



 CC-BY 4.0
CC-BY 4.0
1. Introduction
Intraspecific variation represents in average 25% of individual trait variation found within plant communities, and 32% of the total trait variation among communities (Siefert et al., 2015, in a meta-analysis covering 629 plant communities and 36 functional traits). Intraspecific variation includes within- and between-population components. Within-population trait variation (means, variances and correlations) has three effects on population dynamics and functions that drive population adaptation in the short and the long term (Figure 1). Firstly, at any time, the complementarity among different phenotypes as well as phenotype-dependent interactions such as competition or facilitation mechanisms induce a mixture effect on population structure, ecological functions and biodiversity of associated communities (Hughes et al., 2008, for review). Secondly, within a generation, phenotypic adjustment is a dynamic process of change in the phenotypic composition of the population, not only due to ontogenesis and plasticity, but also to selective elimination of maladapted phenotypes, extending here the notion introduced by Ramírez-Valiente, Poyatos, et al. (2025). Phenotypic adjustment continuously modifies the population structure, functions and biodiversity. Thirdly, across successive generations observed at the same stage, the evolutionary response to selection is a dynamic process of change in the genetic composition of the population resulting from the combination of three mechanisms: phenotypic sorting of reproducing individuals, intercrossing of these individuals with variable reproductive success, and heredity mechanisms. Note that the time scale of genetic evolution can be similar to that of phenotypic adjustment, when phenotypic selection and reproduction continuously operate in overlapping generations. Thus, considering within-population phenotypic and genetic variation in a dynamic perspective in organisms with long life cycles, the time scales of ecological and evolutionary mechanisms may overlap (Carroll et al., 2007).
Three consequences of intraspecific trait variation within a population of long-lived organisms like trees. At all times, within-population trait variation has a mixture effect on the population structure, the ecological functions and the associated communities. In addition, within-population trait variation allows temporal changes in the phenotypic composition. Phenotypic adjustment is a dynamic change in phenotypes before reproduction occurs, due to developmental and environmental (plasticity) effects as well as selection of the adapted phenotypes. Genetic evolution is a dynamic change in genotypes due to selection and reproduction. Genetic evolution starts at the first reproduction event. Here, non-overlapping generations are represented for clarity; phenotypic adjustment and genetic evolution would be concomitant in overlapping generations.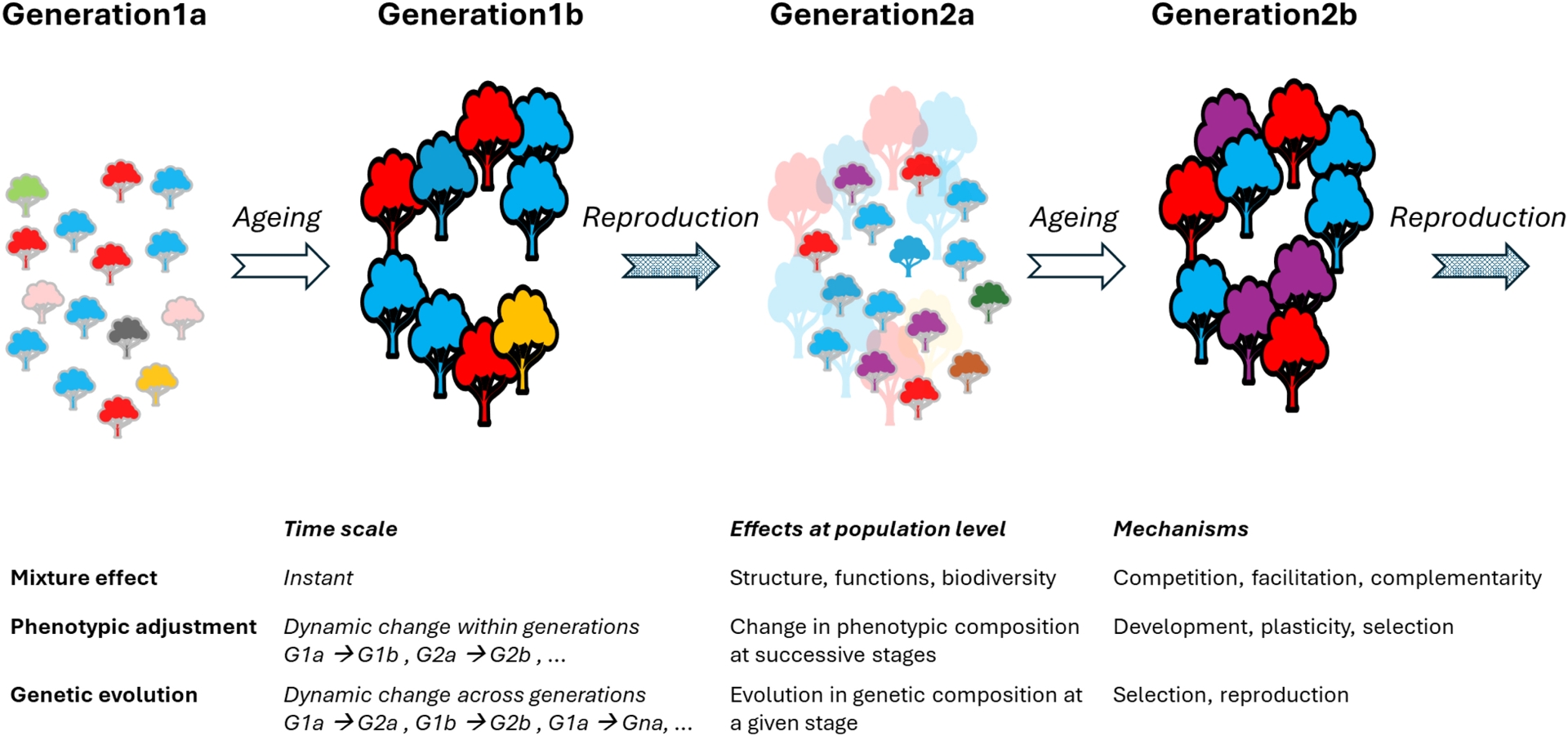
Selection first contributes to phenotypic adjustment within generations and results in genetic evolution across generations. It is a central eco-evolutionary process, whereby changes in the phenotypic and genetic composition of the population drive the ecological functions, demographic dynamics and biodiversity of the ecosystem. These changes can have feedback consequences on selection itself: for instance when the relative fitness of each individual, which drives the selection process, depends on the distribution of traits in the population, or when the intensity of external selection pressures, such as biotic or abiotic stress, is determined by and affects the population density. These two cases can be connected to soft and hard selection mechanisms, respectively (see Bell et al., 2021, for review).
