définition 1
Nous appelons caractéristique à l'équilibre la fonction
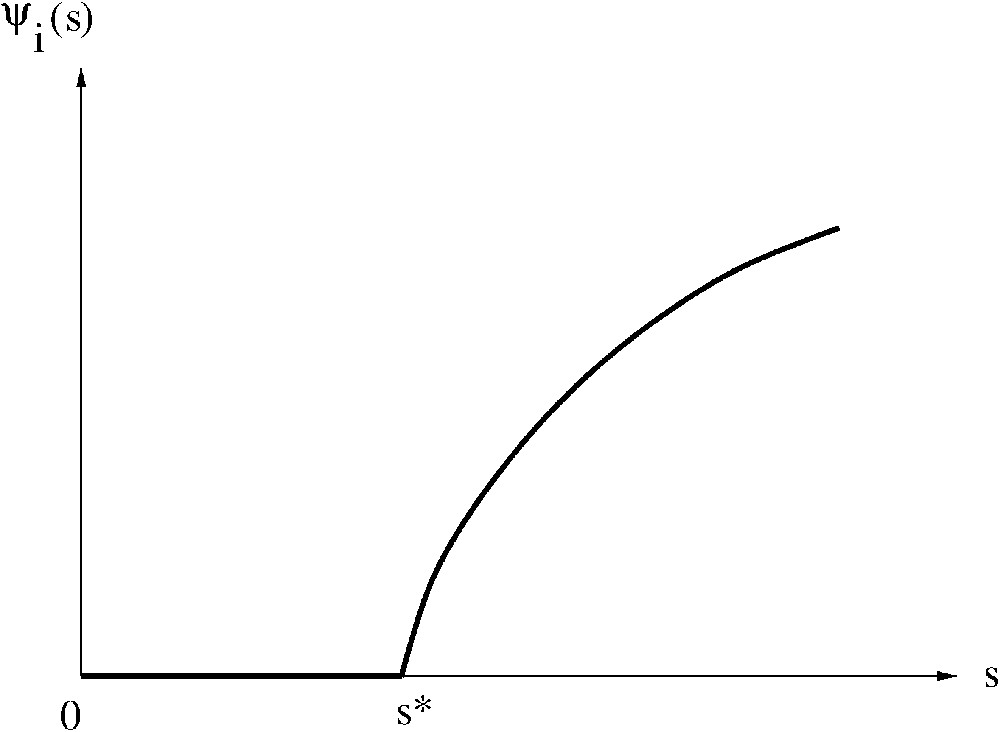
Exemple de caractéristique à l'équilibre.
Remarque 1
La caractéristique à l'équilibre que nous venons de définir est une partie de ce qu'on appelle usuellement l' « isocline du prédateur » dans la relation « proie–prédateur » du modèle (9) ci-dessous
| (9) |
Remarque 2
En théorie mathématique des systèmes, la quantité s s'appellerait « entrée » du système décrit par l'Éq. (8). Dans un article récent [1], l'importance des systèmes pour lesquels à chaque entrée on peut associer un unique équilibre a été démontrée. Cette correspondance a été appelée input-to-state characteristic. Bien que notre fonction
Exemple 1
| (10) |
| (11) |
Exemple 2
| (12) |
| (13) |
Exemple 3
| (14) |
- – Si
- – Si
- – Si
Théorème 1
Considérons le système(9), dont nous supposons que la réponse fonctionnelle
| (15) |
Preuve
Elle est extrêmement simple. Nous devons d'abord montrer que
| (16) |
| (17) |
Le Théorème 1 ne présente aucune originalité et n'est là que pour rappeler ce que nous généralisons. Il est d'autre part connu que, sans hypothèse supplémentaire, cet équilibre n'a aucune raison d'être globalement stable.
