

Version abrégée
Le remplacement, durant le Tithonien, des radiolarites par les calcaires de Vigla (formés essentiellement par des Nannoconidés, Fig. 1b) marque un changement drastique des bioproducteurs et de l'état de paléofertilité dans la zone ionienne (Fig. 1a). La monotonie de faciès de ces calcaires est seulement interrompue dans la partie moyenne du Crétacé, avec la perte soudaine des carbonates et le dépôt de la « zone siliceuse supérieure » [13]. Dans la localité de Paliambela, en Grèce nord-occidentale (Fig. 1a), nous avons pu subdiviser la « zone siliceuse supérieure » en trois termes (Fig. 2a). Nos résultats sur l'intervalle radiolaritique inférieur nous conduisent à définir le « niveau Fourcade », en hommage à Éric Fourcade (micropaléontologue, stratigraphe). Des argillites riches en matière organique dominent la base de ce niveau et alternent avec des calcaires siliceux. Les 4,5 m supérieurs du niveau Fourcade sont dépourvus de carbonates, et consistent en une alternance d'interlits argileux et des jaspes. Les analyses géochimiques et palynologiques indiquent que la matière organique provient d'une biomasse planctonique marine, laquelle a été plus ou moins transformée par des bactéries durant le dépôt et/ou la diagenèse précoce.
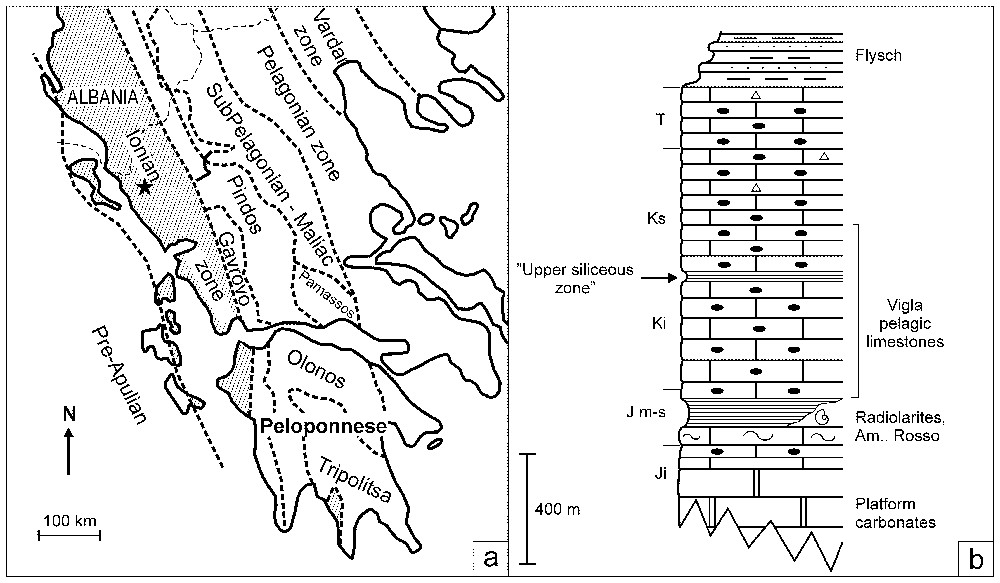
(a) Outcrops of the Ionian zone in Greece and Albania. Star indicates the location of Paliambela section in northwestern Greece. (b) Simplified lithostratigraphy of the Ionian zone. Note the stratigraphic position of the ‘upper siliceous zone’, towards the top of the Vigla Limestone. Ji: Lower Jurassic, Jm-s: Middle–Upper Jurassic, Ki: Lower Cretaceous, Ks: Upper Cretaceous, T: Tertiary.
(a) Extension géographique des affleurements de la zone ionienne en Grèce et en Albanie. L'étoile indique la localité de la coupe de Paliambela en Grèce nord-occidentale. (b) Lithostratigraphie simplifiée de la zone ionienne, avec indication de la « zone siliceuse supérieure », vers le sommet des calcaires de Vigla.