Trait variation and selection are tightly linked. The evolutionary response of a trait to selection depends on (i) its initial variation in the population; (ii) its correlation to fitness; (iii) its correlation to other traits also linked to fitness; (iv) its heredity; and (v) the selection intensity (Endler, 1986; Lande, 1979). Variations and correlations of traits should be considered in a dual perspective: trait variation and covariation patterns constrain the potential response to selection and are themselves impacted by the response to selection, which Pigliucci (2008) synthesized in the concept of evolvable evolvability. Classically, trait variations and correlations between populations are used to infer on past evolutionary response to selection, e.g. local adaptation studies (Kawecki and Ebert, 2004), while within-population variation patterns are often analyzed in terms of evolutionary constraints, e.g. G-matrix studies (Arnold, 1992). However, microgeographic studies extend the exploration of past evolutionary response to selection to the within-population scale (Richardson et al., 2014), and metapopulation studies extend the analysis of potential future responses to selection to the between-deme scale (Araya-Ajoy et al., 2019).
There is empirical evidence that trait variation and evolvability are maintained within populations despite selection, even for traits tightly related to fitness (Mousseau and Roff, 1987). This observation might be surprising at first glance, but several biological and evolutionary mechanisms can explain the somehow limited impact of selection on variance and covariance patterns (Arnold et al., 2008; Kruuk et al., 2008; Pujol et al., 2018). The variability of selection processes at fine spatiotemporal scales is one explanation for the persistence of variation throughout evolution. Indeed, a single trait may be under different types of selection, which are spatially distributed within a population (in the case of microgeographic environmental or population structure heterogeneity at the spatial scale of gene flow), or which operate successively at the generational time scale. The variability of selection at a larger scale, between connected populations, may also result in the persistence of trait variation through balanced effects of migration and selection (Lenormand, 2002, for review). When selection processes operate at multiple scales in a hierarchical system, e.g. in a metapopulation combining local adaptation between demes with microgeographic adaptation within demes, they can interfere in a way that maintains the adaptive genetic diversity within each deme (van Benthem and Wittmann, 2020; Cubry et al., 2022). Thus, selection itself deserves to be considered as a variable and dynamic driver of population dynamics. Having this perspective in mind, the eco-evolutionary framework accounts for feedback between the trajectory of a population or a metapopulation and the trajectory of its selective evolutionary drivers (Griffith et al., 2016; Hanski, 2012; Lamarins et al., 2022), e.g. in a simple case where density-dependent selection affects population density.
Trees, as long-lived and sessile organisms, are particularly prone to the spatiotemporal variability of selection. Exploring the mechanisms driving the trajectory of selection, and how forest management interventions interfere with these mechanisms, helps us to better understand the trajectories of intraspecific trait variation and selection. This review consists of four sections. A first general section uses the genes–traits–fitness mapping framework to explore the expected impacts of selection on different categories of traits and syndromes. Using this general framework, a second section specifically analyzes some typical patterns of trait variation and coordination observed in trees in the dual perspective of biological constraints and response to selection. A third section reviews the main drivers of spatiotemporal variability of natural selection in forest trees. The fourth section provides an overview of emerging knowledge of the impacts of forestry interventions on natural and anthropogenic selection processes. In conclusion, possible uses of this dynamic vision of selection in forest management and conservation are illustrated.
2. Expected response of traits and genes to selection: the genes–traits–fitness mapping framework
The genes–traits–fitness mapping is a helpful conceptual framework to understand and analyze natural selection processes and their impacts on traits and genes in a population. The schematic representation in Figure 2, here specifically illustrated for trees, extends an original representation by Coulson et al. (2006) and gets inspiration from Oddou-Muratorio, Davi, et al. (2020). The map represents the functional links within and between three hierarchical spaces of variation: the space of fitness and fitness components, the space of variable traits, the space of variable genes.
The genes–traits–fitness map representing functional links across three hierarchical spaces, for trees (inspired from Coulson et al., 2006; Oddou-Muratorio, Davi, et al., 2020). In the space of fitness, tree size is an important fitness component having a direct impact on survival and reproduction. Here, the space of variable traits is illustrated with three hierarchical sublevels where multiple basic traits (e.g. water potential inducing 50% loss of hydraulic conductivity, or minimum conductance) determine the physiological functions (e.g. resistance to cavitation, or phenology), which themselves determine the integrated traits (e.g. sensitivity to drought) that are the direct drivers of fitness components. The integrated traits can result from a single functional trait or, more generally, from diverse combinations of functional traits (syndromes). We expect more frequent biological constraints on basic traits, and more impact of local selection on integrated traits (see text). Sublevels in the space of variable genes are not developed in this figure, for simplicity. Environmental (E) and ontogenetic (O) effects drive the values of the traits and links. All links do not have the same strength (line width).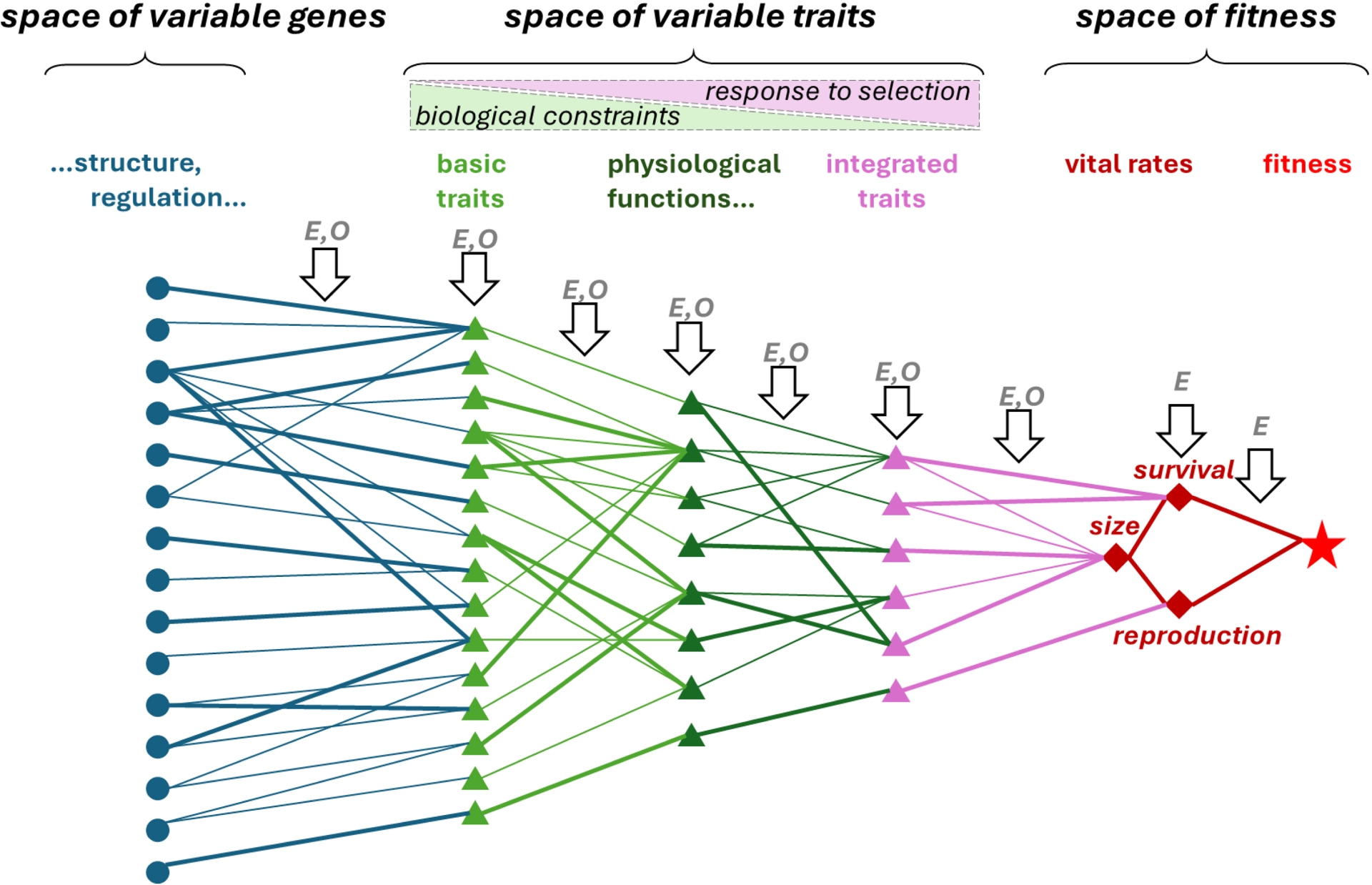
