

Abridged version
For all the marine palaeontological groups, the Permian–Triassic crisis is one of the most dramatic mass-extinction in the Phanerozoic. At the end of the Permian, 51% of the marine families disappear (82% of the genera and 96% of the species) [12,28].
The ostracods, like the other organisms, are subject to this conjunction of noxious events (Fig. 1).

Schéma résumant les événements de la limite Permien–Trias (d'après [12,20], modifié).
Summary of Permian–Triassic events (after [12,20], modified).
The Early Triassic ostracods are very poorly known. Some species are recognised in the Early Triassic (Induan–Olenekian) of the Precaspian Depression [18,26,27], in Nepal [16], in Kashmir [1–3], in the Germanic Basin [17], in Greece [5], in Israel [15], in southern China [14,30,31] and in Pakistan [29].
The Çürük dağ section (N36°41′32–E30°27′40) is located in Western Taurus, in Antalya Nappes (Turkey) (Fig. 2) [23,25]. In the Kemer area, the Pamucak Formation (Middle–Late Permian) is overlain by the Kokarkuyu Formation (Early Triassic) [4,22,24]. In the Çürük dağ section (Fig. 3), the Pamucak Formation presents a thick (400 to 600 m) succession of inner to outer platform facies (Capitanian to Changsingian). The upper part of the formation, detailed here, is made up of black nodular limestones locally with chert. They are rich in calcareous algae (Dasycladacea) and in small foraminifera (mainly Milliolidae), with levels containing brachiopods, echinoderms and crinoids [22,24] The microfacies consists of bioclastic wackstones of low energy (algae biomicrites [4]). Towards level 5 (Fig. 3), the microfacies changes into high-energy grainstones, then into oolitic grainstones (level 8), with echinoderms and lamellibranches. The top of the Pamucak Formation shows evidence of the proximity of subaerial surface. The Kokarkuyu begins with 12 m of microbial limestones (levels 9 to 14) followed by 3 m of calcilutite (level 15). The first Triassic levels are rich in Induan foraminifera (Ammodiscus, Rectocornuspira, Cyclogira, Earlandia) and rare in conodonts (Isarcicella isarcica staeschei (Dai & Zhang) – sample T11 – and Hindeodus parvus Kozur & Pjatakova – sample T13; Figs. 4.12–4.15), whose association is an index of the second Triassic biozone.
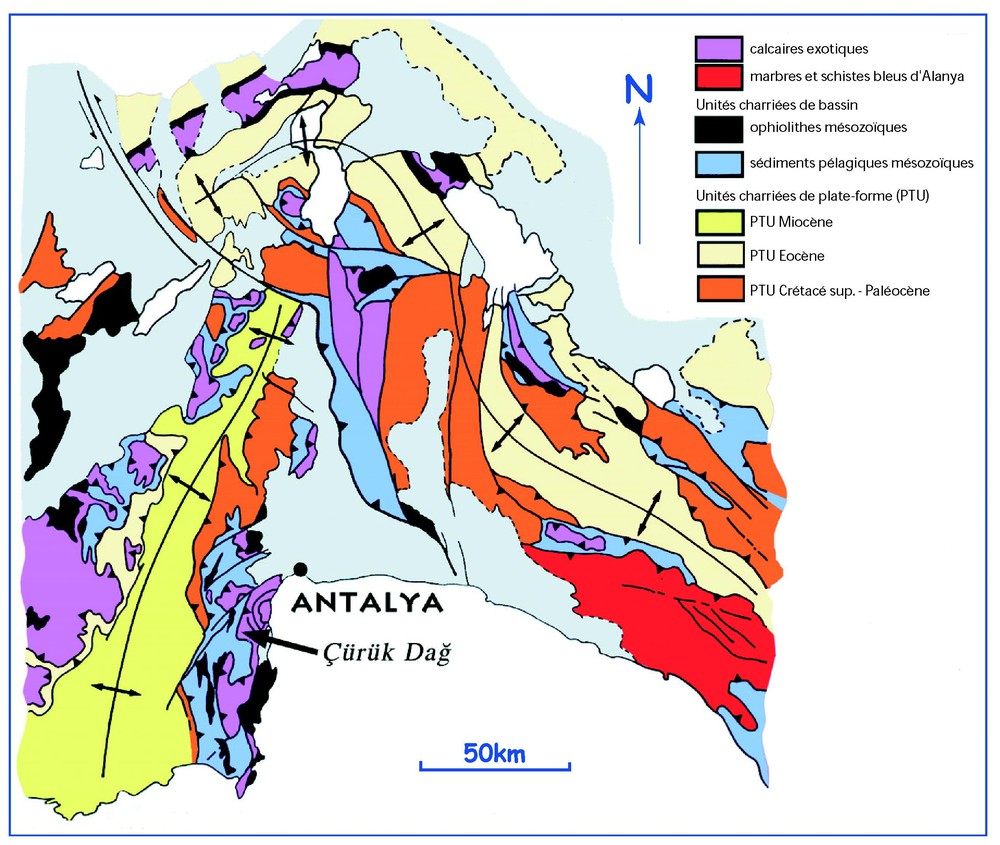
Localisation de la coupe du Çürük dağ sur une carte géologique simplifiée (d'après [23,25], modifié).
