


 CC-BY 4.0
CC-BY 4.0
1. Introduction
La promotion des mélanges d’essences forestières est aujourd’hui reconnue comme une stratégie pertinente d’adaptation des forêts au changement climatique (Jourdan et al., 2021; Legay et al., 2008). De nombreuses études ont mis en évidence une augmentation de la productivité, et, dans certains cas, une amélioration de la résilience et de la résistance des peuplements forestiers avec la diversité des espèces (Jactel et al., 2017; Liang et al., 2016). Toutefois, ces effets positifs ne sont pas systématiques (Grossiord, 2020), et la communauté scientifique s’accorde à souligner que l’identité des espèces — autrement dit, la composition spécifique du mélange — joue un rôle déterminant (Pardos et al., 2021). Il est d’ailleurs important de souligner que cet « effet mélange » peut parfois s’accompagner d’une forte asymétrie des réponses entre les espèces au sein du mélange (Toïgo et al., 2015). Bien qu’il ne soit pas encore possible de définir un mélange optimal pour chaque contexte écologique, un consensus émerge sur le fait que, de manière générale, le recours aux peuplements mélangés doit être dorénavant plus important.
Favoriser les mélanges d’essences implique à la fois de maintenir les espèces déjà présentes ou de favoriser des essences minoritaires, et d’introduire de nouvelles espèces dans les peuplements nettement dominés par une seule essence. Cette double approche (gérer l’existant, créer des assemblages d’espèces) est essentielle, d’autant plus que, selon l’IGN, environ 50 % des forêts françaises sont constituées de peuplements monospécifiques (IGN, 2019). La distinction entre le maintien et la création de mélanges s’avère ainsi fondamentale en gestion forestière. Dans le premier cas, il s’agit de garantir les conditions nécessaires au renouvellement naturel et à la croissance des espèces en place : cela implique de préserver une densité suffisante de semenciers, de répondre aux exigences écologiques de la régénération (cf. niche de régénération ; Grubb, 1977), et de gérer les dynamiques de compétition entre les espèces au cours des différentes phases de développement (Schwinning and Weiner, 1998). Dans le second cas, la création de mélanges repose principalement sur la phase de renouvellement, souvent par plantation. Cette méthode permet un meilleur contrôle de la composition et de l’abondance relative des espèces, notamment dans le cadre de plantations mélangées en plein.
Depuis quelques années, les stratégies de diversification forestière se sont complexifiées avec le développement des plantations d’enrichissement (ex. Billard et al., 2024) et la promotion de la sylviculture mélangée à couvert continu (Mason et al., 2022). Dans le premier cas, l’objectif consiste à introduire de nouvelles essences par plantation, à faible densité, au sein d’une régénération naturelle ou dans un peuplement déjà établi, qu’il soit géré en futaie irrégulière ou en futaie régulière. Cette pratique peut, par exemple, être mise en œuvre dans des trouées en futaie irrégulière, au sein d’une régénération installée en futaie régulière, ou encore sous couvert forestier, quel que soit le mode de traitement. Dans le deuxième cas, l’accent est mis sur le maintien de la diversité en espèces ou l’amélioration de cette diversité dans des systèmes sylvicoles à renouvellement continu (futaie irrégulière), visant ainsi des structures verticalement plus stratifiées et plus résilientes aux perturbations. La mise en œuvre de ces approches toujours plus complexes soulève plusieurs défis : écologiques, en raison de l’incertitude sur l’adaptation des espèces introduites aux conditions locales et sur leurs interactions ; économiques, du fait des coûts associés aux travaux, à l’entretien et au suivi.
Quelle que soit la méthode retenue, l’objectif est bien de garantir à moyen terme l’acquisition d’un mélange (chaque espèce doit être suffisamment abondante dans le peuplement) et de maintenir ce mélange sur une durée significative de la vie du peuplement. Ces notions d’acquisition et de maintien des mélanges reposent à la fois sur des considérations écologiques et sylvicoles et posent la question de la compatibilité des espèces, terme davantage utilisé en gestion que dans la sphère de la recherche académique. Dans le domaine forestier, cette compatibilité, telle que proposée par Combaud (2024), élargit celle, plus théorique, de coexistence en écologie (Chesson, 2018), en y intégrant les objectifs de gestion forestière. Autrement dit, un mélange est dit « compatible » si sa pérennité peut être raisonnablement assurée par la gestion sur une durée significative du cycle de vie du peuplement — par exemple, une révolution en futaie régulière ou plusieurs rotations en futaie irrégulière.
Cette confrontation entre écologie et gestion met en lumière deux enjeux principaux : (i) l’intérêt d’intégrer de manière explicite la sylviculture dans la théorie écologique, qui aborde depuis longtemps la question de la coexistence des espèces (Cordonnier, Kunstler et al., 2018), mais sans prendre en compte les multiples contraintes et leviers de gestion ; (ii) le besoin de développer des itinéraires sylvicoles adaptés aux peuplements mélangés en y intégrant davantage les connaissances écologiques, itinéraires par ailleurs peu développés dans le contexte forestier français.
Dans cet article nous illustrons le premier enjeu au travers de l’exemple de la sylviculture mélangée à couvert continu et de l’utilisation d’un modèle assez simple de dynamique forestière, essentiellement utilisé pour traiter des questions d’écologie et de biogéographie. En particulier, nous montrons comment différents mécanismes de coexistence peuvent être traduits en actions sylvicoles et testons leur efficacité en termes de maintien du mélange. Dans un souci de simplicité, nous concentrons notre analyse sur deux mélanges d’essences connus pour être difficiles à conduire en gestion forestière : le mélange composé de chêne sessile (Quercus petraea (Matt.) Liebl.) et de hêtre commun (Fagus sylvatica L.) ainsi que le mélange de sapin pectiné (Abies alba Mill.) et de pin sylvestre (Pinus sylvestris L.). Notre cas d’étude n’embrasse bien entendu pas toute la complexité de la gestion et du comportement des espèces en mélange. Ils se concentrent davantage sur la démographie des espèces, délaissant d’autres sujets d’importance, comme la compétition avec la végétation concurrente (Balandier et al., 2006) ou la prise en compte d’autres facteurs biotiques comme l’herbivorie (Bernard et al., 2017).
