



 CC-BY 4.0
CC-BY 4.0
1. Introduction
La rapidité et l’ampleur des changements climatiques actuels exercent une pression sans précédent sur les forêts et leur biodiversité, déjà soumises par ailleurs à d’autres facteurs de changement comme, par exemple, les retombées atmosphériques d’azote ou l’augmentation des densités en ongulés (Trumbore et al., 2015; Segar et al., 2022). Compte tenu de la relation étroite entre la biodiversité et le fonctionnement des écosystèmes d’une part (Tilman et al., 2014; Mori et al., 2023), et entre les fonctions écosystémiques et les biens et services fournis aux sociétés humaines d’autre part (Brockerhoff et al., 2017), conserver des écosystèmes forestiers diversifiés et fonctionnels, en état de rendre de multiples services, suppose leur adaptation aux pressions naturelles et anthropiques et un accroissement de leur résilience. Plusieurs leviers sont fréquemment évoqués, depuis l’évolution libre jusqu’au remplacement des essences actuellement en place par d’autres, supposées préadaptées à des climats plus chauds et secs, en passant par la migration assistée de génotypes ou d’espèces (Xu and Prescott, 2024; Koralewski et al., 2015). Dans tous les cas, de nouveaux assemblages d’espèces vont se mettre en place en réponse aux changements climatiques et environnementaux, spontanément ou artificiellement, induisant de nouveaux réseaux d’interactions entre les différentes composantes des écosystèmes et, par conséquent, de nouvelles dynamiques. Un postulat fréquemment mis en avant par une partie de la filière forêt-bois française pour influencer les politiques publiques est que les changements climatiques sont trop rapides pour que la forêt puisse s’adapter spontanément (Nepipvoda, 2025). Cette assertion, bien peu argumentée sur le plan scientifique, sert souvent à justifier un interventionnisme débridé et dispendieux, pouvant aller jusqu’au remplacement d’écosystèmes forestiers multifonctionnels par des monocultures ligneuses artificielles dans l’espoir de préserver la fonction économique de la forêt sur le court terme, au détriment de la biodiversité et des autres services écosystémiques. Cette attitude, surtout prônée par des coopératives peu scrupuleuses, occulte la dimension écosystémique de la forêt et l’importance de la biodiversité, voire la considère comme un frein à la gestion forestière (Gouvernement français, 2023). Dans cette contribution, il s’agit de montrer pourquoi la biodiversité est indispensable au bon fonctionnement des écosystèmes forestiers, y compris pour leur fonction économique, et comment elle peut servir de levier pour l’adaptation des forêts aux changements climatiques. Pour aborder ce sujet, un préalable nécessaire est de comprendre comment cette biodiversité s’est mise en place, à la fois dans le temps et dans l’espace. Ensuite, il s’agit d’analyser les types de pressions auxquels sont soumis les écosystèmes forestiers et la biodiversité qu’ils hébergent pour appréhender les réponses aux forçages climatiques. À partir de là, en fonction des différents scénarios climatiques et des stratégies de gestion forestière, il sera possible d’anticiper les possibles trajectoires à venir des forêts et de leur biodiversité et d’esquisser un rapport bénéfices/risques pour chacune de ces stratégies.
2. Forêt et biodiversité : un héritage
La biodiversité correspond à l’ensemble des espèces vivantes, de leurs gènes et des écosystèmes dans lesquels elles se trouvent (Barbault, 1997). Elle constitue l’héritage d’environ 3,8 milliards d’années d’histoire de l’évolution, c’est-à-dire qu’elle s’est mise en place très progressivement sur les temps longs de l’évolution, au gré de phénomènes de spéciation et d’extinction sous dépendance, au moins partielle, de conditions environnementales changeantes, en particulier climatiques (Bromham and Cardillo, 2019). Actuellement, on estime qu’entre 68 et plus de 80 % de la biodiversité mondiale, selon le groupe taxonomique considéré, se trouvent en forêt, avec par exemple environ 60 000 espèces connues d’arbres (FAO and UNEP, 2020).
À une échelle globale, la biodiversité actuelle est le résultat de l’histoire de la coévolution des êtres vivants dans les différentes régions du globe sous des conditions environnementales particulières ; et de l’histoire géologique de la planète qui a mis en place ces régions (Bromham and Cardillo, 2019). Par exemple, les îles, qui constituent des creusets évolutifs séparés des continents, hébergent classiquement un bien plus grand nombre d’espèces endémiques que ces derniers (MacArthur and Wilson, 2001). De même, les lointains ancêtres des êtres vivants actuels sont apparus sur un continent unique, mais leurs descendants et les descendants de ces descendants ont été soumis à des pressions évolutives différentes, chacun dérivant sur son radeau flottant après la fragmentation de la Pangée, puis des supercontinents Gondwana et Laurentia (Bromham and Cardillo, 2019). Histoire géologique et histoire évolutive sont donc intimement liées et ont façonné, non seulement la biodiversité actuelle, mais aussi les réseaux d’interactions entre les différentes espèces (incluant p. ex. les relations de prédation, de compétition, de mutualisme, de parasitisme). Le résultat de cette coévolution entre espèces sur un territoire biogéographique donné peut être vu comme un équilibre dynamique fragile reposant sur des interactions écologiques (p. ex. interaction proie-prédateur), voire génétiques (p. ex. interaction hôte-parasite, symbiose) entre espèces.