In the fitness space, survival and reproductive success are the ultimate fitness components represented in Figure 2, which can themselves build upon stage-specific vital rates. For trees, tree size is a particular trait with a pivotal role on fitness that can be put in the fitness space (the frontier between fitness components and variable traits is thin). Indeed, in normal conditions, bigger trees have a higher probability of surviving to competition (self-thinning) and also tend to reproduce more, due to higher gamete production and higher dispersal capacity. Inversely, under some stresses or disturbances, bigger trees may be disadvantaged. In addition, tree size makes links between many other traits and fitness, while few traits are directly linked to survival or reproduction without affecting tree size. In a sense, the pivotal role of tree size on fitness is analogous to the role of body size in animals (Araya-Ajoy et al., 2019). Still, some traits, typically those related to defense and resistance mechanisms, may directly act on survival or reproduction without any effect on tree size, or with an antagonistic effect on growth vs. survival or reproduction in case of resource allocation trade-offs (Shipley et al., 2006).
The links between the traits and the fitness components are responsible for the selection gradients. The space of traits can be split into several hierarchical sublevels with different links to fitness (Mousseau and Roff, 1987). In Figure 2, three sublevels of traits are schematized, but the hierarchy might be more complex, or simpler, depending on the considered set of traits. The integrated traits are the traits most directly linked to the fitness components, they build upon functional traits that have more indirect and complex links to fitness and, at the bottom of the hierarchy, the links of basic traits to fitness depend on the whole chain of links across sublevels. To illustrate the concept in forest trees, vigor—defined as potential growth without stress—, growth sensitivity to drought or frost tolerance are three typical examples of integrated traits affecting tree size and sometimes directly affecting survival. Each of these integrated traits is potentially determined by several functional traits, such as leaf phenology, which are themselves driven by multiple basic variable traits, such as chilling and forcing requirements in the case of phenology. The most basic sublevel would include the variable parameters of physiological functions, metabolic pathways, or some variable traits at fine biological level, e.g. at the cellular or tissue level, involved in multiple functions. The genes–traits–fitness map should be seen as a dynamic connection network: trait values and links are dynamic, they vary with the environment and developmental stage, some links are more variable than others.
The space of genes controlling the traits could also be split into several sublevels to represent gene expression and regulation networks, not represented in this figure for simplicity and because they are out of the scope of this review. The links between genes and traits illustrate cases of multiple genes linked to the same trait, which can be purely additive or include epistatic interactions, as well as complex pathways leading to the pleiotropic effect of a single gene on multiple traits. Note that equivalent links also exist between different sublevels of traits, as well as between traits and fitness: the concepts of epistasis, i.e. interactions among several genes on a single trait, and pleiotropy, i.e. the correlated effect of a single gene on several traits, could be extended to the effects of traits on more integrated traits and to the effects of traits on fitness.
Natural selection directly acts on fitness, with indirect effects on traits and genes that depend on the network of links connecting them to fitness. Because the links vary in space and time, selection is a variable and dynamic process responsible for and impacted by phenotypic and genetic evolution in the population.
The quantitative genetics theory tells us that, when a polygenic trait is under selection, the first response in the space of genes consists in the emergence of covariations between quantitative trait loci (QTL) alleles before changes in individual allele frequencies (Kremer and Le Corre, 2012; Latta, 1998). The contribution of covariance between QTL to the response to selection starts with two QTL controlling the trait; it increases with the number of QTL involved and with the intensity of selection (Kremer and Le Corre, 2012). Change in QTL allele frequencies is a rather long-term process requiring some stability of selection on the QTL over the evolutionary time scale, even though rapid evolution of allelic frequencies may occur under strong selection pressure, such as severe epidemics (Metheringham et al., 2025). Extending these well-known results of quantitative genetics, we expect a similar mechanism to apply throughout the genes–traits–fitness map where fitness builds upon multiple traits through a complex and dynamic connection network. Thus, we expect that covariations of traits respond rapidly to selection and that integrated traits are more impacted by selection than basic traits, because integrated traits have more direct and consequently more stable links to the fitness components regarding environmental and ontogenetic effects.
Another expectation is that biological or development constraints are more often driving the variation of basic traits than integrated traits, whereas selection is a more important driver of variation for integrated traits than basic traits (as illustrated in Figure 2). Indeed, some basic morphological traits at cellular or tissue levels or metabolic pathways are so vital that their range of non-lethal variation is expected to be globally constrained, within and between populations, and this constrained variation limits their response to selection. By contrast, integrated traits build upon multiple alternative functional pathways; their variation is expected to be less constrained due to functional redundancy among different combinations of basic or functional traits producing a similar integrated syndrome. Less constrained, these integrated traits are also tightly linked to fitness and more directly impacted by selection, resulting in differentiation between populations and sometimes locally, but not globally, low levels of variation within populations under selection. Let us examine these different expectations on trees.