2. Évaluer la compatibilité des espèces en gestion
Évaluer la compatibilité des espèces dans un cadre de gestion s’avère chose délicate. Elle repose aujourd’hui essentiellement sur les caractéristiques fonctionnelles et de croissance intrinsèques des espèces, notamment :
- la tolérance à la compétition intraspécifique et interspécifique (Kunstler, Falster et al., 2016), souvent évaluée au travers de la tolérance à l’ombre (Niinemets and Valladares, 2006). Plus les espèces auront des tolérances différentes, plus le mélange sera jugé difficile à maintenir (Bauhus et al., 2017).
- La capacité à faire de l’ombrage : plus une espèce aura un fort pouvoir ombrageant (houppier large avec une forte densité foliaire), plus il sera difficile de la mener en mélange (Combaud, 2024). Cette capacité est souvent corrélée positivement à la tolérance à l’ombre (Niinemets, 2010).
- La capacité à croître rapidement en phase juvénile : une forte différence de croissance initiale peut rendre difficile le maintien des espèces dans le cadre d’un mélange dit pied à pied (Combaud, 2024).
Ces caractéristiques sont par exemple prises en compte dans les documents de gestion pour définir le design de mise en place des mélanges, que ce soit dans le cadre de plantations en plein ou des plantations en enrichissement (Sardin et al., 2026). Elles sont également utilisées lors des éclaircies pour tenir compte des sensibilités différentielles des espèces à la compétition (Bauhus et al., 2017; Pretzsch et al., 2021).
Par exemple, pour nos mélanges retenus, le hêtre commun représente une espèce fortement compétitive pour le chêne sessile, car elle est beaucoup plus tolérante à l’ombre (indice de Niinemetz et Valladares de 4,56 contre 2,73) tout en ayant une croissance similaire, voire supérieure en phase juvénile dans les peuplements où les deux espèces sont en présence (ex. Ligot et al., 2013). Par ailleurs, le hêtre commun présente un pouvoir ombrageant supérieur à celui du chêne sessile, formant des peuplements à plus forte surface foliaire (Leuschner and Meier, 2018). La conduite du mélange de chêne sessile et de hêtre commun est donc considérée comme particulièrement difficile, quel que soit le mode de traitement (ex. von Lüpke, 1998). Dans le cas du mélange de sapin pectiné et de pin sylvestre, c’est le sapin pectiné qui est considéré comme plus compétitif, étant plus tolérant à l’ombre que le pin sylvestre (indice de Niinemetz et Valladares de 4,6 contre 1,67). Le sapin pectiné a également un pouvoir ombrageant supérieur à celui du pin sylvestre (Leuschner and Meier, 2018). En revanche, dans ce mélange et en pleine lumière, le pin sylvestre possède souvent une croissance initiale en phase juvénile supérieure à celle du sapin pectiné, ce qui le rend par exemple particulièrement intéressant comme espèce d’enrichissement.
3. Des théories de la coexistence en écologie
Comme nous l’avons souligné en introduction, la notion de coexistence en écologie est plus restrictive que celle de compatibilité, puisqu’elle suppose le maintien des espèces à l’échelle d’un grand nombre de générations, voire une coexistence stable.
Un des résultats forts en écologie est que la coexistence des espèces ne peut être obtenue que si la compétition entre espèces (dite interspécifique) est plus faible que la compétition au sein de chaque espèce (dite intraspécifique). Cela a été redémontré par exemple par (Chesson, 2000b; Chesson, 2018), qui a développé un cadre mathématique pour caractériser les mécanismes de coexistence au sein de communautés en environnement stable et en environnement variable, dans le temps ou dans l’espace. Tout mécanisme écologique permettant d’obtenir ou facilitant l’obtention de cette condition (en espérance lorsque les conditions sont variables) constitue ainsi un mécanisme de coexistence. Chesson (2018) distingue alors les mécanismes stabilisateurs, qui conduisent les espèces à occuper des niches différentes et stabilisent fortement la coexistence, et les mécanismes égalisateurs, qui réduisent les différences d’aptitude (fitness) entre espèces et donc renforcent l’effet des mécanismes stabilisateurs. Malgré un fort intérêt de la communauté scientifique pour cette théorie, elle demeure encore aujourd’hui assez déconnectée de l’écologie empirique et n’a fait l’objet que d’un nombre limité de tests in situ (Ellner et al., 2019). Un mécanisme stabilisateur simple de la coexistence est, par exemple, la fréquence-dépendance négative qui réduit l’aptitude d’une espèce lorsqu’elle devient abondante (Chesson, 2000b).
D’autres théories et mécanismes de coexistence impliquant davantage les caractéristiques des espèces ont été mis en avant. En sus de la théorie de Chesson et de la fréquence-dépendance négative déjà citées, nous nous concentrerons ici sur deux autres théories qui semblent bien adaptées au cas des écosystèmes forestiers (Cordonnier, 2019).
La théorie de la niche de succession repose sur le compromis entre croissance en pleine lumière et survie à l’ombre, ainsi que sur un taux de perturbation minimal dans un fonctionnement spatial en mosaïque (Gravel et al., 2010; Pacala and Rees, 1998). L’espèce intolérante à la compétition et à fort potentiel de croissance persiste en profitant davantage des pics de ressources créées par les perturbations. En revanche, l’espèce tolérante à la compétition survit mieux en situation de faible ressource. Le compromis entre croissance en pleine lumière et survie à l’ombre a été mis en évidence aussi bien en forêt tropicale (Hubbell, 2001) qu’en forêt tempérée (Seiwa, 2007). À l’échelle locale du peuplement, ce compromis, lorsqu’il existe, peut également expliquer la coexistence de deux espèces, mais uniquement dans des contextes caractérisés par des fréquences de perturbations élevées (Gravel et al., 2010). Dans ce cas, le mécanisme de coexistence à l’œuvre, qui opère dans un environnement variable, est la non-linéarité relative (Chesson, 2018; Gravel et al., 2010), laquelle traduit une différence dans la manière dont les espèces répondent aux variations temporelles de la compétition, ici pilotées par les perturbations puis la fermeture progressive du couvert.