Toujours à l’échelle de la planète, les grands types de forêt (biomes équatorial, tropical, tempéré, boréal …) sont naturellement agencés selon une zonation latitudinale (et altitudinale) dont le déterminisme est principalement d’ordre climatique (Gaston, 2000). Puisque nous sommes dans une période de réchauffement climatique global, il n’est pas étonnant que la répartition spatiale de ces biomes forestiers à l’échelle de la planète subisse actuellement des modifications, avec, par exemple, une migration de la forêt vers les hautes latitudes et les hautes altitudes (Bonannella et al., 2023). Pour un même type de biome, on peut avoir des répartitions latitudinales sensiblement différentes, car l’histoire de l’évolution combinée à l’histoire climatique et à l’histoire géologique a permis la différenciation et le maintien d’espèces adaptées à des climats plus contraignants sur un continent, mais pas sur un autre. Par exemple, le sud des États-Unis et le Mexique hébergent des forêts sèches à canopée fermée structurées par diverses essences feuillues et résineuses, alors qu’à la même latitude et sous des climats comparables, il n’y a guère d’écosystèmes forestiers sur le continent africain.
À une échellerégionale, la biodiversité forestière dépend d’abord du réservoir régional d’espèces qui a été façonné spontanément par les processus globaux précédemment évoqués, mais qui a été largement modifié par l’homme au cours du temps, par l’introduction d’espèces venant d’autres régions (espèces exotiques) et l’élimination d’espèces jugées indésirables (Decocq et al., 2021). Mais d’autres facteurs interviennent, comme les conditions stationnelles (climat, nature du substrat géologique, type de sol, topographie, etc.) et les interactions biotiques (compétition, symbiose, parasitisme, prédation, etc.) qui, elles-mêmes, sont souvent modifiées par les activités humaines. Dans le cas des écosystèmes forestiers, il faut ajouter trois autres facteurs :
- les activités humaines, qui agissent sur la biodiversité, soit directement en privilégiant certaines essences au détriment d’autres ou en supprimant un compartiment (p. ex. la strate arbustive dans certaines plantations ou dans certaines futaies régulières), soit indirectement en modifiant les conditions stationnelles (par exemple drainage, amendement ou fertilisation du sol).
- l’ancienneté de la forêt, qui renvoie à la continuité forestière d’un site (Hermy and Verheyen, 2007) : les écosystèmes forestiers qui sont en place depuis plusieurs siècles à millénaires, qu’ils soient gérés ou pas, ont eu davantage de temps pour accumuler des espèces. La biodiversité y est donc plus élevée que dans des forêts récentes, c’est-à-dire sur d’anciennes terres agricoles qui ont été boisées au cours des derniers siècles à décennies.
- la maturité des peuplements, qui renvoie à l’âge des arbres de la canopée et de l’écosystème qu’ils structurent : en l’absence d’intervention humaine, un écosystème forestier consiste le plus souvent en une mosaïque de phases d’un cycle sylvigénétique (régénération, croissance, maturation, vieillissement, sénescence, effondrement), chacune avec son propre cortège d’espèces (Decocq et al., 2021). La suppression de certaines phases dans les forêts gérées (notamment celles de vieillissement, de sénescence et d’effondrement), s’accompagne mécaniquement de la disparition des espèces associées à ces phases, donc d’une réduction de la biodiversité totale de l’écosystème. Les écosystèmes forestiers en phase de sénescence en particulier, sont connus pour héberger un cortège d’espèces très spécialisé et très riche, notamment en organismes décomposeurs (p. ex. les insectes saproxyliques et leurs prédateurs, les champignons saprotrophes, les bactéries capables de métaboliser des substrats récalcitrants).
3. Forêt et biodiversité : un lien fonctionnel organique
Quelle que soit l’échelle d’appréhension, la biodiversité forestière ne se limite pas au nombre d’espèces d’arbres : elle inclut toutes les espèces de l’écosystème (végétales, animales, microbiennes) et leur diversité génétique. L’ensemble des êtres vivants interagissant au niveau de la partie aérienne de l’écosystème forme son « réseau vert » : c’est la partie visible de l’iceberg « forêt » (Wardle et al., 2005). L’ensemble des êtres vivants interagissant au niveau de la partie souterraine en forment le « réseau brun » : c’est la partie immergée du même iceberg « forêt », qui renferme pourtant la majeure partie de la biodiversité forestière, au rôle fonctionnel crucial (Moore et al., 2004). Réseau brun et réseau vert interagissent en permanence, entre eux et avec l’environnement qui les entoure, et forment le support du fonctionnement de la machinerie écosystémique (Zou et al., 2016).