3. Trait variation and covariation patterns in trees: a matter of biological constraint or response to selection?
The patterns of phenotypic variation observed at different levels of organization result from the combination of multiple factors: biological or developmental constraints, phylogenetic legacy from an ancestor species or population, plasticity, and response to selection. Similarly, the covariation patterns among traits result from biological or developmental constraints, phylogenetic heritage, correlated plasticity and correlated response to selection, which are differently expressed at interspecific or intraspecific levels, within and between populations. For trees, common garden experiments are classically used to avoid confounding plasticity and response to selection in the analysis of trait variation and covariation patterns. Some methods are also used to disentangle these effects, or at least to achieve a conservative test of a response to selection, using in situ observations in the actual environmental conditions (Acuña-Míguez et al., 2026; Halliwell et al., 2025; Sanchez-Martinez et al., 2024).
At the interspecific level, Maherali et al. (2004) reviewed the literature on two functional traits that contribute to drought resistance, resistance to xylem cavitation and water transport capacity, among 167 woody plant species. They found differences in median values between vegetation types adapted to different drought levels. However, there was still huge variation among species within each vegetation type, with considerable overlap across the different vegetation types. This observation is consistent with the idea that any single functional trait is not the unique driver of adaptation: adaptation is primarily a matter of syndromes of multiple traits. In addition, Maherali et al. (ibid.) found a strong effect of the phylogenetic legacy on the within-species variation of resistance to cavitation as well as on the within-species correlation between the two studied traits, rather than evidence of convergent adaptive responses to selection. This is consistent with the idea that biological or developmental constraints on the variation and covariation of basic or functional traits somehow prevent them from big changes along species phylogenies. Similarly, in a phylogenetic study of life history and adaptive traits among Mediterranean pine species, Grivet et al. (2013) detected almost no adaptive convergence of single traits across taxa. However, they found three trait correlations that they interpreted as alternative genetic adaptation strategies to fire: two negative correlations, between seed dispersal and bark thickness, as well as between serotiny and maximum life span, illustrating a classical contrast between r and K-strategies; and a positive correlation between serotiny and seed dispersal, illustrating a variation of efficiency in the r-strategy.
At intraspecific level, provenance trial experiments in trees usually reveal clinal genetic differentiation patterns for growth-related traits among populations sampled along environmental gradients, thus indicating the high evolvability and effective response to selection of such integrated traits (Savolainen, Pyhäjärvi, et al., 2007, for review). We also have evidence of the response of growth-related integrated traits and phenology to selection occurring at microgeographic scale within gene dispersal distance (Scotti, Gonzalez-Martinez, et al., 2016, for review). Evidence of temporal genetic changes of integrated traits in response to selection across successive generations of trees is still scarce, but cases of rapid genetic adaptation have been observed in populations introduced in a new environment (Fallour-Rubio et al., 2009; Peterken, 2001; Skrøppa et al., 2010). At the bottom of the hierarchical space of traits, globally limited variation of resistance to cavitation was shown across maritime pine populations over the species’ range, suggesting biological or developmental constraints (Lamy et al., 2014).
Generally, the highest values of genetic differentiation among tree populations are observed for growth, phenology and frost hardiness, whereas more basic traits linked to leaf morphology and chemistry, wood anatomy, hydraulic properties or photosynthetic capacity show much lower differentiation (Alberto et al., 2013; Hajek et al., 2016). A similar pattern was also found in the case of selection at microgeographic scale: that is, a higher value of differentiation for growth-related traits and phenology than for leaf morphology and chemistry or water use efficiency (Gaüzere et al., 2020). These results are consistent with the expectation that basic traits should be less impacted by selection than integrated traits due to more distant and flexible links to fitness for basic traits. Interestingly, in these studies of local adaptation in different environments, the variation patterns of phenological traits are highly impacted by recent selection (in the time scale of local adaptation), as much as or even more than growth-related traits: this is due to the potentially strong impact of phenology on all fitness components across different environments. In a rare study assessing selection gradients within a single population for different types of traits in two sympatric oak species, Alexandre et al. (2020) found significant selection gradients for growth traits and defense traits in both species, for leaf morphology only in one of the species, but not for phenology in either species: phenology was not tightly linked to fitness in this particular environment. By contrast, evidence of spatial variation in selection gradients on phenology was found along an environmental gradient in beech, showing that phenology was tightly linked to fitness along this gradient (Oddou-Muratorio, Bontemps, et al., 2024).
Cases of GxE interaction are frequently observed in forest trees, making plasticity itself a variable trait. Alía, Climent, et al. (2024) used a provenance/progeny test in two contrasted environments to assess the evolvability of different traits in 11 maritime pine populations: the plasticity of water-use efficiency (WUE) showed the highest evolvability, even higher than growth, while mean WUE showed the lowest evolvability. Using a provenance/clone experiment in two contrasted environments with the same species, Ramírez-Valiente, González-Martínez, et al. (2025) showed that populations adapted to a mild climate had higher growth plasticity than populations adapted to arid conditions. Analyzing dendrochronological data between and within successive generations within an introduced cedar population initially planted in a dry eroded land, Fallour-Rubio et al. (2009) detected a significant change in the growth plastic response to summer drought across generations, but no generational change in mean growth. In this case, growth plasticity increased from the first to the third generation, while drought stress progressively decreased due to soil restoration under the development of the new forest ecosystem. All these results show that plasticity traits can be highly responsive to selection, at least as much as integrated traits and more than basic traits. However, this is not always the case: in a common garden experiment of oak populations sampled along an altitudinal gradient, Soularue et al. (2023) observed a genetic cline on budburst date but not on its plastic response to temperature.