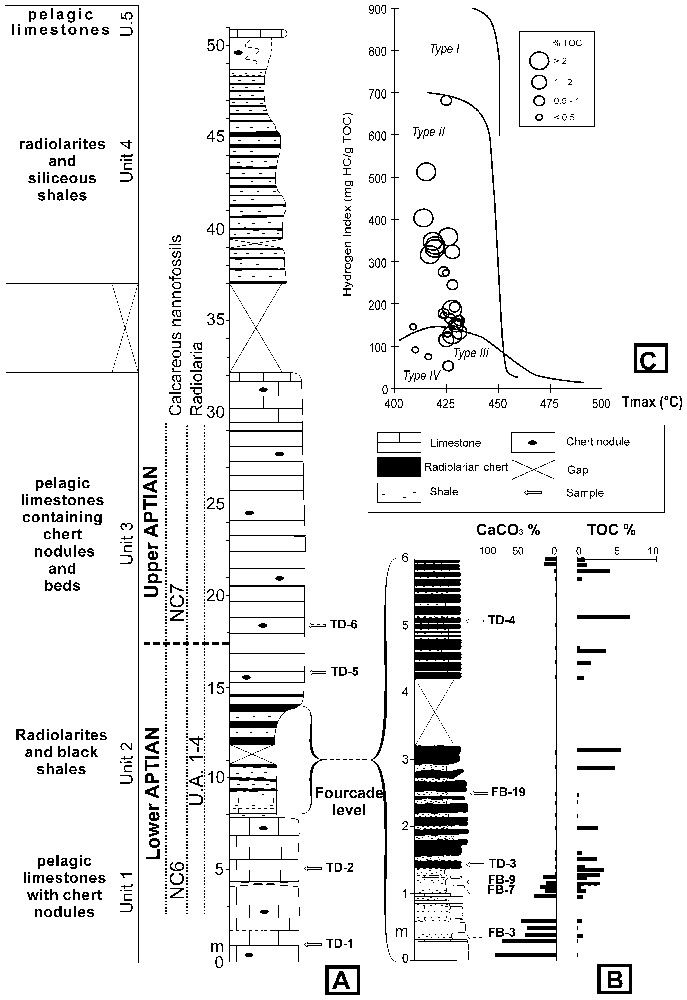
(A) Lithostratigraphic units within the ‘upper siliceous zone’, as observed at Paliambela. (B) Detailed lithostratigraphy of the Fourcade level (Unit ‘2’ and CaCO3–TOC (Total Organic Carbon) measurements of black shales. (C) Hydrogen vs Tmax plot-diagram for the analysed black shales of the Fourcade level.
(A) Unités lithostratigraphiques reconnues à l'intérieur de la « zone siliceuse supérieure » dans la coupe de Paliambela. (B) Détail de la lithostratigraphie au sein du niveau Fourcade et du contenu en carbonates (CaCO3) et carbone organique total (COT) au sein des interlits argilleux analysés. (C) Diagramme Tmax – indice d'hydrogène des échantillons analysés pour la matière organique.
Les nannofossiles calcaires observés dans les argillites marneuses à la base du niveau Fourcade forment un assemblage oligospécifique, caractérisé par quelques espèces de coccolithes et d'abondants nannolithes (dont certains sont d'une grande taille) et l'absence totale des Nannoconidés. La présence de l'espèce R. irregularis (Fig. 3) dans les calcaires de l'unité « 1 » (échantillons TD-1 et 2) établit que ceux-ci sont d'âge Barrémien terminal–Aptien inférieur. La présence de E. floralis et R. angustus dans l'échantillon TD-6 suggère que l'Aptien supérieur débute vers la base de l'unité « 3 ». Les assemblages des Radiolaires extraits des jaspes du niveau Fourcade sont dominés par des membres de la famille des Archaeodictyomitridae, aussi bien en termes de diversité que d'abondance. La présence des espèces P. carpathica et L. nudum dans l'échantillon TD-3 (Fig. 4) permet de corréler l'assemblage avec le Barrémien–Aptien inférieur [18].