La biodiversité est un attribut crucial de la forêt en contexte de changements environnementaux, car il existe une relation connue de longue date des scientifiques entre la diversité fonctionnelle et la résilience d’un écosystème, c’est-à-dire entre la diversité des interactions au sein des réseaux verts et bruns (et, mécaniquement, le nombre d’interactions augmente avec le nombre d’espèces) et la capacité d’un écosystème partiellement détruit par une perturbation à recouvrer son fonctionnement initial malgré l’éventuelle perte d’espèces (Isbell et al., 2015) (Figure 1). Derrière cette relation se trouve le fait que le nombre d’espèces présentes dans un écosystème est virtuellement illimité, tandis que le nombre de fonctions écologiques participant au fonctionnement de l’écosystème est lui limité. Ainsi, le nombre de fonctions écologiques augmente mécaniquement avec le nombre d’espèces jusqu’à atteindre un seuil, lorsque toutes les fonctions écologiques possibles sont remplies (de Bello et al., 2021). On dit que la complémentarité fonctionnelle des espèces augmente pour atteindre une valeur maximale, qui dépend du type d’écosystème. Si le nombre d’espèces continue d’augmenter, le nombre de fonctions n’augmente plus, mais plusieurs espèces se retrouvent à jouer le même rôle : on accroît ainsi la redondance fonctionnelle de l’écosystème, qui est finalement une assurance « résilience ». En effet, quand un écosystème forestier subit une perturbation (i.e. un évènement ponctuel, survenant de manière imprévisible, qui détruit tout ou partie d’un écosystème (Grime, 1977), comme une tempête, un incendie, une coupe), qui provoque la perte de plusieurs espèces, on comprend aisément que dans un écosystème avec une grande biodiversité, donc beaucoup d’espèces redondantes, la probabilité de perdre en même temps des fonctions écologiques est nulle à faible ; alors que dans un écosystème avec une faible biodiversité, donc peu ou pas d’espèces redondantes, cette probabilité est beaucoup plus élevée, entrainant une altération de la complémentarité fonctionnelle et, donc, du fonctionnement de l’écosystème : certaines fonctions assurées par une seule espèce disparaissent. Un exemple bien connu est celui de la disparition des grands prédateurs en forêt, qui a permis la prolifération des herbivores et l’augmentation de la pression sur les phases de régénération.
Représentation schématique de la relation entre diversité fonctionnelle et résilience d’un écosystème. Chaque espèce possède une niche écologique fonctionnelle. Si le nombre d’espèces augmente (valeur croissante sur l’axe des abscisses), mécaniquement, le nombre de fonctions dans l’écosystème augmente (valeur croissante sur l’axe des ordonnées). Si le nombre d’espèces est virtuellement illimité, le nombre de fonctions écologiques est, lui, en nombre limité (la niche fonctionnelle des espèces dans un espace à une dimension est représentée par une parabole occupant une partie de cet espace fonctionnel). Dans un premier temps, si on ajoute une à une des espèces dans un système qui en serait dépourvu au départ (point O), le nombre de fonctions ajoutées augmente de manière plus ou moins linéaire avec le nombre d’espèces, jusqu’à atteindre une valeur maximale (point F) où toutes les fonctions possibles sont remplies (c’est-à-dire que toutes les niches fonctionnelles sont occupées). Sur cette première partie de la courbe, l’augmentation du nombre d’espèces permet donc d’accroître la complémentarité fonctionnelle entre ces espèces et donc la multifonctionnalité de l’écosystème qui les héberge. Au-delà de cette valeur maximale, si le nombre d’espèces continue à