Trait correlations also vary at intraspecific levels and, therefore, potentially respond to selection (Climent et al., 2024, for review). Intraspecific trait correlations between and within populations are generally different (Acuña-Míguez et al., 2026; Alía, Climent, et al., 2024; Prada et al., 2016). Between-population trait coordination potentially indicates a correlated response to selection, i.e. the emergence of adaptive trait syndromes as bases of intraspecific ecotypes (Blasini et al., 2021). Within-population trait coordination can also result from a correlated response to selection within populations, when several trait syndromes increase fitness as observed in a beech population at the southern margin of the species distribution range (Bontemps et al., 2017). Trade-offs between growth vigor in favorable conditions and stress resistance traits are commonly observed at both scales, between populations (Rehfeldt et al., 2001) and within populations (Fririon, Davi, Oddou-Muratorio, Lebourgeois, et al., 2023). Note that having a similar trade-off within and between populations is necessary but not sufficient to provide evidence of a biological “cost” of resistance such as a resource allocation balance. For instance, at the between-population level, Modrzyński and Eriksson (2002) observed in a phytotron experiment that Norway spruce provenances from high elevations were more tolerant to drought, but grew less in good conditions, than provenances from low elevation. Rather than representing a “cost of drought tolerance”, this difference could be explained by a difference in phenology, because in this species high elevation provenances start growing earlier in the common environment and develop deeper root systems before the drought occurs. At within-population level, adaptive trade-offs also simply result mathematically from some types of correlated selection, such as threshold selection on a linear combination of multiple traits, i.e. index selection (Figure 3). For trees in particular, trade-offs emerging from selection are expected between traits that jointly contribute to tree size. Indeed, tree size can be seen as a selection index being the sum of annual growths in variable conditions, which mathematically results in a trade-off after selection between the different traits that contribute to increase tree size depending on different annual conditions, as classically observed between vigor and stress tolerance traits (Figure 3). More generally, when fitness is expressed as a weighted sum of multiple traits, selection is expected to generate trade-offs among these traits, depending on their relative contributions to fitness. Thus, considering that trait contributions to fitness are represented through the functional links on the genes–traits–fitness map, this representation could also serve to predict trade-offs among traits that could emerge from selection.
Phenotypic trade-offs emerging from index selection. (a) X1 and X2 are two independent traits, in a standardized scale, contributing positively to a weighted selection index with relative contributions 0.6 and 0.4, respectively: index = 0.6X1 + 0.4X2. The rate of selection on the index is 0.6 (the 60% lowest values are eliminated). In grey are the values before selection, the white “+” is the initial population mean before selection. In red are the values of selected individuals, the green “+” is the mean after selection. With these values, selection generates a negative phenotypic correlation (no reproduction considered here) −0.49 within the selected population. Considering the situations before and after selection as two populations, note the opposite (positive) sign of the between-population correlation caused by the increase of both traits means in the selected population. (b) Values of the correlation (trade-off) within the selected population as a function of (i) the weighting of the traits in the index and (ii) the rate of selection. Note the important effect of the weighting. For instance, tree size is the sum of annual growths in good years (related to vigor) and annual growths in stressful years (related to stress tolerance): selection on tree size creates a trade-off between vigor and stress tolerance, and the value of the negative correlation depends on the frequency and intensity of stress events that determine the relative contributions of both traits to the index.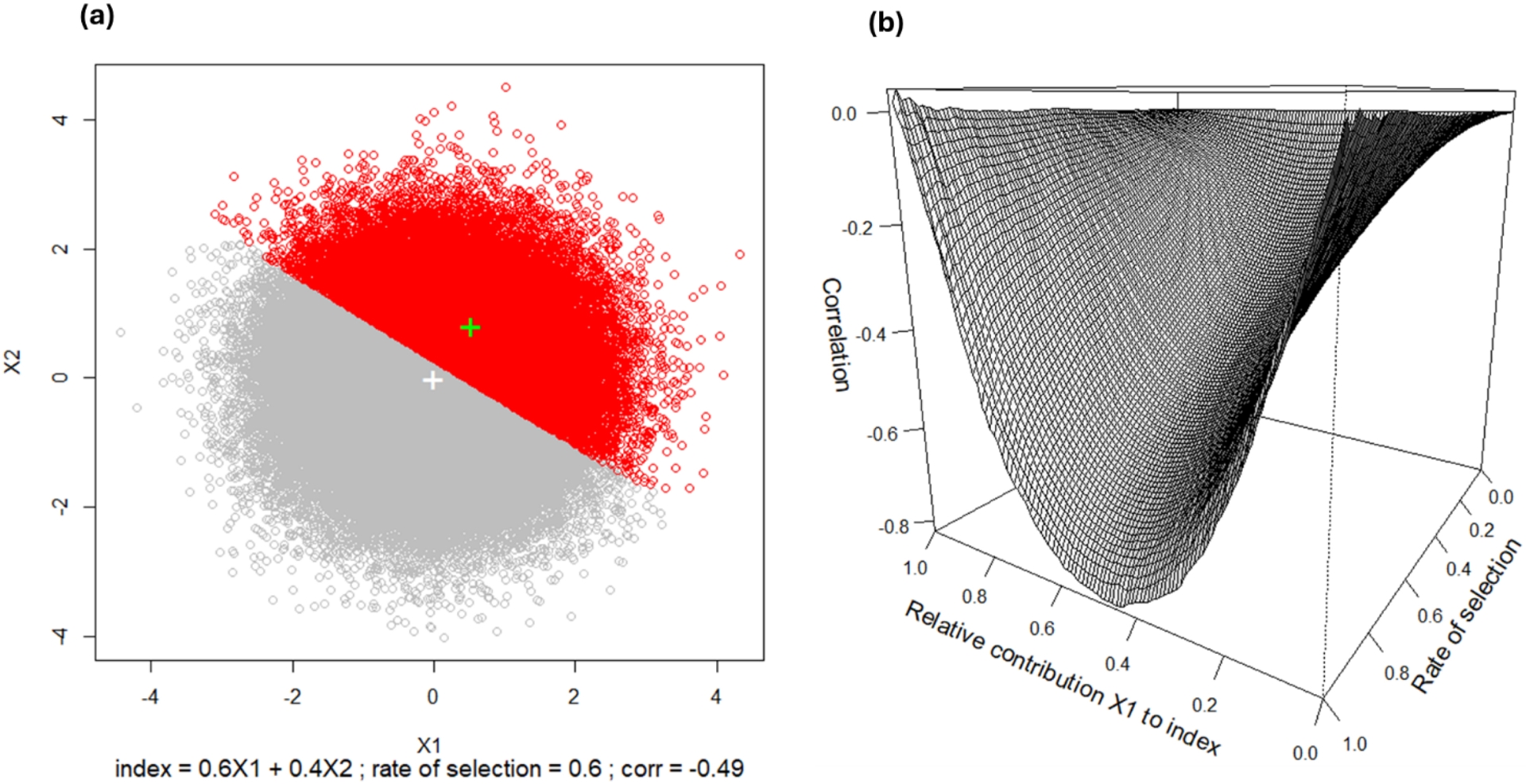
4. Spatiotemporal variability of selection in trees
Selection pressures, and therefore the traits under selection, vary in time and space within tree populations. Within a forest, the external causes of tree mortality change from year to year driven by the annual climatic fluctuations and other disturbances (Petit-Cailleux et al., 2021). Thus, living trees in the forest can be seen as survivors to multifactorial and stepwise selection: e.g., a rare severe frost event occurring in an arid site will select the frost tolerant among drought tolerant trees, and the genetic impact of the selection event on frost tolerance may last over several generations. Selection pressures on trees also depend on the environment (Alía, Chambel, et al., 2014), and high within-population micro-environmental heterogeneity results in the interference between multiple selection processes spatially distributed within gene dispersal distance (Brousseau et al., 2015; Gaüzere et al., 2020).