La théorie de la stratification (ou théorie du partitionnement foliaire lorsque les espèces occupent la même strate à maturité) repose sur quatre traits : la hauteur à maturité sexuelle, la croissance maximale, la survie au stade adulte et la fécondité (T. Kohyama and Takada, 2009; T. S. Kohyama and Takada, 2012). Des compromis entre certains de ces traits conduisent à la coexistence de plusieurs espèces. Dans cette théorie, le compromis fécondité-survie ou recrutement-survie offre une alternative à ceux basés sur la tolérance à l’ombre (cf. niche de succession ci-dessus). Tout comme dans le cas de la niche de succession, le nombre d’espèces pouvant coexister à l’échelle du peuplement (patch) selon ce compromis reste faible. Il s’agit d’une des rares théories qui s’affranchisse des perturbations et donc puisse offrir une explication à la coexistence des espèces dans des situations à faible régime d’ouverture de la canopée. À notre connaissance, aucun lien explicite n’a encore été établi entre cette théorie et les mécanismes de coexistence définis dans la théorie de Chesson.
4. La sylviculture comme facteur de coexistence
Quels peuvent être les effets de la gestion sur la coexistence des espèces à l’échelle du peuplement ? Quelles sont les actions les plus efficaces ? Quels sont les mélanges les plus faciles à gérer ? Dans le cadre de la diversification pour l’adaptation, ces questions deviennent essentielles. En corollaire, on peut légitimement se demander en quoi les théories existantes peuvent nous aider sur ces questions. Dans cette section, nous présentons un exercice de simulation où un lien explicite est réalisé entre des paramètres de sylviculture et les théories et mécanismes de la coexistence présentés précédemment.
4.1. Le modèle MATREEX
Nous utilisons le modèle de projection intégrale MATREEX qui permet de simuler la dynamique de communautés d’arbres forestiers sur le long terme (Jaunatre et al., 2026; Kunstler, Guyennon et al., 2021). Ce modèle a été utilisé pour aborder des questions d’écologie fondamentale, par exemple les effets du climat et de la compétition sur les performances démographiques des espèces (Baranger, 2025; Guyennon et al., 2023) ou l’influence de la diversité fonctionnelle sur la résilience des peuplements (Barrere et al., 2024). Ce modèle comprend des modules de sylviculture, dont un est adapté à la gestion en futaie irrégulière qui favorise la diversité des tailles des arbres au sein du peuplement (Jaunatre et al., 2026, Figure 1). Ce module comprend un nombre limité de paramètres clés en sylviculture irrégulière : diamètre d’exploitabilité dha (seuil à partir duquel on privilégie la récolte des arbres plutôt que l’amélioration par l’éclaircie), surface terrière cible après coupe (BAtarget), taux maximum de coupe (Pmax), taux maximum de récolte (hmax), seuil de diamètre minimal d’éclaircie (dth). Le paramètre k, déterminé par optimisation numérique, décrit quant à lui l’évolution du taux de prélèvement, décroissant, entre le diamètre d’exploitabilité et le diamètre minimal d’éclaircie. Un paramètre (a) a également été introduit pour gérer une fréquence-dépendance des coupes : le prélèvement cible ainsi davantage l’espèce la plus abondante (quand a = 1, il n’y pas de fréquence-dépendance). La préférence à l’espèce la plus abondante est définie par (p : proportion en surface terrière de l’espèce la plus abondante) :
| \begin {equation*} f(p,a)= \frac {p^a}{p^a+(1-p)^a} \end {equation*} |
Exemple de courbe déterminant le taux de coupe (harvesting) h selon le diamètre des arbres dbh. dth = 17,5 cm, dha = 57,5 cm, hmax = 0,8. Le paramètre de décroissance k de la courbe s’adapte à la structure du peuplement (ici k = 2).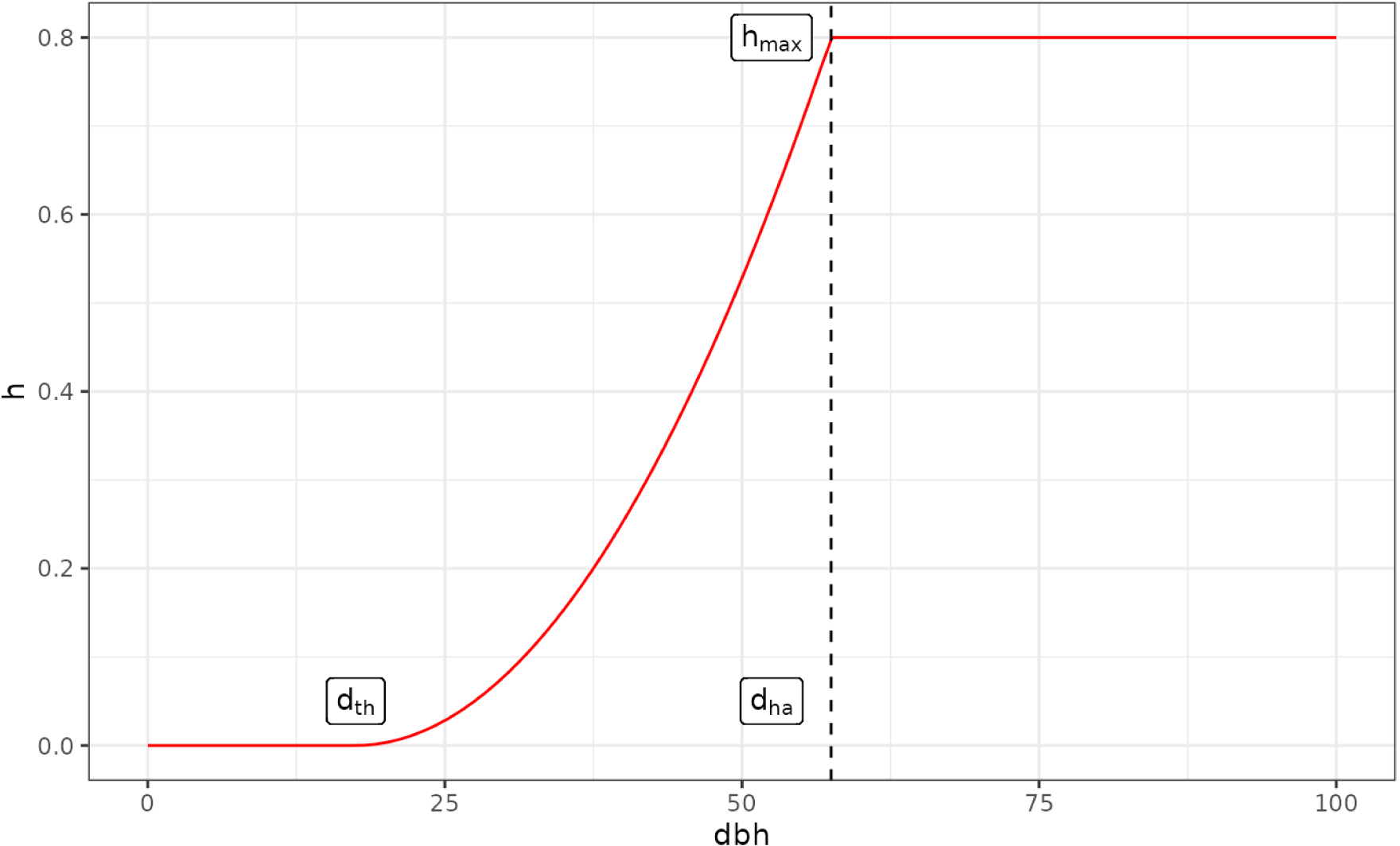
Le modèle MATREEX est disponible sur la plateforme GITHUB (GitHub—gowachin/matreex: LESSEM internal package for simulation of forest dynamic depending on climatic variables) sous forme de package R (matreex). Le modèle de coupe dit « uneven-aged » est décrit de manière détaillée dans une vignette à l’adresse suivante https://gowachin.github.io/matreex/articles/Harvesting.html.