augmenter, le nombre de fonctions reste stable (c’est-à-dire qu’il n’y a plus d’augmentation possible de la complémentarité fonctionnelle), mais la probabilité augmente qu’une même fonction soit remplie par plusieurs espèces différentes : on accroît donc la redondance fonctionnelle entre les espèces au sein de l’écosystème (c’est-à-dire que leurs niches fonctionnelles se superposent). En cas d’une survenue d’une perturbation dont l’intensité est capable de détruire un certain nombre d’espèces (flèche rouge) : si la redondance fonctionnelle est faible (A), la complémentarité fonctionnelle est fortement réduite (c’est-à-dire qu’on se déplace sur la pente de la courbe), d’où un fonctionnement très altéré de l’écosystème, qui perd certaines de ses fonctions ; si, en revanche, la redondance fonctionnelle est élevée (B), malgré la perte du même nombre d’espèces, la complémentarité fonctionnelle reste maximale (c’est-à-dire qu’on se déplace sur l’asymptote de la courbe), car il existe des espèces redondantes capables de remplacer les espèces perdues et de maintenir le fonctionnement de l’écosystème à l’identique de ce qu’il était avant la perturbation. Il existe donc une relation forte entre redondance fonctionnelle des espèces et résilience aux perturbations de l’écosystème (symbolisée par un gradient de couleur d’intensité croissante de la gauche — faible résilience — vers la droite — résilience élevée).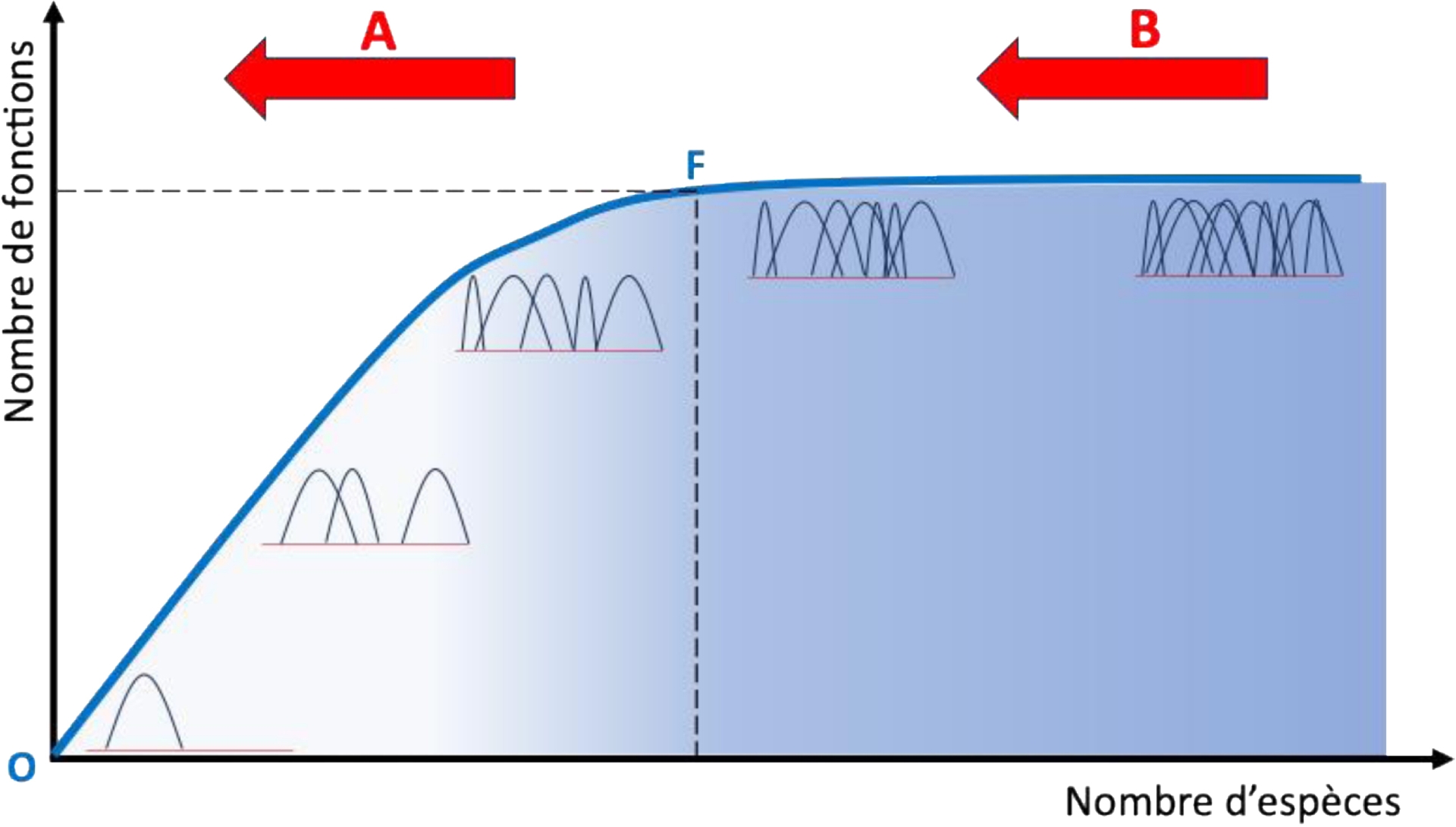
Le fonctionnement « optimal » d’un écosystème forestier est par conséquent intimement lié à la biodiversité qu’il héberge, et nombre des services écosystémiques rendus dépendent, directement ou indirectement, de cette même biodiversité (Brockerhoff et al., 2017).
4. Forêt et biodiversité : quelle(s) réponse(s) aux changements climatiques ?
La forêt se reconnaît à ses arbres. Ce sont les espèces arborescentes qui forment la canopée, structurent les écosystèmes forestiers, créent les conditions environnementales — notamment microclimatiques — favorables aux espèces du sous-bois, font entrer la plus grande part de l’énergie qui s’écoule dans l’écosystème — par conversion de l’énergie solaire en énergie biochimique utilisable par les organismes hétérotrophes qui le peuplent —, et fournissent une grande quantité de matière organique aux processus de minéralisation des différents cycles biogéochimiques qui assurent la circulation de la matière et le recyclage des nutriments. La densité du couvert arboré variant naturellement au cours du cycle sylvigénétique, la biodiversité et l’organisation structurelle et fonctionnelle de l’écosystème ne sont pas stables dans le temps, mais changent de manière cyclique ou, plus exactement, de manière « spiralée ». Ces changements cycliques se superposent en effet à des changements linéaires, c’est-à-dire à une trajectoire directionnelle de changements plus ou moins lents et progressifs comme, typiquement, le réchauffement climatique global (Zellweger et al., 2020). Si bien que le cycle sylvigénétique ne revient jamais tout à fait à son point de départ, mais à une phase pour laquelle les conditions environnementales ne sont plus tout à fait les mêmes que celles qui prévalaient pour la même phase du cycle précédent. Par conséquent, la biodiversité change spontanément au cours de cette trajectoire spiralée, à la fois de manière cyclique et linéaire, comme le montrent de nombreuses études de suivi à long terme (p. ex. De Frenne et al., 2013; Bernhardt-Römermann et al., 2015; Perring et al., 2018). Par conséquent, il serait illusoire de penser qu’une gestion forestière « adaptative » pourrait maintenir les forêts dans leur état actuel.