In a stable environment, the forest stand dynamics itself act as an internal driver of spatial and temporal variability of selection, for two reasons. Firstly, trees’ adaptive capacities and their relations to fitness vary across developmental stages (Alía, Chambel, et al., 2014; Pardos et al., 2014; Peltier and Ogle, 2020), making selection gradients variable throughout individual lifespan. The consequences of such developmental effects on the spatiotemporal variability of selection might differ between even-aged forests, with synchronous developmental stages, and uneven-aged forests, where various developmental stages coexist, forming an irregular stand structure. Secondly, stand growth and demography dynamics generate temporal changes in the intensity of density-dependent selection and other soft selection mechanisms (Godineau et al., 2023). They also induce temporal changes in the level of stresses like drought that increases with stand leaf area (Breda et al., 1995) or in the intensity of pests and parasites pressures that vary positively or negatively with host density (Knight et al., 2013; Sholes, 2008).
In the context of climate change, the temporal horizon of climatic scenarios matches the lifespan of trees. Disturbances and uncertainties are key drivers of selection on this time scale. Climate change projections consist in two phases: a first phase of predicted climatic transition with annual fluctuations and disturbance regime shifts in the next 30 to 50 years, followed by a second phase with increased uncertainties due to divergent model predictions (IPCC, 2021). Most of the current forest trees will experience the first phase of change in selection pressures, possibly survive until the more uncertain second phase, and some of them might even survive beyond the longest climate projections. In other words, shifts in selection processes are expected at the same velocity or even faster than the response of tree populations to selection, and the spatiotemporal variation of selection processes is highly uncertain.
Beyond the effects of the mean climatic trend on the dominant selection pressures, such as the global increase of drought and heat stress impacts on forests (Allen et al., 2010), disturbance regimes on European forests have already shifted towards higher frequency and intensity since 1950 (Patacca et al., 2023). Thom et al. (2017) showed that small scale hazards, when surviving trees remain at a distance reachable by gene flow, are expected to exert a selection at population or metapopulation levels and simultaneously offer opportunities for the establishment of selected progenies, which can ultimately foster adaptation, while large-scale disturbances are expected to impede forest adaptation. Peltier and Ogle (2019) detected a detrimental impact of repeated droughts on growth decline in ponderosa pine, with regional differences of sensitivity among populations. Seidl and Turner (2022) used the adaptive cycle framework to explore post-disturbance reorganization of forest ecosystems, depending on the changes in the structure and composition of the ecosystem: this approach could probably be extended to the reorganization of phenotypic trait variation and genetic diversity.
5. Impacts of forestry interventions on trait variation and selection
Intentionally or not, silviculture and forest planning interventions are expected to drive trait variation, not only through the control of the genetic composition of plantation forests but also through multiple impacts on selection in naturally regenerated forests: (i) by inducing or modifying genetic drift and gene flow that interfere with selection; (ii) by changing the environmental conditions that determine the type and intensity of natural selection; (iii) by generating additional anthropogenic selection (Aitken and Whitlock, 2013; Lefèvre, 2004; Lefèvre et al., 2014; Savolainen and Kärkkäinen, 1992). Currently, the quantitative assessment of these potential impacts mostly relies on modeling work, as empirical assessment is just emerging based on ex-post reanalysis of long-term monitoring experiments and new experimental designs.
Simulation studies on different types of forests show that the large population size and important gene flow capacities of trees generally prevent significant genetic drift due to management. In the case of large populations, as commonly observed for temperate tree species, classical silvicultural strategies are not expected to reduce the number of reproducing trees as much as required to induce genetic drift (Godineau et al., 2023). Even in the case of highly diverse tropical forests with low population density for each species (a few adult trees per hectare), Degen et al. (2006) showed that the frequency and intensity of selective logging have an impact on tree growth and population demography, without a significant reduction of gene diversity or an increase in inbreeding.
Intentional assisted gene flow is an option to rescue populations that suffer demographic and genetic decline under strong selection. The expected benefits include demographic support with pre-adapted genotypes, genetic enrichment with new alleles of interest and reduction of inbreeding depression, while the main associated risks are outbreeding depression and introduction of undesired alleles (Aitken and Whitlock, 2013, for review). In particular, this option is envisaged to foster forest adaptation to climate change (Browne et al., 2019; Devresse et al., 2025). Practical experiments are still rare and recent, lacking long term monitoring of potential risks associated to the introduction of long distance material that could bring in undesired heritable characteristics linked to the target traits of interest (Young et al., 2020). When assisted gene flow would consist in introducing distant genetic material into a small local population, a strategy to reduce the risks of maladaptive genetic swamping and of further drop in effective population size is to use a highly genetically diverse donor gene pool (Lefèvre, 2004). Assisted gene flow is also envisaged in the case of emerging pests and diseases like the ash dieback in Europe (Semizer-Cuming et al., 2021). In this case, we expect lower risk of maladaptation or outbreeding depression when the introduced resistant genotypes come from the surroundings of the declining stand.
Kramer et al. (2008) made a distinction between three categories of traits with regard to natural and anthropogenic selection: (i) traits under natural selection with little consideration in management, such as flowering phenology when it does not induce major defaults; (ii) traits selected in forestry with little adaptive consequences, such as spiral grain; and (iii) traits under both natural and anthropogenic selection, such as growth-related traits and resistance traits. The genes–traits–fitness map could help identify complex links between natural and anthropogenic selection through their respective target traits. All traits that are somehow linked to tree size fall in the category of traits submitted both to natural and anthropogenic selection. For these traits, management interventions interfere with natural selection through different mechanisms. Understanding these mechanisms can help to solve potential trade-offs between short-term objectives, such as reducing competition or stress levels, and long-term objectives, such as fostering genetic adaptation.
In the short term, increasing morphological or functional intraspecific trait variation by planting genetic mixtures can increase forest productivity and resilience due to positive mixture effects, e.g. reduced competition for resources, and phenotypic adjustment. In a 40-year provenance mixture experiment of Norway spruce, Pretzsch (2021) showed that increased provenance diversity around trees reduces competition and increases tree growth and stand growth in the same magnitude as in species mixtures (+28%). Similarly, at more juvenile stages, the reduction of competition and increased tree growth in clonal mixtures compared to monoclonal stands in 9-year-old loblolly pine experiments (+4–5%) was attributed to morphological trait variation (Carter et al., 2025). Thus, genetic mixing induces positive mixture effects and phenotypic adjustment. However, the ultimate consequences on genetic evolution are context dependent, because of antagonistic impacts on the response to selection. Indeed, genetic mixing increases the variance of the trait under selection on the one hand, but reduces the intensity of one potential mechanism of selection, competition, on the other hand, and the balance between these antagonistic effects is contingent to the local conditions.