Location of the Çürük dağ section on a simplified geological map (after [23,25], modified).

Colonne stratigraphique de la coupe du Çürük dağ pour la limite Permien–Trias, avec situation de l'échantillonnage.
Stratigraphic log of the Çürük dağ section for the Permian–Triassic boundary with the sampling location.
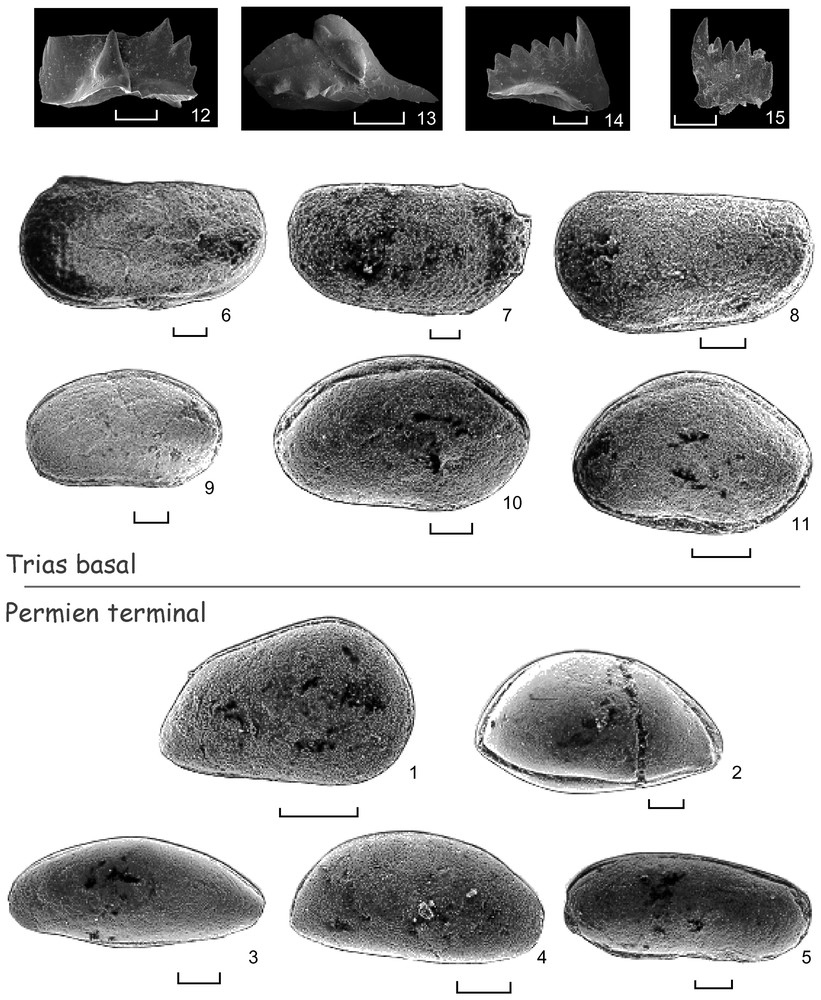
Spécimens représentatifs de la faune d'ostracodes et de conodontes à la limite Permien–Trias dans la coupe du Çürük dağ (Taurides occidentales, Turquie). Barre d'échelle : 100 μm. Toutes les photographies ont été réalisées au CIM de l'UPMC. 1 à 5 : Permien supérieur, Changsingien ; échantillon 01TK49.1 ; niveau 4, formation de Pamucak. 1. Spinocypris sp. 1 ; vue latérale droite. 2. Acratia sp. 1 ; vue latérale gauche. 3. Acratia sp. 2 ; vue latérale gauche. 4. Acratia sp. 3 ; vue latérale gauche. 5. Fabalicypris sp. ; vue latérale gauche. 6–15 : Trias basal, Induen ; formation de Kokarkuyu. 6–8 : Revyia? sp. 1 ; niveau 11. 6. Vue latérale gauche ; échantillon 01TK57. 7. Vue latérale droite ; échantillon 01TK56. 8. Vue latérale gauche ; échantillon 01TK56. 9. Bairdiocypris sp. 1 ; vue latérale gauche ; échantillon 01TK56 ; niveau 11. 10. Bairdia sp. 1 ; vue latérale gauche ; échantillon 01TK57 ; niveau 11. 11. Bairdia sp. 2 ; vue latérale gauche ; échantillon 01TK57 ; niveau 11. 12. Isarcicella isarcica staeschei (Dai & Zhang), vue latérale. Zone à I. isarcica : échantillon T11-3-150, niveau 10. 13. Isarcicella isarcica staeschei (Dai & Zhang), vue supérieure. Zone à I. isarcica. Zone. échantillon T11-3a-150, niveau 10. 14. Hindeodus parvus (Kozur & Pjatakova), vue latérale. Zones à H. parvus et I. isarcica ; échantillon T13-2b-150, niveau 10. 15. Hindeodus parvus (Kozur & Pjatakova), vue latérale. Zones H. parvus et I. isarcica. Échantillon T18-1-150, niveau 11.