À l’heure où les changements climatiques s’accélèrent, les arbres qui structurent les écosystèmes forestiers sont soumis à deux ensembles de pressions d’intensité croissante : des stress, c’est-à-dire des contraintes chroniques qui limitent l’accumulation de biomasse, en particulier un stress thermohydrique de fréquence et d’intensité croissantes (sécheresses et épisodes caniculaires) ; et des perturbations, c’est-à-dire des évènements ponctuels, souvent imprévisibles, qui détruisent une partie de la biomasse (p. ex. tempête, feu de forêt, attaque de ravageurs) (Grime, 1977). Or, la théorie écologique montre qu’il existe des compromis évolutifs (« tradeoffs ») sur les traits fonctionnels qui permettent à une plante (ou à un organisme en général) d’avoir un phénotype adapté à son environnement, si bien que cette plante ne peut pas être à la fois résistante à des stress intenses et résiliente à des perturbations trop fréquentes/intenses (ibid.). Transposée à l’échelle de l’écosystème, la théorie prédit qu’un écosystème forestier ne peut pas persister dans un environnement trop stressé et/ou trop perturbé ; il est progressivement remplacé par d’autres types d’écosystème de type lande, pelouse, savane, friche, etc., avec tous les intermédiaires possibles (Grime, 1977; Seidl et al., 2017). Beaucoup d’auteurs, par exemple, considèrent que la forêt préhistorique, soumise aux grands herbivores, ressemblait davantage à une savane (ou à un pâturage boisé) qu’à une forêt à canopée fermée comme on la connait aujourd’hui (Rackham, 1998; Vera, 2000). Il en est de même pour la forêt médiévale, dans laquelle les troupeaux d’animaux domestiques avaient remplacé les grands herbivores sauvages (Braunstein, 1990).
La double pression, stress thermohydriques récurrents et perturbations par les tempêtes, feux, ravageurs, etc., qui s’exerce sur la forêt française induit un dépérissement forestier important, qui se traduit par des descentes de cimes chez nombre d’essences structurantes (hêtre, chênes, sapin) et des mortalités de masse des arbres fragilisés en lien avec des attaques d’insectes (p. ex. scolyte sur épicéas) ou de champignons parasites (p. ex. chalarose du frêne) (Allen et al., 2010; Anderegg et al., 2016). Ce dépérissement forestier est associé à des changements des conditions environnementales, qui peuvent devenir défavorables à la persistance de certaines espèces et favorables à la colonisation d’autres espèces, ce qui se traduit inexorablement par des changements qualitatifs et quantitatifs de la biodiversité hébergée par l’écosystème forestier en question. Face à un environnement défavorable, une espèce ou un génotype n’a que trois échappatoires à l’extinction locale (Berg et al., 2010) : s’adapter, se réfugier (dans des habitats offrant des conditions viables, par exemple dans des fonds de vallée pour des espèces mésophiles de niveaux topographiques supérieurs) ou migrer (ce qui suppose que la vélocité des diaspores d’une espèce soit supérieure à la vitesse à laquelle son habitat actuel devient défavorable). À partir de là, il est possible de définir des stratégies de gestion forestière en contexte de changements climatiques.
5. Quelles stratégies pour la gestion forestière face aux changements climatiques ?
En France, la stratégie nationale vise à conserver des écosystèmes forestiers partout où cela est possible, pour répondre à un impératif économique (maintien d’une filière bois de qualité, compétitive et rentable) plus que pour des considérations environnementales ou sociétales (MAASA — Ministère de l’Agriculture, de l’Agro-alimentaire et de la Souveraineté alimentaire, 2021). Le principe de la multifonctionnalité passe au second plan et la biodiversité n’est plus guère considérée, malgré le consensus scientifique qui clame que les changements climatiques ne peuvent être envisagés séparément de l’érosion de la biodiversité.
Très schématiquement, il est possible de distinguer trois stratégies de gestion, qui ne doivent pas être vues comme des catégories discrètes, mais plutôt comme les régions d’un continuum allant du non-interventionnisme à la conversion de la forêt en culture d’arbres. Chacune présente des avantages et des inconvénients, tant d’un point de vue économique qu’écologique ; c’est ce que nous décrivons ci-après.