4.2. Absence de coupes et sylviculture de référence
Le modèle permet de simuler la dynamique de nos deux mélanges en l’absence de coupes (il existe tout de même un taux de prélèvement constant minimal de 0,5 % par an utilisé pour éviter de surestimer l’espérance de vie des arbres, cf. Kunstler, Guyennon et al., 2021). Le modèle MATREEX intègre les effets du climat sur les processus démographiques (mortalité, croissance). Pour chaque mélange, nous retenons le climat optimal de l’espèce la moins compétitive (chêne sessile pour le mélange de hêtre commun et de chêne sessile, et pin sylvestre pour le mélange de pin sylvestre et de sapin pectiné). Nous partons d’une situation de très faible densité des deux espèces (surface terrière de 1 m2). Il est important de souligner que la disparition complète d’une espèce (exclusion compétitive) n’est pas possible dans le modèle MATREEX (la proportion de l’espèce tend vers 0) et que l’on s’intéresse ici plutôt aux taux de mélange, avec l’idée que, du point de vue de la gestion, l’objectif consiste à maintenir une proportion en surface terrière non négligeable de l’espèce la moins abondante, c’est-à-dire supérieure ou égale à 0,2.
Les simulations indiquent la quasi-disparition de l’espèce la moins compétitive, en l’absence de perturbations ou de gestion (Figure 2A,B). Celui du pin sylvestre est particulièrement rapide. En appliquant une sylviculture de référence (cf. Tableau 1), établie sur la base des travaux de Lafond et al. (2017), le patron reste inchangé pour le mélange de pin sylvestre et de sapin pectiné alors qu’on observe un très léger effet positif pour le mélange de chêne sessile et de hêtre commun, le pourcentage de chêne sessile en surface terrière restant bien inférieure à 20 % (7,6 %).
Dynamique simulée de la surface terrière BA (m2⋅ha−1) des espèces par le modèle MATREEX pour les deux mélanges étudiés : (A,B) sans coupes (sur 1500 ans) et (C,D) avec coupes en futaie irrégulière (sur 750 ans). On remarque un très léger effet positif des coupes sur la présence du chêne sessile en mélange avec le hêtre commun (proportion très inférieure à 0,2). En revanche, le pin sylvestre ne se maintient guère en mélange avec le sapin pectiné (tend vers 0).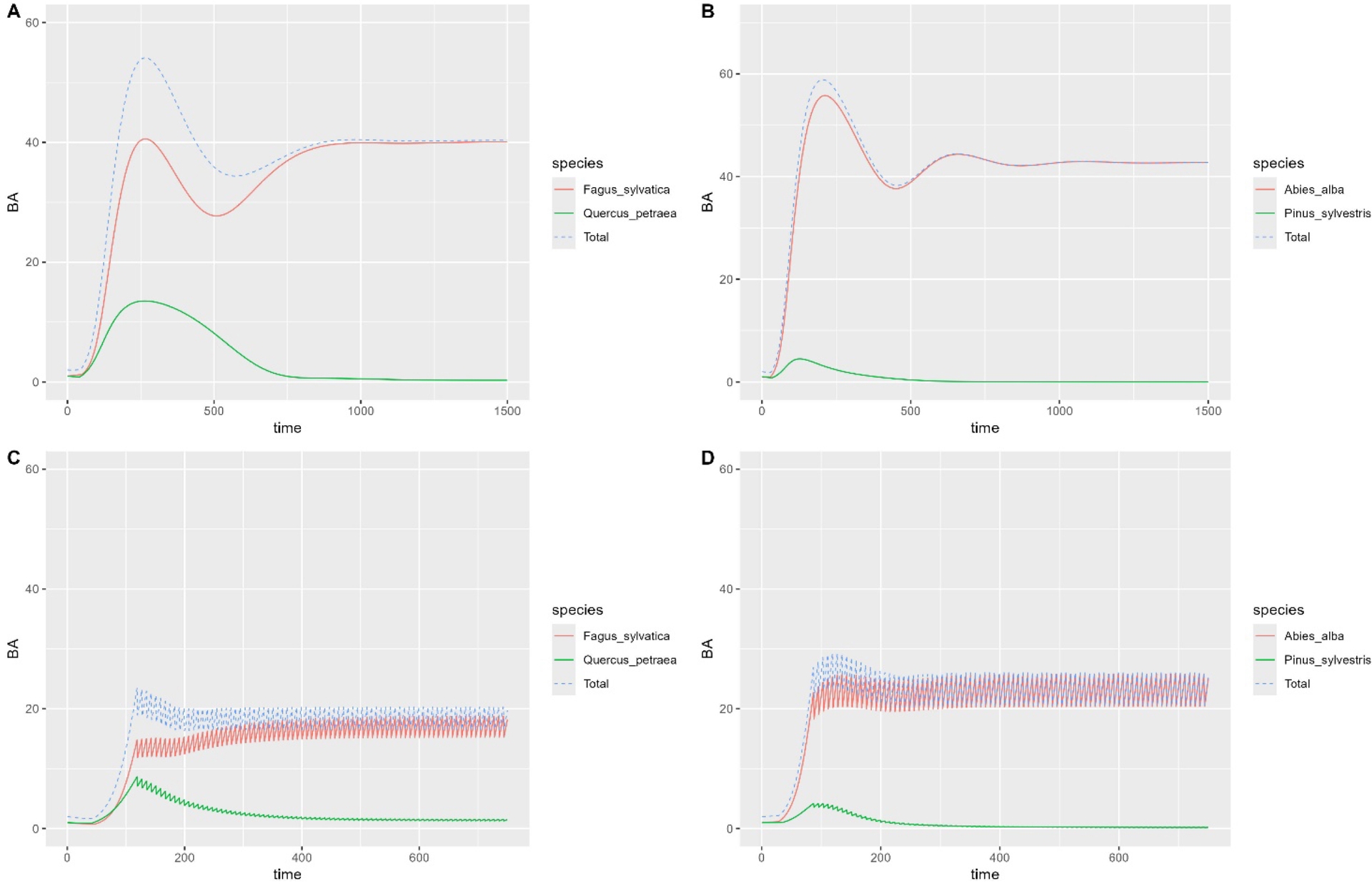
Paramètres utilisés pour les simulations réalisées avec le modèle MATREEX (sylvicultures irrégulières). Seul le diamètre d’exploitabilité dha varie entre les deux mélanges : 62,5 cm pour le mélange de hêtre commun et de chêne sessile et 52,5 cm pour le mélange de sapin pectiné et de pin sylvestre. ΔBAmin : surface terrière minimale de prélèvement lors d’une coupe, fixée à 3 m2⋅ha−1. Un événement de coupe est réalisé tous les 8 ans. Le terme gros bois désigne ici les arbres ayant dépassé le diamètre d’exploitabilité (arbres récoltables).