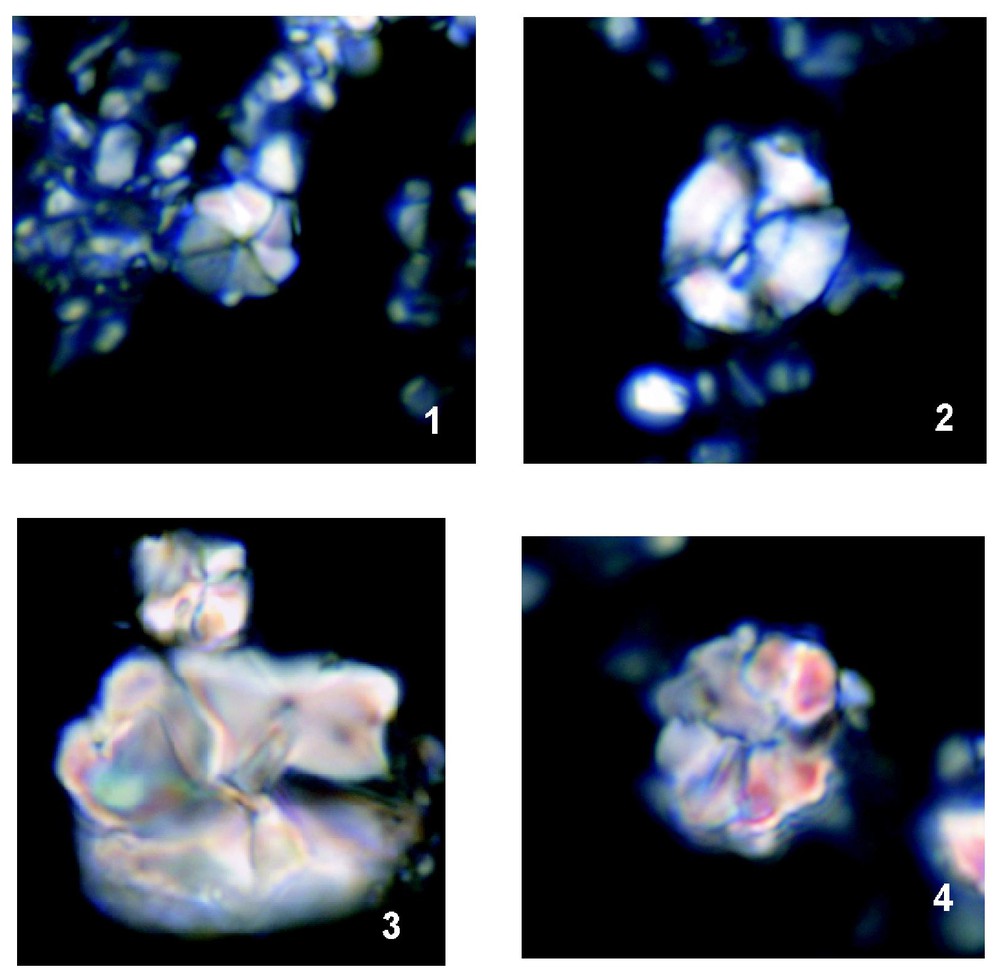
Key and dominant taxa in the calcareous nannofossil assemblage: (1) R. irregularis, sample TD-1; (2) W. barnesae, sample FB-3; (3) A. infracretacea, normal and giant size, sample FB-3; (4) E. floralis, sample TD-6. All micrographs are under polarised light and magnification is ×2100.
Espèces « marqueurs » et dominantes au sein des assemblages étudiés des nannofossiles calcaires. Toutes les photos ont été prises sous une lumière polarisée et au grossissement ×2100.

Age diagnostic radiolaria yielded by sample TD-3 (cf. Fig. 2B). (a) P. carpathica; (b) L. nudum; (c) D. communis; (d) T. lacrimula; (e) T. broweri; (f) T. pacifica. Scale corresponds to 100 μm for all specimens.
Radiolaires caractéristiques de l'assemblage extrait de l'échantillon TD-3 (cf. Fig. 2B). L'échelle indiquée en haut, à droite, correspond à 100 μm pour tous les spécimens.