5.1. Stratégie 1 : laisser la forêt en évolution libre
Une première réaction face au dépérissement forestier induit par les changements climatiques peut être d’attendre de voir si les écosystèmes forestiers sont capables de s’adapter par eux-mêmes. Face au caractère inédit de la rapidité de ces changements climatiques, nombreuses sont les inconnues sur les capacités des organismes à s’adapter et il n’existe pas de consensus scientifique à l’heure actuelle sur ce sujet. Certains généticiens considèrent que les espèces d’arbres forestiers, au cycle de développement naturellement long, n’ont pas les capacités d’adaptation « rapide » nécessaires (Williams and Dumroese, 2013), mais ce postulat est très controversé (Kremer et al., 2025; Plomion et al., 2018). En effet les mécanismes génétiques moteurs de l’évolution (p. ex. la sélection de certains génotypes mieux adaptés au sein de populations génétiquement diversifiées, ou les mutations à valeur adaptative) agissent sur les temps longs de l’évolution et supposent de disposer d’un réservoir de gènes suffisant au sein de chaque espèce, ce qui est loin d’être évident en foresterie, puisqu’on a rarement cherché à diversifier les provenances lors de plantations. D’autres la pensent possible grâce à l’intervention de mécanismes non strictement génétiques :
- des mécanismes holobiontiques, qui font intervenir le microbiome associé aux plantes et en particulier le microbiome racinaire, qui inclut l’ensemble des microorganismes associés aux racines de la plante et qui modulent son phénotype (Allsup et al., 2023; Cosme, 2023) ;
- des mécanismes épigénétiques, où les facteurs environnementaux agissent sur la méthylation de l’ADN pour moduler l’expression du génome et qui pourraient orienter des mutations, ces mutations non aléatoires ayant alors une valeur adaptative (Amaral et al., 2020; Sow et al., 2021).
Une alternative à l’adaptation locale est la migration. Mais concernant les arbres, les espèces sont généralement peu véloces : si celles dont les diaspores sont dispersées par le vent (p. ex. aulnes, bouleaux, érables) ou les animaux (p. ex. cerisiers, sorbiers) peuvent parcourir des distances kilométriques, beaucoup d’autres ont des graines ne s’éloignant guère des pieds mères (p. ex. chênes, hêtres) (Matlack, 2005). De plus la fragmentation forestière représente un obstacle supplémentaire, surtout en plaine, à la migration spontanée des espèces (Lenoir et al., 2008). Les espèces les moins véloces doivent alors trouver un refuge à la faveur éventuelle de conditions topoclimatiques particulières, au risque de s’éteindre localement ou régionalement (Selwood and Zimmer, 2020).
Le discours selon lequel la forêt ne peut pas s’adapter spontanément quand on la laisse en évolution libre s’appuie surtout sur des données génétiques. Il doit être nuancé à la lumière des récents travaux portant sur les mécanismes holobiontiques et épigénétiques, mais surtout, les données scientifiques valables à l’échelle de l’espèce et des individus de cette espèce ne peuvent pas être transposés à l’échelle de l’écosystème forestier, où il existe normalement une diversité des essences et, au-delà des arbres, une biodiversité constituée en réseau d’interactions. Ainsi, spontanément, la biocénose de l’écosystème forestier « s’adapte » via la modification de l’abondance-dominance relative des espèces (d’où l’intérêt d’une redondance fonctionnelle élevée), l’extinction locale de certaines espèces et la colonisation par d’autres, capables d’immigrer spontanément. Les changements spontanés dans les assemblages d’espèces sont une réponse « adaptative » de la forêt en évolution libre face aux changements environnementaux.
Laisser la forêt en évolution libre permettrait a priori de maintenir une biodiversité élevée, mais nécessairement modifiée. À cet égard, les réserves biologiques intégrales peuvent jouer le rôle de laboratoire grandeur nature pour observer et suivre les capacités d’adaptation des écosystèmes forestiers : ce sont des témoins, indispensables pour les scientifiques comme pour les gestionnaires. Mais l’évolution libre au sens où nous l’entendons ici peut tout aussi bien concerner des forêts exploitées. Dans ces forêts gérées, elle implique une gestion opportuniste en fonction de ce qui se présente : les coûts de mise en œuvre sont moindres, mais il est probable que la rentabilité soit réduite aussi et les trajectoires dynamiques — donc les prévisions du type et de la quantité de bois récoltés — difficiles à anticiper. À quoi s’attendre ? Il est probable que dans de nombreuses régions d’Europe, qui vont connaître une hausse des températures et une baisse des précipitations, les forêts tempérées vont devenir de plus en plus « (sub-)méditerranéennes », avec une extension des espèces xérothermophiles au détriment des espèces mésophiles, une réduction de la hauteur des canopées et une diminution de la densité des arbres (McIntyre et al., 2015). Dans des régions littorales où il est prévu que la hausse des températures soit accompagnée d’une augmentation des précipitations, la forêt deviendra peut-être de plus en plus lauriphylle, comme le suggère le succès invasif de certaines espèces ligneuses, à l’instar du rhododendron pontique ou du laurier cerise (Casati et al., 2023). Dans les secteurs où le changement de saisonnalité s’accompagnera d’une augmentation de la pression des ongulés, la physionomie des forêts pourrait ressembler à celle de pâturages boisés, voire de « savanes » arborées (Rackham, 1998). Cette trajectoire dynamique est peut-être inexorable en l’absence d’un infléchissement du réchauffement climatique global et, même si on applique des scénarios plus interventionnistes comme ceux décrits ci-dessous, il est possible que l’échéance soit seulement retardée.