| Sylviculture | BAtarget (m2⋅ha−1) | hmax | a | dth (cm) | Pmax | ΔBAmin (m2⋅ha−1) |
|---|---|---|---|---|---|---|
| Référence | 15 | 0,8 | 1 | 17,5 | 0,25 | 3 |
| Coupe préférentielle de l’espèce la plus abondante1 | 15 | 0,8 | 1 → 4 | 17,5 | 0,25 | 3 |
| Diminution de la densité objectif2 | 15 → 10 | 0,8 | 1 | 17,5 | 0,25 | 3 |
| Diminution des prélèvements des gros bois3 | 15 | 0,8 → 0,1 | 1 | 17,5 | 0,25 | 3 |
1En référence à la fréquence-dépendance négative et la théorie de Chesson.
2En référence à la non-linéarité relative et les théories de la niche de succession et de Chesson.
3En référence à la théorie du partitionnement foliaire et le compromis survie adulte-recrutement.
4.3. Sylvicultures favorisant la coexistence
Afin d’étudier l’effet de la sylviculture sur la coexistence, nous avons testé trois modalités d’action liées aux trois théories présentées précédemment :
- augmentation du prélèvement relatif de l’espèce la plus abondante en référence à la théorie de Chesson (fréquence-dépendance négative) avec une diminution de l’aptitude (fitness) de l’espèce compétitive lorsqu’elle devient abondante. À noter que l’idée de maintenir les mélanges en ciblant préférentiellement dans les coupes les espèces les plus abondantes a déjà été avancée par certains auteurs (ex. Wohlgemuth et al., 2002) ;
- diminution de la surface terrière cible après coupe en référence aux théories de Chesson (non-linéarité relative) et de la niche de succession (compromis croissance en pleine lumière – survie à l’ombre) avec un taux de perturbation plus élevé censé favoriser l’espèce la moins compétitive ;
- diminution du taux de prélèvement des arbres ayant atteint le diamètre d’exploitabilité (arbres récoltables) en référence à la théorie du partitionnement foliaire avec une amélioration de la durée de survie des arbres adultes de l’espèce peu compétitive, en supposant un taux de recrutement (incluant fécondité, croissance et survie juvéniles) faible de cette espèce en situation de compétition.
Le Tableau 1 indique les paramètres utilisés pour le plan de simulation.
5. Résultats
Le prélèvement préférentiel de l’espèce la plus abondante influence de manière significative le taux de mélange dans les deux types de mélange étudiés (Figure 3). Le chêne sessile dépasse 20 % de la surface terrière à l’équilibre lorsque le paramètre de fréquence-dépendance négative est supérieur ou égal à 1,4 (ex. prélèvement de 64 % de la surface terrière de l’espèce la plus abondante si elle occupe 60 % de la surface terrière dans le peuplement). Dans le cas du pin sylvestre, l’effet s’avère moins important et cette espèce dépasse 20 % lorsque le paramètre de fréquence-dépendance négative est supérieur ou égal à 1,6 (ex. prélèvement de 66 % de la surface terrière de l’espèce la plus abondante si elle occupe 60 % de la surface terrière dans le peuplement).