Par conséquent, la biochronostratigraphie intégrée des nannofossiles calcaires et des radiolaires permet de contraindre l'âge du niveau Fourcade à l'Aptien inférieur. Outre son âge, l'abondance de la matière organique d'origine marine, l'absence des Nannoconidés ( « crise des Nannoconidés » de Erba [10]) et la présence d'abondants nannolithes de grande taille suggèrent que le niveau Fourcade peut être corrélé avec l'événement anoxique global OAE 1a. Cet événement est actuellement plutôt considéré comme le résultat d'une haute productivité océanique, suite à une intense activité volcanique, qui a exacerbé un climat global chaud de type greenhouse [14,21]. L'expression sédimentaire de cet événement dans la zone ionienne est caractérisée par la pauvreté ou absence des carbonates, par opposition à des intervalles du même âge en Italie (niveau Selli en Ombrie–Marches, sur la ride de Trente, dans le Gargano et en Sicile), lesquels sont plus riches en plancton calcaire.
L'enregistrement des bioproducteurs sédimentaires planctoniques en zone ionienne de Grèce suggère que l'installation d'un contexte paléoécanographique de haute productivité durant l'intervalle OAE 1a a d'abord affecté la structure des communautés nannofloristiques, en favorisant certains groupes (pullulement et « gigantisme » des nannolithes opportunistes), mais en en défavorisant d'autres ( « crise des Nannoconidés »). Enfin, avec le temps, la paléoproductivité a dû s'accroı̂tre d'avantage, comme ceci est suggéré par la présence exclusive des radiolaires dans la partie moyenne et supérieure du niveau Fourcade et par la plus forte abondance de matière organique dans cet intervalle dépourvu en carbonates. Il est néanmoins difficile d'affirmer si l'absence des nannofossiles calcaires au sein de ce sous-intervalle supérieur est due à l'arrêt total de la production carbonatée par la « machine » nannofloristique, ou si ceci est dû à une augmentation de la dissolution des boues à nannofossiles calcaires, avec la remontée locale du NCC, comme ceci est bien connu dans les contextes océanographiques modernes de haute productivité.
1 Introduction
The Ionian zone of the external Hellenides (Fig. 1a) corresponds to a relatively deep basin created following Late Liassic intense block-faulting and segmentation of a previously unique carbonate platform, situated at the northern margin of Gondwana [9]. Siliceous radiolarian ooze accumulated since the late Middle Jurassic [6] and its intrabasinal dispersal was controlled by the action and intensity of submarine currents [8]. The Late Tithonian onset of the Maiolica-type Vigla Limestone [13] marks a sharp switch in sedimentation from carbonate-free radiolarites to nannofossil-rich pelagic limestones (Fig. 1b), made essentially of Nannoconids. The facies monotony of the Vigla Limestone breaks only in the mid Cretaceous part of the sequence with the sudden loss of carbonates and accumulation of the ‘upper siliceous zone’ [13], for which we here present preliminary results of a multidisciplinary stratigraphic and geochemical study. The results further our understanding of the palaeoceanographic conditions that influenced the abundance of different sediment-producing planktonic groups and favoured organic matter preservation in the Ionian zone.
2 Access and lithological description of the studied section
The studied section crops out in a relatively remote area of the Khionistra mountains (Fig. 1a), along the Paliambela ravine. The ‘upper siliceous zone’ crops out in this locality along the pathway leading to the abandoned village of Elataria (Fig. 1a) and can be further subdivided into a number of alternating carbonate and siliceous units (Fig. 2A). The platy limestones of the lower unit ‘1’ display the typical Maiolica facies. They are overlain by 6 m of argillites and radiolarites (unit ‘2’), studied here in detail and named ‘Fourcade level’ in honour of the late Dr Éric Fourcade (Paris) for his contribution to Stratigraphy and Palaeogeography. Argillite facies dominate the base of the Fourcade level and display traces of jarosite (a sulphate resulting from pyrite alteration). Argillites alternate with thin beds of laminated, but partially bioturbated limestones (packstones), which contain pyrite, fish scales, and abundant organic matter. Further up the sequence, the Fourcade level is characterised by a total absence of carbonates as confirmed by analyses of shales intercalated within the chert bands (see Fig. 2). Carbonate ooze accumulated again for some time (unit ‘3’) but was once more interrupted while the c. 18 m thick radiolarite unit ‘4’ was deposited. In this account, we will concentrate on the study of the Fourcade level (unit ‘2’).