Some representative specimens of ostracod and conodont fauna from the Permian–Triassic in the Çürük dağ section (Western Taurides, Turkey). Scale bare: 100 μm. All the pictures were realized at the CMI of UMPC, Paris, France. 1–5: Late Permian, Changsingian; sample 01TK49.1; level 4, Pamucak Formation. 1. Spinocypris sp. 1; right lateral view. 2. Acratia sp. 1; left lateral view. 3. Acratia sp. 2; left lateral view. 4. Acratia sp. 3; left lateral view. 5. Fabalicypris sp. 1; left lateral view. 6–15: Earliest Triassic, Induan; Kokarkuyu Formation. 6–8: Revyia? sp. 1; level 11. 6. Left lateral view; sample 01TK57. 7. Right lateral view; sample 01TK56. 8. Left lateral view; sample 01TK56. 9. Bairdiocypris sp. 1; left lateral view; sample 01TK56; level 11. 10. Bairdia sp. 1; left lateral view; sample 01TK57; level 11. 11. Bairdia sp. 2; left lateral view; sample 01TK57; level 11. 12. Isarcicella isarcica staeschei (Dai & Zhang), lateral view. I. isarcica zone. Sample T11-3-150, level 10. 13. Isarcicella isarcica staeschei (Dai & Zhang), upper view. I. isarcica zone. Sample T11-3a-150, level 10. 14. Hindeodus parvus (Kozur & Pjatakova), lateral view. H. parvus and I. isarcica zones. Sample T13-2b-150, level 10. 15. Hindeodus parvus (Kozur & Pjatakova), lateral view. H. parvus and I. isarcica zones. Sample T18-1-150, level 11.
In the Upper Permian samples (samples 01TK43-51 and 01TK60-64), the ostracod assemblage (Figs. 4.1–4.5) is similar to other Middle and Late Permian assemblages observed in the Tethys (see for example [9,10]). It is mainly made up of Bairdiacea (genera Bairdia, Acratia, Fabalicypris, Bairdiocypris) and Cypridacea (genus Spinocypris). We observe the absence of Palaeocopidae, perhaps due to a too deep environment. The ostracod assemblage is characteristic of a marine environment, open, circalittoral under tropical latitude.
In the basal beds of the Kokarkuyu Formation (samples 01TK58 to 01TK00), the ostracods are represented by Bairdiacea (Figs. 4.9–4.11) (mainly the genus Bairdia) et by Palaeocopidae (Kirkbyacea–genus ?Revyia) (Figs. 4.6–4.8). This assemblage, which shows indisputably a lower specific and generic diversity than assemblages observed in the Upper Permian levels, seems to point out a shallower environment and/or with salinity variations, always at a tropical latitude.
The Palaeocopidae, ostracods with straight dorsal border, have been considered to become extinct during the Permian and their presence as indicative of the Palaeozoic for a long time. Nevertheless, Palaeocopidae have been reported from the Early Triassic of southern China (Induan and Olenekian) [14,30,31] and from Pakistan (Olenekian) [29].
Ostracods of the lower part of the Kokarkuyu Formation in the Çürük dağ section are the oldest Triassic forms discovered up to now (parvus and staeschei biozones). The Palaeocopidae, Kirkbyacea, here discovered, are the youngest observed outside of southern China and Pakistan.