Effet de l’intensité de la fréquence-dépendance négative (variation du paramètre a, de 1 à 4) sur la proportion de l’espèce peu compétitive dans les deux mélanges étudiés : (A) hêtre commun et chêne sessile, (B) sapin pectiné et pin sylvestre.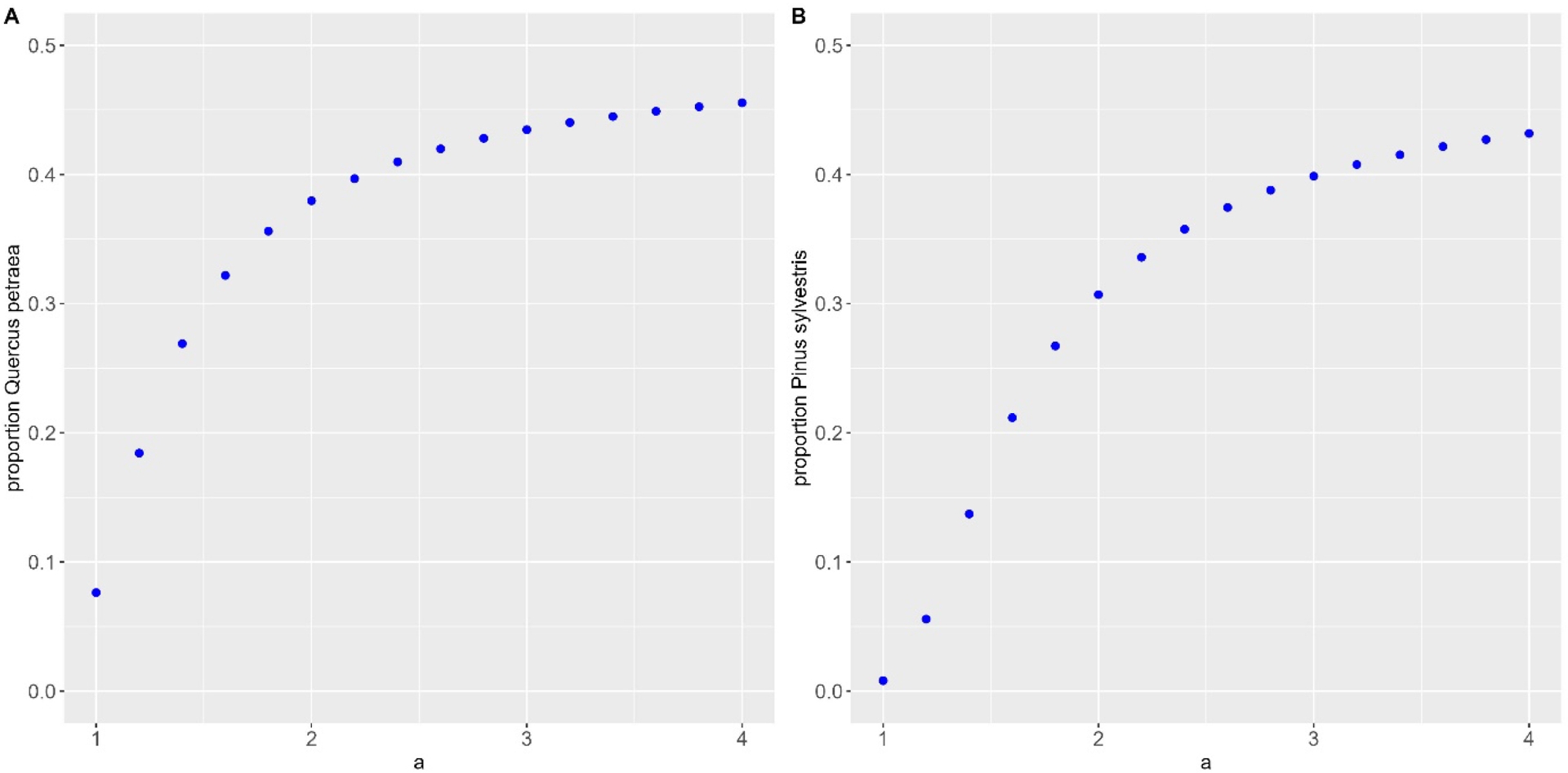
Dans le cas d’une baisse de densité, on n’observe aucun effet dans le cas du mélange de sapin pectiné et de pin sylvestre et un très léger effet positif et saturant dans le cas du mélange de chêne sessile et de hêtre commun (Figure 4). Le chêne sessile ne dépasse guère 13 % de la surface terrière totale, ce qui est très faible.
Effet de la diminution de densité objectif (variation du paramètre BAtarget) sur la proportion de l’espèce peu compétitive dans les deux mélanges étudiés : (A) hêtre commun et chêne sessile, (B) sapin pectiné et pin sylvestre.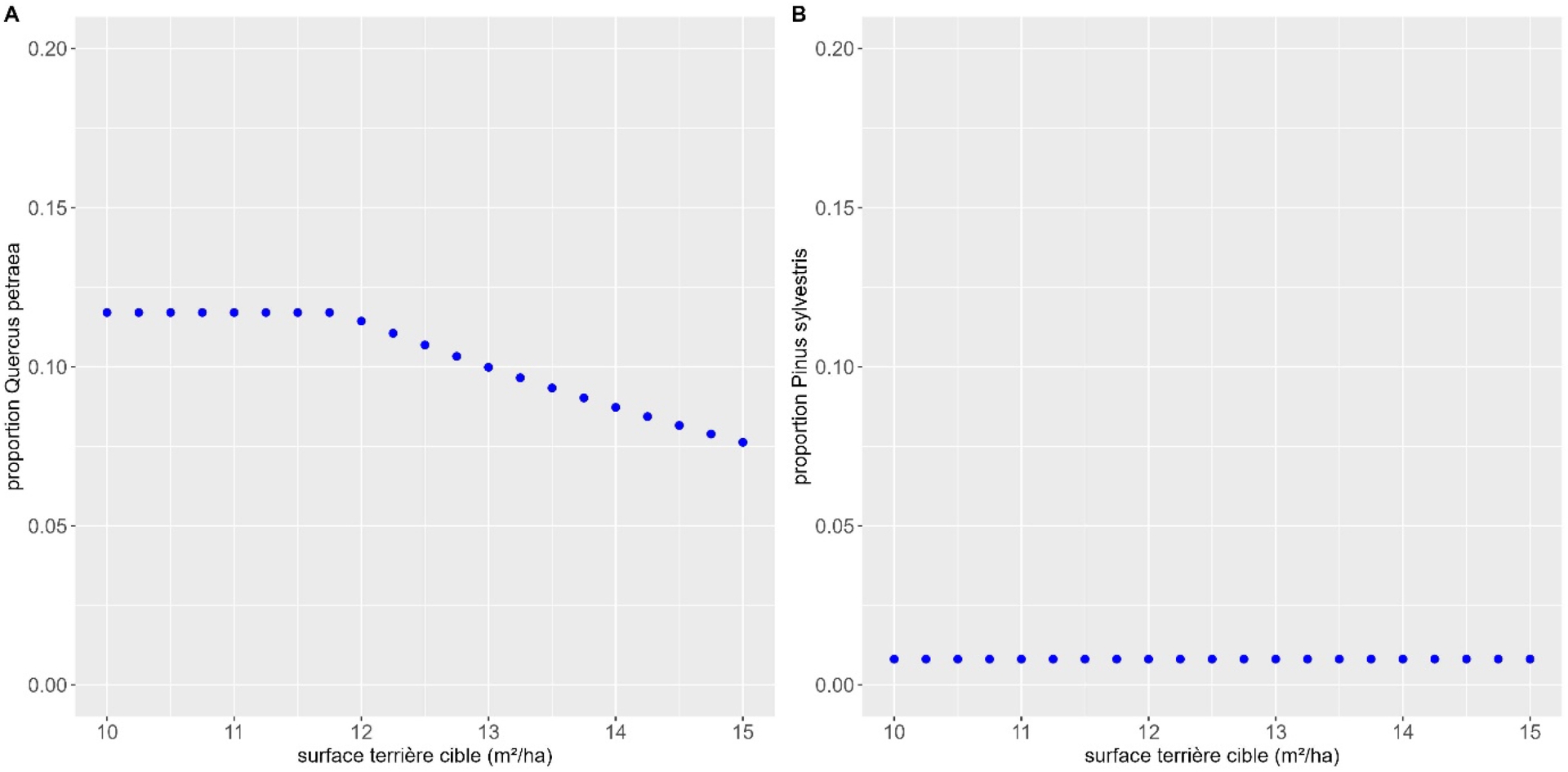
On peut observer (Figure 5) un très faible effet également pour la diminution des prélèvements des arbres récoltables avec une proportion qui reste inférieure à 10 % pour le pin sylvestre et qui monte certes à 44 % dans le cas du chêne sessile, mais pour des valeurs très faibles de hmax (0,1 : au maximum 10 % des arbres ayant dépassé le diamètre d’exploitabilité sont prélevés). Pour hmax supérieur à 0,2, la proportion de chêne sessile ne dépasse pas 10 %.