5.2. Stratégie 2 : gérer la forêt de manière à accroître sa résilience
Cette stratégie implique de réinventer l’ingénierie forestière et de mettre au point de nouveaux itinéraires techniques sylviculturaux. Pour y parvenir, il est nécessaire de mobiliser les apports des sciences écologiques, auxquels la foresterie a jusqu’ici été relativement imperméable. Les leviers sont nombreux et complémentaires. Il s’agit par exemple de :
- privilégier les mélanges d’espèces ligneuses là où c’est possible, et les classes d’âges pour « irrégulariser » les peuplements, étant donné la relation qui existe entre résistance/résilience aux perturbations et irrégularité et diversité des peuplements (p. ex. Raymond et al., 2009) ;
- accroître la complémentarité fonctionnelle parmi les essences codominantes (essences-objectifs en sylviculture), de manière à maximiser les fonctions et services écosystémiques ;
- augmenter la redondance fonctionnelle dans le peuplement en jouant sur les essences subordonnées (essences d’accompagnement en sylviculture), de manière à accroître la résilience de l’écosystème ;
- favoriser la diversité génétique en multipliant les provenances au sein de chaque espèce, de manière à constituer un réservoir suffisant pour que les mécanismes génétiques de sélection naturelle puissent intervenir (Smith et al., 2025) ;
- ménager un couvert forestier continu pour préserver le capital sol des forêts (Fedrowitz et al., 2014), notamment la biodiversité microbienne à partir de laquelle les mécanismes holobiontiques d’adaptation des essences peuvent se mettre en place.
Une autre technique fréquemment évoquée est la migration assistée, c’est-à-dire le déplacement artificiel de génotypes exogènes d’espèces indigènes (p. ex., en plantant dans le nord de la France des génotypes de hêtres venant du sud de la France, parce qu’ils sont naturellement acclimatés à des climats plus chauds et secs) (Aitken and Bemmels, 2016). Par extension, la migration assistée peut concerner des espèces au lieu de génotypes (Xu and Prescott, 2024; Koralewski et al., 2015) : on étend artificiellement l’aire de répartition d’une espèce en l’introduisant artificiellement dans une région où elle est actuellement absente, à partir d’une région adjacente du même continent où elle est naturellement présente (p. ex., en plantant dans le nord de la France du chêne vert méditerranéen). Derrière le concept de migration assistée, il est considéré que si elle en avait le temps, l’espèce migrerait spontanément vers la région d’introduction parce qu’il n’existe pas de barrière biogéographique l’en empêchant, l’homme n’intervenant que pour accélérer un processus naturel. Le terme est hélas parfois galvaudé pour être appliqué, à tort, à l’introduction d’espèces originaires d’autres continents, qui relèvent plutôt de la translocation d’espèces (Michalet et al., 2024). La technique est trop récente pour permettre une évaluation rigoureuse de son efficacité et de ses éventuelles conséquences négatives, mais elle est déjà mise en œuvre sur le territoire métropolitain, par exemple dans certaines forêts domaniales. Toutefois, certains risques ont été évoqués, comme, par exemple, celui d’exacerber les stress thermohydriques estivaux sur les espèces du sous-bois — et donc sur les régénérations ligneuses — par altération du microclimat forestier, du fait que les espèces de climats plus chauds et secs ont des surfaces foliaires réduites par rapport à celles de climats plus tempérés (ibid.).
Avec cette stratégie, il est a priori possible de maintenir une biodiversité élevée, mais modifiée. L’enjeu pour la sylviculture est de se réinventer, en apprenant à gérer la complexité (Filotas et al., 2014). C’est ce qui se fait déjà dans des systèmes de futaies irrégulières mélangées ou de futaies « jardinées ». Mais il est souhaitable de diversifier les modes de sylviculture irrégulière à couvert continu pour ménager une mosaïque d’habitats favorables à la biodiversité, par exemple en faisant varier le grain d’hétérogénéité, depuis la gestion « pied-à-pied » à la futaie irrégulière par parquets en passant par la gestion en bouquets. Une telle diversification spatiale est envisageable à l’échelle d’un massif forestier, mais peu à celle d’une parcelle.