3 Le modèle de compétition « n consommateurs – une ressource »
Nous considérons dans ce paragraphe le modèle :
| (18) |
Théorème 2
Considérons le système(18)dont nous supposons que tous les
| (19) |
Preuve
Nous devons montrer que
| (20) |
| (21) |
| (22) |
Le Théorème 1 est clairement le cas particulier du Théorème 2 dans le cas où
Il est clair que pour tous les i tels que
4 Discussion
4.1 Réalisme biologique des Hypothèses H1
Pour de nombreuses d'espèces, il est réaliste d'imaginer que le taux de croissance d'une population, toutes choses égales par ailleurs, diminue quand la taille de la population augmente. C'est un phénomène de compétition « intraspécifique » pour l'accès à la ressource. Toutefois, il peut exister, pour certaines espèces, une forme de mutualisme qui contredirait cette hypothèse. Il existe un cas, celui de la croissance d'une espèce dans un milieu liquide, où la diffusion du substrat peut devenir un facteur limitant si les organismes ont tendance à constituer des colonies plus ou moins sphériques. Dans ce cas, seule la zone en périphérie a accès à la ressource et alors la considération du rapport volume/surface conduit à introduire une réduction du taux de croissance tel que dans l'Exemple 2. Le cas classique de la « loi d'action de masse » pure où la fonction
Le cas de la « ratio dépendance », dès lors qu'on introduit un petit paramètre α comme dans l'Exemple 3, apparaît comme un cas particulier de notre modèle. L'introduction de ce petit paramètre ne change rien d'essentiel à la philosophie développée par les partisans de la ratio-dépendance. Une partie des arguments en faveur de la ratio dépendance peut donc être reprise en faveur de notre modèle.
Nous avons supposé un taux de disparition d constant. Poursuivant la même logique il serait judicieux de le rendre lui aussi dépendant de la densité et d'écrire
| (23) |
Il est possible de le faire sans changer les conclusions de cette note, mais la présentation aurait été un petit peu plus compliquée. Toujours par souci de simplicité, nous avons supposé que la réponse fonctionnelle est égale au taux de croissance. On sait que, si ces derniers sont proportionnels, on peut, par simple changement d'unités, éliminer la constante de proportionnalité. Mais, même dans le cas où il n'y a pas proportionnalité, on peut sans peine étendre les résultats de la note.
Dans la Réf. [10] sont développés, dans le contexte de l'écologie microbienne, des arguments en faveur de réponses fonctionnelles du type de notre Exemple 3.
4.2 Critique du modèle de compétition proposé
Le modèle défini par les Éqs. (18) n'est pas la conséquence logique du modèle défini par les Éqs. (9), comme les Éqs. (4) le sont de (3). En effet, dans ce dernier cas, il était supposé que tous les individus d'une espèce ont à tout moment le même accès à la ressource, et il y a une certaine logique à supposer qu'il en est de même pour un mélange d'individus d'espèces différentes. Dans notre cas, dans la mesure où nous supposons une certaine forme de compétition entre les individus d'une même espèce pour accéder à la ressource, il serait peu logique de ne pas supposer qu'elle existe aussi entre les individus des espèces différentes. Cela se traduirait mathématiquement par l'introduction de fonctions
| (24) |
Toutefois, nous pouvons justifier notre modèle avec l'argument suivant. Dans tous les cas où les individus de la même espèce ont tendance à rester ensemble (que ce soit la conséquence d'un comportement grégaire ou la conséquence de la division cellulaire dans le cas de micro-organismes), les effets de la compétition intraspécifique seront plus importants que ceux de la compétition interspécifique. Mathématiquement, grâce aux théorèmes généraux de stabilité structurelle [5], nous pouvons affirmer que, si les dérivées partielles de
4.3 Critique de l'étude à l'équilibre
Les remarques de Armstrong et McGhee restent valables, mais il faudrait continuer à examiner le modèle (18), non plus du seul point de vue des équilibres, mais aussi de celui de la persistance « tout court ». Dans la note [9], il est montré que, dans le cas du chémostat (
Dans [6], une condition suffisante de persistance est démontrée pour le modèle particulier où la fonction
Annexe A
Soit un système de n équations différentielles
| (A.1) |
| (A.2) |
Définition 2
La matrice jacobienne du système (A.1) à l'équilibre est la matrice
| (A.3) |
Soit λ une valeur propre de la matrice J, c'est-à-dire un nombre réel ou complexe tel qu'il existe un vecteur V non nul vérifiant
| (A.4) |
| (A.5) |
| (A.6) |
| (A.7) |
- – soit
- – soit
(A.8)
| (A.9) |
| (A.10) |
| (A.11) |
| (A.12) |
Vous devez vous connecter pour continuer.
S'authentifier