3 Geochemical and palaeontological characterisation of Fourcade level
3.1 Carbonate and organic content
Calcium carbonate content was determined using a carbonate bomb, whereas total carbon content, source, and thermal maturation of the organic matter were estimated using a Rock-Eval OSA device [12]. Most of the studied samples are virtually devoid of carbonate (<3%) except for the basal and uppermost layers, which contain up to 84% and 16% calcite-equivalent carbonates, respectively (Fig. 2B). TOC contents range from 0 to 6.65%, while HI-values range 55 to 520 mg HC/g TOC (Fig. 2C). Samples with low to medium HI-values (55 to 200 mg HC/g TOC) are associated with low organic carbon content.
3.2 Palynology
Palynological preparations were made for ten samples across the Fourcade level.
Palynofacies is dominated by orange to dark-brown granular amorphous organic matter, which is usually attributed to phytoplankton and bacteria. Other organic components represent small quantities of small-sized wood fragments, fusinite, and some dinoflagellate cysts. These particles never represent more than 5% of the whole palynofacies. Most of the studied samples contain badly preserved spores, pollens, and dinoflagellates. Only one sample (FB 19; Fig. 2B) allowed the identification of the dinoflagellate species C. distinctum and T. castanea, the latter known to be from the Late Barremian–Maastrichtian interval [22].
3.3 Calcareous nannofossils
The limestones and marly argillites of units ‘1’, ‘2’ and ‘3’ (samples TD-1, TD-2, FB-3, FB-7, FB-9, TD-5, TD-6 and TD-7; Fig. 2) were processed according to standard techniques for calcareous nannofossil analysis under a light microscope. All the samples studied contain poorly preserved but diagnostic assemblages. Samples TD-1 and TD-2 are assigned to the Uppermost Barremian–Lower Aptian Chiastozygus litterarius zone (NC6 [19], modified by [4]) due to the occurrence of R. irregularis and the absence of R. angustus and E. floralis. The marly argillites of unit ‘2’ contain oligospecific assemblages (Fig. 3), characterised by very common W. barnesae and common ‘nannoliths’ (incertae sedis) such as A. terebrodentarius and A. infracretacea, some of which are of an unusually large size. The assemblage is also characterised by the virtual absence of Nannoconids. Calcareous nannofossils are more abundant in unit ‘3’; the occurrence of E. floralis in sample TD-6 allows us to recognise the Upper Aptian R. angustus zone (NC7 [19], modified by [4].
3.4 Radiolaria
Following laboratory processing with diluted hydrofluoric acid (4%), relatively well-preserved radiolarian assemblages were yielded by two chert samples situated within the carbonate-free interval of the Fourcade level (samples TD-3 and TD-4; Fig. 2B). The assemblages are well diversified, with over 25 species identified so far, the majority of the species being multicyrtid Nassellaria, and more particularly part of the family Archaeodictyomitridae, which is dominant both in terms of abundance and diversity. The presence of species P. carpathica and L. nudum in sample TD-3 (Fig. 4) is particularly important as it allows correlation with the Late Barremian–Early Aptian Unitary Associations (UA) 1–4 (zone H. asseni to zone Turbocapsula, subzone H. verbeeki [18]).
4 Discussion of the data and conclusions
No correlation can be established between richness of organic matter and lithology. The wide range of HI-values suggests that the organic matter is distributed between Type II and Type IV (Fig. 2C). Type II derived mainly from plankton or bacteria and is usually related to marine reducing environments, whereas Type IV argues for significant alterations, as a consequence of fully oxygenated palaeoenvironmental conditions. Temperatures of maximum pyrolytic yield (Tmax) average 425 °C, indicating that the organic matter did not experience high temperature during burial and is still immature with respect to oil-generation. Such a low thermal maturation of this part of the Ionian zone was previously deduced from studies on older stratigraphical levels [1,7]. Both geochemical and palynological data indicate that most of the organic matter of unit ‘2’ derived from a marine planktonic biomass, that was certainly more or less reworked by bacteria during settling and/or early diagenesis. As organic matter becomes more abundant towards the upper part of the studied interval, which is devoid of carbonates, it is likely that the organic matter was produced by organisms other than calcareous nannofossils.