Favoring tree species mixtures is an option to increase forest resilience. In such a situation, the interference of species diversity with the potential evolution of each species is questionable. Using a demo-genetic modeling approach in a climate change scenario, Devresse et al. (2025) showed that beech and fir are not expected to have slower genetic evolution in mixture than in monospecific stands, because both species evolve jointly and the interspecific mixture does not induce sufficient genetic drift to hamper the response to selection. However, the introduction of other pre-adapted species may over-compete the local species and preclude their evolution.
Thinning regimes, i.e. frequency, intensity and type of thinning (selective from below, from above or not selective), are generally aimed at reducing competition. They have multiple impacts on selection. Demo-genetic models coupling forest dynamics and genetics are useful for disentangling these impacts. If they were strictly non-selective, some intensive thinning regimes currently used in forestry could drastically reduce natural density-dependent selection mechanisms, i.e. substitute selective self-thinning with non-selective thinning, resulting in a drastic reduction of the evolutionary rate of growth-related traits across generations (−84%) (Godineau et al., 2023). However, in practice, thinning is often selective from below, in which case the forester selects in the same direction as natural selection does, in favor of trait values that promote tree size, and the substitution of natural selection with anthropogenic selection could even result in increased adaptive evolutionary rate (+30%) compared to no management (Fririon, Davi, Oddou-Muratorio, Ligot, et al., 2024). Note that selective thinning partly from above, sometimes used to obtain an economic revenue to pay for the intervention, always opposes natural selection with possible long-lasting consequences on the maintenance of maladapted genotypes in the forest.
Under drought stress conditions, density reduction is also used to alleviate stress, providing a short-term functional benefit for the current tree generation. In this case, the short-term benefit is associated with reduced selection intensity for drought tolerance, due to reduced drought mortality (hard selection mechanism) and reduced contribution of drought tolerance to tree size (soft selection mechanism through competition and self-thinning), slowing down genetic adaptation. A strategy to avoid this short vs. long-term trade-off—reducing stress in the short term hampers genetic adaptation in the long-term—consists in leaving more room for natural selection in the juvenile stages and reducing stress in later stages (ibid.). Thinning regimes in harvested stands can also result in enhanced growth of the remaining trees. This leads to shorter rotation between successive harvests and, therefore, shorter generation time (Kramer et al., 2008), which interfere with the velocity of genetic evolution.
6. Conclusion for research and forest management
6.1. Extended views on the genes–traits–fitness mapping conceptual framework
Here, I used the genes–traits–fitness mapping conceptual framework, focusing on the spaces of variable traits and fitness, to explore the joint dynamics of trait variation and selection in trees. The schematic representation on Figure 2 only shows part of the picture; other aspects that are not represented in this figure need to be considered. Regarding the space of traits, the multiple effects of the environment appear independently on this representation, which does not account for coordinated environmental effects on multiple traits and links. Missing coordinated environmental effects does not facilitate the exploration of the role of plasticity in the joint dynamics of trait variation and selection (Chevin, Collins, et al., 2013; Chevin and Chauhan, 2025), even though we have empirical evidence that plasticity traits are tightly linked to fitness. Obviously, the extension of the representation of the space of variable genes, through an analog dynamic connection network with links between environmental and ontogenetic effects on gene expressions and pathways, would deserve exploration.
Another aspect hardly represented on a diagram is the spatiotemporal variability of selection mechanisms: their integration through time and space still remains to be analyzed. For instance, in even-aged forests, multiple stepwise selection mechanisms globally result in a composite selection process from one generation to the next. The integrated composite selection relates to the genes and traits spaces through an integrated genes–traits–fitness map. Obtaining this integrated map, some genes and traits may appear to be tightly linked to the composite fitness for two reasons: either they are tightly linked to one important selection mechanism in particular, or they have consistent and robust links with multiple selection mechanisms. These two situations are not mutually exclusive. In the second case, we might even imagine some genes or traits poorly linked to each selection mechanism, but consistently linked to all: these genes or traits would be strongly involved in and impacted by the response to composite selection. Looking at between-population adaptation, Yeaman et al. (2016) detected cases of convergent selection on specific gene alleles between two distantly related species, lodgepole pine and interior spruce. At within-population scale, Scotti, Lalagüe, et al. (2023) found genetic signature of microgeographic selection in different tree species, without evidence of convergent selection across species so far. These results suggest the possibility of strong links between genes and fitness, and particularly robust links in case of convergent selection in different species. In plants, some types of genomic elements playing a key role in phenotypic variation, notably some transcription factors (Engelhorn et al., 2025), could be good candidates for genes having an important role in composite selection processes.
Nevertheless, although incomplete, the present review already provides indications for applied management.
6.2. A dynamic vision of adaptation in trees
Trees are submitted to temporally variable, successive, selection pressures during their lifespan. Tree populations cover large areas due to high dispersal capacities, and microgeographic environmental heterogeneities make selection spatially variable within populations. Selection pressures, selection intensities and selection gradients (the links of traits to fitness) vary in space and time, even at fine scales, driven by the local environmental conditions, which consist of external biotic and abiotic factors, combined with internal factors such as population structure or developmental stages. Therefore, local adaptation in trees should be seen as a dynamic process of response to multiple selection pressures with spatiotemporal dependencies, rather than an achieved long-term response to a single selection driven by mean local conditions. Within-population trait (co)variation is simultaneously a constraint for and the result of selection; it continuously varies within and across generations of trees. Considering the spatiotemporal variability of selection helps us to better understand the causes and consequences of trait variation patterns. The interdependent dynamics of trait (co)variation and selection are impacted by developmental factors, population dynamics, external environmental factors, and forest management interventions.
Selection is expected to have more impact on integrated traits and trait syndromes than on the variation of functional and basic traits. This flexibility across the traits–fitness map allows for rapid response to selection without severe erosion of the variation of functional and basic traits, which are the basic bricks of the capacity of response to variable selection pressures. An illustration of this flexibility was given above with the southern beech population adapted to drought through multiple trait syndromes conferring drought resistance while maintaining the variation of each trait involved (Bontemps et al., 2017).
Forest management interventions have multiple impacts on the spatiotemporal variability of selection, which can ultimately result in reinforcement or impairment of natural selection, and eventually create additional anthropogenic selection. Understanding these impacts is strategic for the design of management options, whether it be for forestry or conservation purposes, particularly in the context of environmental changes and uncertainties. In this context, natural selection should be considered as a lever of action in forest management and forest genetic resource conservation, as illustrated below in two situations. This is not an exhaustive list of situations where evolution-oriented forestry would be beneficial (Lefèvre et al., 2014).