Effet d’une diminution des taux de prélèvement des arbres récoltables (appelés gros bois) (hmax) sur la proportion de l’espèce peu compétitive dans les deux mélanges étudiés : (A) hêtre commun et chêne sessile, (B) sapin pectiné et pin sylvestre.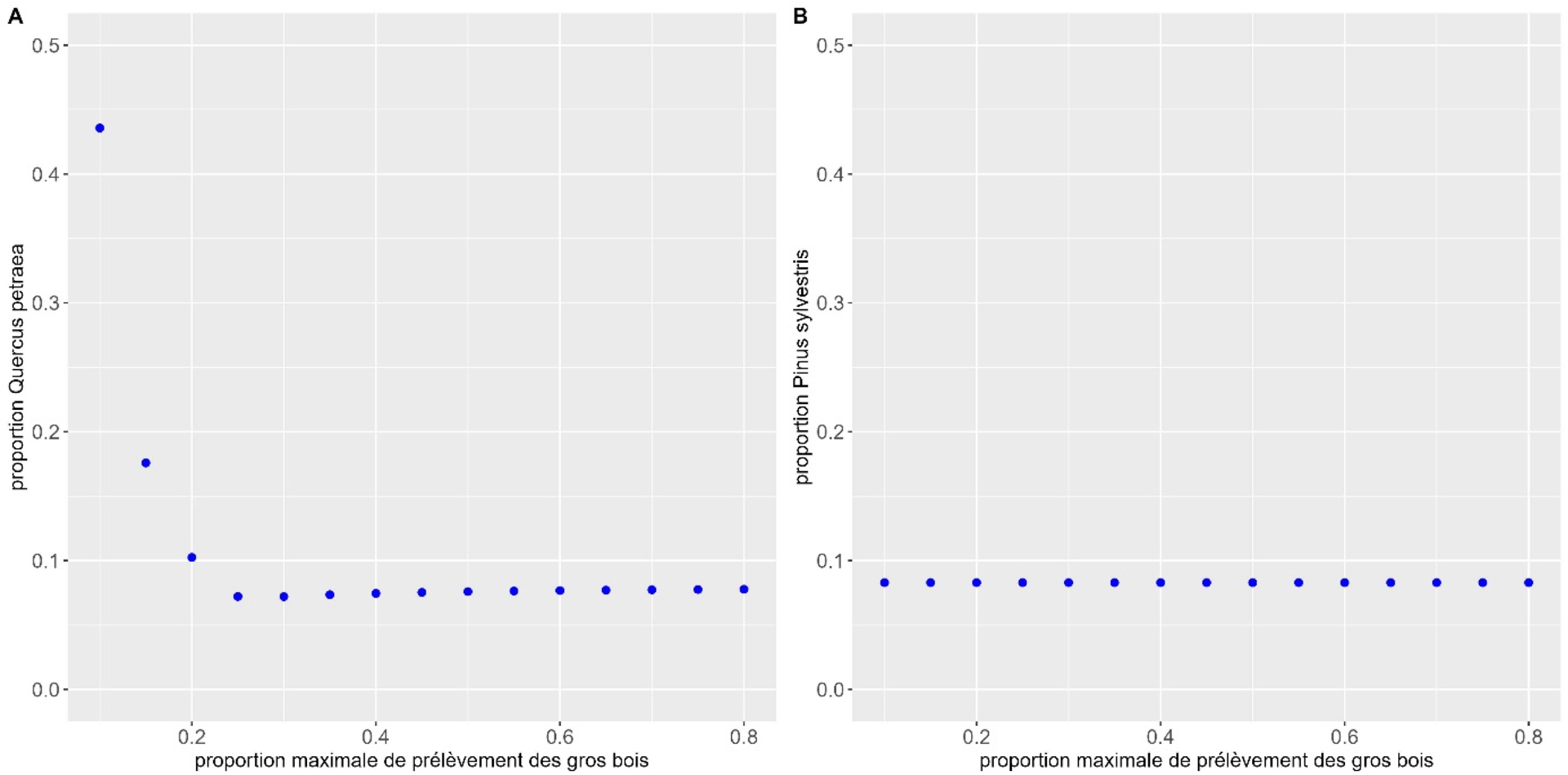
6. Discussion et conclusions
Dans MATREEX, la sélection préférentielle de l’espèce la plus abondante (ou fréquence-dépendance négative des coupes) se révèle être une stratégie très efficace pour favoriser la coexistence des espèces, en accord avec Wohlgemuth et al. (2002) et la théorie de Chesson (2018). Il s’agit en outre d’une action sylvicole aisée à mettre en œuvre et d’ores et déjà mise en application de manière empirique par les gestionnaires forestiers. En comparaison, la diminution de la surface terrière cible et la diminution des prélèvements des gros bois ne s’avèrent guère efficaces pour les deux mélanges envisagés. En outre, les valeurs de BAtarget et hmax permettant d’améliorer légèrement la proportion des espèces peu compétitives sont irréalistes en gestion, car elles conduiraient à des pertes de production pour les faibles valeurs de surface terrière (O’Hara and Gersonde, 2004) dans le premier cas ou à des arbres de trop gros diamètre pour la récolte dans le deuxième (Lafond et al., 2017).