The integrated biostratigraphic constraints provided by the calcareous nannofossils and radiolarians date the Fourcade level as Early Aptian in age. Despite palaeobiogeographic specificities or diagenetic modifications which might have altered the nannofloral components to some extent, we believe that the examined samples recorded a primary biosignal within the nannofloral community: the virtual absence of nannoconids and the presence of large Assipetra specimens in samples from the Fourcade level is the expression of the well known Early Aptian ‘nannoconid crisis’ [10,16,20].
The Early Aptian age, the abundance of preserved marine organic matter and biogenic (radiolarian) silica, the absence of Nannoconids but presence of abundant nannoliths of remarkably large size, suggest that the Fourcade level at Paliambela can be correlated with the Oceanic Anoxic Event (OAE) 1a. This is currently regarded as the result of enhanced oceanic productivity driven by abnormally high plate volcanism and an intensified greenhouse climate [14–16,21]. The sedimentary expression of this event in the Ionian zone is characterised by the paucity/absence of carbonates, unlike coeval intervals in Italy (i.e. Selli level in Umbria–Marche [18], in Cismon Drill site [11], in Gargano [17], and in Sicily [3]), which are slightly richer in calcareous plankton. It is noteworthy that both the Fourcade and Selli levels are characterised by organic matter of the same origin and that both can be subdivided into two distinct carbonate-poor/free intervals marked by the same lithological succession [5]: a lower marly interval, overlain by a thicker, carbonate-free interval, which is much richer in organic matter [2]. It is likely that the latter interval was the result of increased palaeoproductivity.
The sediment-producing planktonic record in the Ionian zone of Greece suggests that the onset of increasingly more fertile waters during the Fourcade level first affected the structure of calcareous nannofossil communities (‘nannoconid crisis’ but bloom and ‘gigantism’ of opportunist nannoliths). It was subsequently followed by massive blooms of radiolaria, as reflected by their abundance in the upper carbonate-free interval of the Fourcade level. The question remains whether the absence of calcareous plankton within the latter is due to complete shut-off of the carbonate-producing nannoplankton ‘factory’ or to increased dissolution of their already impoverished communities. Indeed, the CCD could have risen locally, as is commonly the case with oceanographic settings associated with high levels of primary productivity.
Acknowledgements
Funding by ‘Naturalia & Biologia’ and the French Ministry of Research (project ‘coup de pouce’ to T. Danelian) is gratefully acknowledged. The manuscript was improved by the constructive comments of the reviewers, H. Weissert and L. O'Dogherty. J. Sanfourche assisted with SEM work, F. Savignac with Rock-Eval analysis, B. David with nannofossil smear-slides and radiolarian picking, P. Ranguis and C. Abrial with photography and A. Lethiers with drafting.
Appendix
| Taxonomic appendix/Annexe taxonomique | |
| Dinoflagellates/Dinoflagellés | Circulodinium distinctum (Deflandre and Cookson, 1955) Jansonius, 1986 |
| Trichodinium castanea Deflandre, 1935 ex Clarke and Verdier, 1967 | |
| Calcareous nannofossils/ | Rucinolithus irregularis Thierstein in Roth and Thierstein (1972) |
| Nannofossiles calcaires | Rhagodiscus angustus (Stradner, 1963) Reinhardt (1971) |
| Eprolithus floralis (Stradner, 1962) Stover (1966) | |
| Watznaueria barnesae Black in Black and Barnes (1959) | |
| Assipetra terebrodentarius (Applegate et al. in Covington and Wise 1987) | |
| Rutledge and Bergen in Bergen (1994) | |
| Assipetra infracretacea (Thierstein 1973) Roth 1973 | |
| Radiolaria/Radiolaires | Dictyomitra communis (Squinabol 1904) sensu O'Dogherty (1994) |
| Loopus nudum (Schaaf 1981) | |
| Pseudodictyomitra carpathica (Lozyniak 1969) | |
| Thanarla broweri (Tan 1927) | |
| Thanarla lacrimula (Foreman 1973) | |
| Thanarla pacifica Nakaseko and Nishimura 1981 |