5.3. Stratégie 3 : remplacer les écosystèmes forestiers par des plantations d’arbres (souvent exotiques) supposés mieux adaptés au climat de demain
Cette stratégie consiste à remplacer les écosystèmes forestiers par des plantations monospécifiques et équiennes d’essences à croissance rapide, qui sont soit des essences indigènes généralement résineuses (p. ex. pin maritime), soit des essences exotiques, feuillues (p. ex. chêne rouge) ou résineuses (p. ex. pin de Monterey, mélèze du Japon). L’intérêt de raccourcir la rotation est de diminuer la probabilité qu’un évènement de type catastrophique survienne avant la récolte des arbres (p. ex. tempête, incendie) qui compromettrait la rentabilité de l’investissement, qui est considérable (Drouineau et al., 2000). C’est un moyen assez efficace de conserver la fonction économique de la forêt, aux avantages certains : les itinéraires techniques sont simples et très mécanisés ; la rentabilité financière à court terme est potentiellement élevée ; le forestier conserve un contrôle total sur la composition en essences ; le choix de l’essence peut être orienté vers des espèces exotiques venant de régions plus arides où le climat actuel correspond au climat prédit dans un prochain avenir dans une région donnée de France. Cependant, cette pratique se fait au détriment de la multifonctionnalité de la forêt et de la biodiversité, car les pratiques associées (travail du sol, utilisation d’intrants, coupe à blanc, exploitation mécanisée à l’aide d’engins très lourds et polluants, utilisation d’essences exotiques, etc.) sont souvent dévastatrices pour les écosystèmes qui, rapidement, voient leur capacité de production diminuer (Wang et al., 2022). De plus, les peuplements monospécifiques sont beaucoup plus vulnérables aux aléas comme les attaques de ravageurs, les épidémies ou les incendies ; la simplicité structurelle facilite aussi les dégâts de tempête (Dobor et al., 2020). Il s’agit donc d’une sylviculture spéculative de court terme, non durable, contraire au principe de multifonctionnalité qui est pourtant parfois érigée en dogme de la gestion forestière « à la française ».
Si ce type de sylviculture peut trouver une certaine justification dans des situations particulières (réhabilitation de terrains pollués ou dégradés, boisement de terres agricoles, etc.), il ne devrait pas se substituer à la gestion durable dans les forêts anciennes, en raison de ses impacts négatifs sur la biodiversité et les services écosystémiques. Pourtant, certaines coopératives forestières n’hésitent pas à en faire la promotion, quitte à détruire sans vergogne des écosystèmes forestiers exceptionnels au prétexte que certains arbres dépérissants signeraient l’inexorable déclin de tout l’écosystème et la mise en danger d’un pan de l’économie nationale (p. ex. Le Cozic, 2023).
6. Conclusion : La biodiversité comme assurance « résilience » des forêts en environnement changeant
La biodiversité en forêt, c’est bien plus que le nombre d’espèces d’arbres, contrairement à ce que mentionnent nombre de documents de planification forestière. De la même manière, la biodiversité est bien plus que le nombre d’espèces rares ou patrimoniales, que mettent souvent en avant les défenseurs de la nature. La biodiversité doit d’ailleurs être regardée comme autre chose qu’un simple nombre d’espèces ou qu’un attribut quantitatif d’un écosystème : il est urgent de prendre en compte le rôle fonctionnel de ces espèces, et comme nous l’avons vu, la diversité, la complémentarité et la redondance fonctionnelles des communautés biotiques au sein de l’écosystème. Il faut garder en tête que la majorité de la biodiversité est invisible : elle se trouve dans le sol et, pourtant, c’est la plus importante d’un point de vue fonctionnel (Bakker et al., 2019). Quelles forêts pour quelle biodiversité dans un environnement changeant ? Face aux changements environnementaux rapides, toutes les plantes en place et en particulier les arbres ne sont pas capables de s’adapter, d’où des pics de mortalité au décours d’épisodes de stress thermohydriques majeurs. Pour autant, la présence d’arbres morts en forêt est un phénomène normal, et elle est même indispensable au bon fonctionnement de l’écosystème (Wijas et al., 2024). La mort de quelques arbres de plus à l’hectare, ne devrait pas faire conclure hâtivement au dépérissement de l’écosystème forestier tout entier et encore moins justifier des coupes à blanc et le remplacement des mélanges d’espèces indigènes par des cultures d’espèces exotiques supposées mieux adaptées au climat de demain. Au contraire, il est urgent d’adopter une gestion forestière plus fine, qui ménage les capacités d’adaptation spontanées des espèces, qui prenne réellement en compte et respecte toute la complexité de l’écosystème, et pas seulement les arbres qui le structurent. Ce n’est pas la première fois que la forêt française est soumise à d’intenses stress climatiques ; pendant le Petit Âge glaciaire, des crises de dépérissement massif l’avaient déjà affectée. Pourtant, elle s’est rapidement et spontanément reconstituée (Buridant, 2008). Il est temps d’admettre que la forêt de demain ne sera nécessairement pas celle d’aujourd’hui, pas plus que celle d’aujourd’hui dont on déplore le dépérissement ne ressemble à la forêt d’hier. Pas davantage que l’écosystème forestier qui l’héberge, la biodiversité forestière n’est figée dans le temps ; l’enjeu est de préserver son intégrité fonctionnelle pour conserver les capacités de résilience des écosystèmes aux contraintes climatiques émergentes.
Déclaration d’intérêts
L’auteur ne travaille pas, ne conseille pas, ne possède pas de parts, ne reçoit pas de fonds d’une organisation qui pourrait tirer profit de cet article, et n’a déclaré aucune autre affiliation que son organisme de recherche.