Occurrence of Palaeocopidae in the Early Triassic and the similarity between Permian and Induan assemblages suggest that the Induan forms are a survival assemblage after the Permo-Triassic mass extinction. This confirms our first investigations [8,21], which locate the main change of the ostracod fauna in the Middle–Late Triassic more certainly than at the Permian–Triassic boundary. This may be the recovery interval of Erwin [13], with a peak of diversification.
1 Introduction
Dans tous les groupes paléontologiques, surtout marins, la crise du Permien–Trias constitue un événement majeur. Même si elle se déroule par étapes, sur un intervalle de temps évalué entre 2 et 20 Ma, ses conséquences sur la biosphère sont impressionnantes : à la fin du Permien, 55 % des familles présentes dans les océans mondiaux disparaissent, soit 83 % des genres et 96 % des espèces [12,28].
Les ostracodes, à l'instar de tous les autres organismes, ont subi cette conjonction d'événements néfastes (Fig. 1). La disparition des formes à bord dorsal droit a été classiquement considérée comme marqueur de la limite Paléozoı̈que–Mésozoı̈que.
Les ostracodes du Trias inférieur marin sont très peu connus. Ceci est dû, selon toutes vraisemblances, à la rareté des faciès favorables à leur extraction. Quelques espèces sont reconnues dans le Trias inférieur (Induen–Olénékien) de la dépression Pré-Caspienne [18,26,27], au Népal [1], au Cachemire [1–3], dans le Bassin germanique [17], en Grèce [5], en Israël [15], en Chine du Sud [14,30,31] et au Pakistan [29].
2 Contexte géologique
La coupe du Çürük dağ (N36°41′32–E30°27′40) est située dans le Taurus occidental, dans les nappes d'Antalya (Turquie) (Fig. 2) [23,25]. Dans la région de Kemer, on peut observer la formation de Pamucak (Permien moyen et supérieur), surmontée de la formation de Kokarkuyu (Trias inférieur) [4,22,24].
La coupe du Çürük dağ (Fig. 3) débute par la formation de Pamucak, épaisse de 400 à 600 m. Ce sont des faciès de plate-forme interne à externe, d'âge Capitanien à Changsingien. La partie supérieure de cette formation, étudiée en détail ici, est composée de calcaires noirs noduleux, avec quelques niveaux à silex. Ils sont riches en algues calcaires (Dasycladacés) et en petits foraminifères (dominance des Milliolidés), avec également des niveaux à brachiopodes, échinodermes et crinoı̈des [22]. Les microfaciès montrent un wackstone à bioclastes de faible énergie (biomicrites à algues [4]). Puis, vers le niveau 5 (Fig. 3), les microfaciès montrent un grainstone de plus haute énergie, sans pour autant que le contenu faunistique change. Une cimentation tardive, des dissolutions et des recristallisations révèlent une forte diagenèse sur ces derniers niveaux [23]. Ce grainstone bioclastique est suivi par un grainstone oolitique (niveau 8) contenant des échinodermes et des lamellibranches, montrant, là aussi, une forte diagenèse, indicateur probable d'une zone adjacente émergée [22]. La formation de Kokarkuyu débute par 12 m de calcaires microbiens (niveaux 9 à 14), surmontés de 3 m de calcilutite (niveau 15), de 80 m de grainstone oolitique et se termine par des marnes bariolées. Le premier niveau du Trias (niveau 9) présente des stromatolites en dôme de 40 à 60 cm, puis une succession de stromatolites à structures laminaires horizontales et de thrombolites massives. Le microfaciès des stromatolites montre une alternance de ciments microsphérilutiques et de micrites grumeleuses, contenant des pseudopéloı̈des, des ostracodes, des microforaminifères et des filaments d'origine microbienne [19]. Le microfaciès des thrombolites est composé de micrites grumeleuses, avec des traces de filaments micritiques d'origine microbienne, qui peuvent parfois former des concrétionnements de types onchoı̈des. Ces niveaux sont très riches en foraminifères (Ammodiscus, Rectocornuspira, Cyclogira, Earlandia), qui dominent généralement la faune benthique de l'Induen et ont livré de rares conodontes (Isarcicella isarcica staeschei (Dai & Zhang) – échantillon T11 – et Hindeodus parvus Kozur & Pjatakova – échantillon T13 ; Figs. 4.12–4.15), dont l'association constitue un marqueur de la deuxième zone du Trias [19]. On constate donc le passage d'une sédimentation de plate-forme, constituée de carbonates squelettiques, à une sédimentation dominée par des communautés microbiennes [7]. La courbe isotopique du carbone montre, de plus, une forte variation négative à la base de la formation de Kokarkuyu, soulignant les perturbations importantes de l'environnement qui surviennent à la limite Permien–Trias [6]. Une régression marque également le passage entre les deux formations et provoque une lacune probable du Changsingien supérieur et éventuellement du Griesbachien inférieur.