Le modèle MATREEX comporte un certain nombre de limites qu’il est important de souligner dans cette étude et qui nous incitent à être prudents sur la portée des résultats présentés.
En premier lieu, la compétition subie par une espèce est principalement guidée par la surface terrière totale. Elle ne tient pas compte de la composition des compétiteurs. Or, plusieurs études ont montré que l’effet compétitif sur la croissance et la mortalité dépend des traits des espèces (Beauchamp et al., 2025; Kulha et al., 2023; Kunstler, Falster et al., 2016). Une étude récente de modélisation a d’ailleurs révélé que les différences de réponses et d’effets entre espèces peuvent être déterminantes pour la coexistence (T. S. Kohyama, Shigesada et al., 2025). Le modèle est par ailleurs entièrement déterministe, sans composante stochastique. En particulier, la stochasticité démographique n’est pas représentée dans la version utilisée. De ce fait, il n’est pas possible d’évaluer la coexistence des espèces sensu stricto (Hening et al., 2021). Ce point doit toutefois être relativisé dans le cadre de notre étude : pour un gestionnaire, l’enjeu principal réside dans l’estimation du taux minimal de mélange permettant de maintenir les espèces dans le peuplement, un objectif pour lequel le modèle se révèle opérationnel.
MATREEX n’intègre pas encore l’ensemble des éléments nécessaires pour reproduire certains compromis interspécifiques. Dans la version utilisée, les espèces ne diffèrent ni par l’âge ni par la taille auxquels elles commencent à produire des graines (Journé et al., 2024). Or, ce trait peut fortement influencer le compromis recrutement–survie, en particulier si l’espèce ayant la plus faible durée de survie atteint plus rapidement la maturité sexuelle. L’intégration en cours dans le modèle de la dépendance de la fécondité à la taille des individus (diamètre minimal de reproduction) permettra d’explorer plus finement le rôle de ce compromis dans la coexistence des espèces.
Par ailleurs, le modèle n’intègre pas encore de flux de graines provenant de l’extérieur du peuplement (processus d’immigration). Pourtant, ce processus — lié à la dynamique du pool régional d’espèces — peut fortement influencer la trajectoire des espèces, notamment en interaction avec les perturbations du couvert (Cordonnier, Courbaud et al., 2006). L’intégration de ce mécanisme constitue également une voie d’amélioration en cours d’implémentation. Enfin, notre approche se concentre sur l’échelle du peuplement (un seul patch), alors que d’autres mécanismes de coexistence peuvent émerger à des échelles spatiales supérieures (plusieurs patchs). Dans ce cas, la dynamique spatiale des perturbations ainsi que la dynamique de dispersion et de recolonisation des espèces peuvent générer des mécanismes additionnels de coexistence (Chesson, 2000a; Gravel et al., 2010; T. Kohyama, 1993).
Malgré ces limites, et sous réserve de prendre toutes les précautions nécessaires dans l’interprétation des résultats, notre exemple illustre que la sylviculture peut être utilement assimilée, en théorie écologique, à un « agent biotique » doté de caractéristiques propres : dépendance à la densité, spécificité envers certaines espèces, sélection préférentielle de classes de diamètre, fréquence de retour des interventions, intensité des prélèvements, etc. La particularité de la sylviculture réside dans sa capacité à agir simultanément sur plusieurs leviers, en reproduisant — de manière parallèle ou séquentielle — différents mécanismes de coexistence. Elle possède en outre un caractère adaptatif : le gestionnaire peut diagnostiquer à tout moment des déviations de trajectoire par rapport aux objectifs fixés et ajuster ses interventions en conséquence.
Notre cas d’étude met également en évidence la capacité de la sylviculture à influencer de manière significative les interactions entre espèces, en modulant les prélèvements selon leurs abondances relatives, y compris dans des mélanges traditionnellement considérés comme difficiles à conduire. Ce résultat demeure toutefois très théorique et appelle à des approfondissements. D’une part, il serait pertinent d’identifier plus précisément les stades de développement auxquels les prélèvements différenciés sont les plus efficaces, par exemple via des travaux ciblés ou des éclaircies sélectives. D’autre part, la validité de ces conclusions mériterait d’être évaluée au moyen d’expérimentations in situ (Cordonnier, 2019).
Remerciements
Nous remercions Bjoern Reineking, Benoît Courbaud et Arnaud Guyennon pour les discussions enrichissantes sur les modèles de coupes. Nous remercions également tous les collègues écologues et sylviculteurs avec lesquels nous avons pu échanger de manière passionnante sur les liens entre théories écologiques et sylvicultures.
Déclaration d’intérêts
Les auteurs ne travaillent pas, ne conseillent pas, ne possèdent pas de parts, ne reçoivent pas de fonds d’une organisation qui pourrait tirer profit de cet article, et n’ont déclaré aucune autre affiliation que leurs organismes de recherche.
Usage d’une intelligence artificielle générative
L’auteur principal de l’article a utilisé une IA générative (Copilot) uniquement pour améliorer la formulation de certains paragraphes rédigés. Aucune IA générative n’a été employée pour produire du texte ex nihilo, ni pour générer des images, tableaux ou graphiques.