6.3. Forest management with natural regeneration under climate change
In naturally regenerated production forests facing new stresses, e.g. climate change, there is a need to find a compromise between a short-term objective to enhance the production of the current stand under pressure and a longer-term objective to foster genetic improvement for stress tolerance in the next generation. Natural selection can definitely help to find such a compromise. Simulations of a regular Douglas fir stand under drought stress provided a first estimate of the expected genetic gain in productivity that can be obtained through natural selection on vigor and drought tolerance: +13% gain per generation on mean stem diameter at the final cut in the simulated context (Fririon, Davi, Oddou-Muratorio, Ligot, et al., 2024), which is in the same order of magnitude as the average gain expected through breeding (Jansson et al., 2017). A multi-objective strategy that enhances short-term productivity and fosters long-term adaptation can be designed through a thinning regime that allows for natural selection to operate in the juvenile stage before reducing stress, and thus reducing natural selection, at a later stage (Fririon, Davi, Oddou-Muratorio, Ligot, et al., 2024). To speed up adaptation, it is also proposed to use more dynamic silviculture, i.e. to reduce the rotation time and, therefore, accelerate generation turnover. This action on the denominator of the “gain per generation” assumes that most of the natural selection operates at the juvenile stage, so that reducing the generation time does not significantly decrease the total gain (the numerator). However, this assumption may be questionable. Firstly, reducing generation time may result in enhanced selection for vigor and reduced selection for stress resistance, due to fewer or less intensive stress events and to the trade-off between vigor and resistance traits, which might, at the end result, in faster but lower adaptation. This is expected to happen in forests currently under moderate and rare stress, whereas the initial assumption that sufficient selection for stress tolerance is achieved in shorter time remains valid in forests under heavy stress. Secondly, considering the huge uncertainty about future selection pressures and adaptation needs, reducing the duration of exposure to natural selection also reduces the chance of multiple selection criteria to combine within each generation. This results in sequential multi-trait selection, i.e. sparing different selections across different generations, rather than multi-trait index selection within each generation. Will one selection process be more efficient than the other at the end? In any case, multi-trait index selection offers more insurance through time. Considering these different aspects together, reducing generation time may not be much effective to accelerate the adaptation of forests that are currently under moderate stress and will suffer a heavy shift in the stress regime; it will be more effective and efficient in forests already under strong selection. The effects of thinning on the selection intensity for traits of interest, on the one hand, and on the generation time, on the other hand, ultimately combine in higher or lower evolutionary rates per generation. A compromise to combine short-term stress reduction and faster evolution towards stress tolerance can be reached by adjusting the dates, intensities and types of successive thinning, taking into account the local context of natural selection.
6.4. Dynamic conservation of forest genetic resources
Regarding the conservation of forest genetic resources, priority is given to dynamic in situ strategies whenever possible, allowing the conserved populations to evolve in their original environment (EUFORGEN, 2021). Relocation is sometimes envisaged for some marginal populations that have developed specific adaptive characteristics of interest but considered locally at risk due to severe selection pressure. The so-called dynamic ex situ approach consists in moving these marginal populations of interest to a safer place and letting them dynamically evolve there. However, it should be clear from this review that conserving a specific adaptation in an evolving population cannot be achieved without conserving the selection mechanisms that create and maintain this adaptation. If the new site where the population is transferred is less selective, the next generation of trees will lose part (possibly a large part) of their ancestral adaptive specificities and rapidly evolve towards new characteristics fitting the new environment, as previously illustrated with the cases of rapid adaptation of introduced populations (Section 2). Therefore, if the conservation objective is to preserve the original adaptive specificities, dynamic ex situ conservation is not the most effective strategy. In a dynamic approach, not only genetic diversity but also selection itself should become a target of conservation. Before taking a decision of relocation, the risk of local extinction should not be overestimated. Having a reasonable amount of dieback is a rather positive sign of ongoing adaptation, given that the threshold of population viability is not passed, of course. An alternative option overlooked for these extreme populations developing specific adaptive characteristics under severe stress could be to ensure sufficient variation to feed selection and rescue the population at risk where it is, i.e. using genetic mixing strategies like provenancing or evolutionary rescue (Hoffmann et al., 2021). This would be a type of assisted gene flow strategy, not introducing pre-adapted material (that may not be available for such marginal situations) but introducing the least-maladapted material with the highest genetic diversity possible, as a demographic supply, with the hope that enough offsprings will reach the level of adaptation required. For species lacking such marginal populations of interest in conservation programs, installing highly diverse genetic mixtures in stressful marginal areas surrounding the species range would not necessarily be an absurd option to consider and assess, aiming to achieve conservation with genetic improvement by natural selection. This would be a dynamic ex situ approach in which genetic mixtures would be installed not in sites with less selection but in sites with specific selection of interest, despite the evident associated risk for management and probable additional costs. This would mean looking for the effectiveness of the conservation rather than just its efficiency, i.e. emphasizing the level of adaptation potentially conserved despite the cost rather than emphasizing the chance of conservation success with a reduction of adaptive capacity (which is another type of cost).
6.5. Back to the original definition of a resilient approach in management of natural resources
In the context of climate change, forest managers classically identify some specific predictable adaptation goals, such as climate adaptation. A further goal is to integrate the evolvability, or adaptive potential, as an additional objective to cope with unpredictable future needs. In addition to these goals, integrating the variability of selection into forest management thinking, and considering the selection process as an objective per se, would further help to develop a flexible adaptive strategy, taking the benefit of natural selection processes rather than counteracting them, to drive the forests towards multiple adaptation requirements. This environmental stewardship approach in management (Mathevet et al., 2018) would also be a resilient approach, as defined by (Holling, 1973):
The stability view emphasizes the equilibrium, the maintenance of a predictable world, and the harvesting of nature’s excess production with as little fluctuation as possible. (…) A management approach based on resilience, on the other hand, would emphasize the need to keep options open, the need to view events in a regional rather than a local context, and the need to emphasize heterogeneity. Flowing from this would be not the presumption of sufficient knowledge, but the recognition of our ignorance; not the assumption that future events are expected, but that they will be unexpected. The resilience framework can accommodate this shift of perspective, for it does not require a precise capacity to predict the future, but only a qualitative capacity to devise systems that can absorb and accommodate future events in whatever unexpected form they may take.
Disclaimer
Intentionally, the author did not use AI for this review, neither for the literature search and analysis nor for the writing and illustrations, leaving perfectibility as an opportunity for the reader to develop her/his own improved personal thoughts on the topic.
Déclaration d’intérêts
The author does not work for, advise, own shares in, or receive funds from any organization that could benefit from this article, and has declared no affiliations other than their research organization.