3 Les ostracodes de la coupe du Çürük dağ
Au Permien terminal (échantillons 01TK43-51 et 01TK60-64), l'assemblage découvert dans la coupe du Çürük dağ est en tous points comparable aux assemblages de la Téthys. Il est composé majoritairement ici de Bairdiacea (principalement les genres Bairdia, Acratia, Fabalicypris, Bairdiocypris) et de Cypridacea (genre Spinocypris). Nous noterons l'absence de Palaeocopidae, peut-être due à un environnement trop profond. Les ostracodes présents dans le Permien terminal sont caractéristiques d'un milieu marin franc, ouvert, de type circalittoral, en zone tropicale. Dans ces niveaux, contrairement aux foraminifères (C. Jenny, comm. pers.), les ostracodes (Figs. 4.1–4.5) ne présentent pas de formes aberrantes ou de nanisme généralisé. La taille moyenne des spécimens observés ici est légèrement inférieure à celle des individus du Permien moyen et supérieur (voir par exemple [9,10]), mais est tout à fait du même ordre que les dimensions observées sur l'ensemble des formes du Paléozoı̈que supérieur de la Téthys.
Dans les premiers niveaux du Trias (01TK58–01TK00), les ostracodes sont représentés, là aussi, par des Bairdiacea (Figs. 4.9–4.11) (principalement le genre Bairdia) et par des Palaeocopes (Kirkbyacea–genre Revyia ?) (Figs. 4.6–4.8). Cet assemblage, qui montre une diversité générique et spécifique indéniablement plus faible que celle observée dans les derniers niveaux du Permien, semble indiquer un milieu un peu moins profond et/ou soumis à des variations de salinité, toujours situé en zone tropicale.
Les Palaeocopes, ostracodes à bord dorsal droit, ont été longtemps considérés comme s'éteignant au cours du Permien, leur présence constituant un marqueur du Paléozoı̈que. Toutefois, des Palaeocopes ont été signalés ponctuellement dans le Trias inférieur (formations de Feihsienkuan de l'Induen et Jialingjiang de l'Olénékien) de Chine du Sud dans le Sichuan et à Guizou [14,30,31] et au Pakistan (membre Narmia de la formation Mianwali de l'Olénékien [29]).
L'étude systématique détaillée des faunes est en cours [11].
Les ostracodes de la base de la formation de Kokarkuyu dans la coupe du Çürük dağ sont donc les formes triasiques les plus anciennes découvertes à ce jour (zone à staeschei).
Les Palaeocopes présents ici, des Kirkbyacea, sont les plus jeunes jamais observés hors de Chine du Sud et du Pakistan.
La présence des Palaeocopes dans le Trias basal et la similitude entre les assemblages du Permien et ceux de l'Induen suggèrent qu'il s'agit de faunes ayant survécu aux événements de la crise de la fin du Paléozoı̈que. Cette nouvelle découverte conforte nos premières investigations [8,21], selon lesquelles le renouvellement majeur chez les ostracodes se produit au Trias moyen et supérieur, plus vraisemblablement qu'à la limite Permien–Trias.
Les assemblages survivants après une extinction en masse sont généralement dominés par des taxons à vaste répartition géographique et à grande tolérance paléoenvironnementale [13]. Le genre Bairdia présente effectivement une vaste répartition paléogéographique ; en revanche, ses tolérances paléoécologiques sont limitées. La situation est exactement inverse pour les Palaeocopidae. Des investigations supplémentaires seront nécessaires pour élucider ce point.
Le développement des stromatolites opportunistes à la base de la formation Kokarkuyu est en accord avec l'existence d'un « intervalle de survie » au-dessus de la limite Permien–Trias. La faible diversité observée de l'assemblage d'ostracodes dans les niveaux de base de la formation Kokarkuyu témoigne de cet intervalle avant la diversification qui se produira plus tard (sensu [13]). Ces observations montrent que « l'intervalle de survie », de très courte durée, correspond au minimum aux premières biozones du Trias inférieur. Ceci est en accord avec Erwin [13], qui postule 1 à 3 Ma de durée pour cet intervalle.
Remerciements
Les auteurs remercient MM. les Pr. Jean Dercourt et Maurizio Gaetani pour leurs critiques constructives. Nous tenons à exprimer notre gratitude à l'association Naturalia & Biologia, qui a permis la mission de terrain de S.C.-S.